методички по физиологии / методичка ЦНС / 1 Возбуждение. Подпороговый ток оказывает на клетку
1 Возбуждение - Стр 2
Одиночный цикл возбуждения.
К возбудимым тканям относятся только те, клетки которых генерируют потенциал действия (ПД). Это мышечные и нервные клетки. Нередко к возбудимым тканям необоснованно относят и «железистую ткань», хотя железистой ткани нет, а имеются различные железы и железистый эпителий как вид тканей. В процессе активной деятельности железы в ней действительно регистрируются биоэлектрические явления, поскольку железа как орган состоит из различных клеток: соединительной ткани, эпителиальной, мышечной. ПД проводится по мембранам нервных и мышечных клеток, с его помощью передается информация и обеспечивается управление деятельностью клеток организма.
Невозбудимыми тканями являются эпителиальная и соединительная (собственно соединительная, ретикулярная, жировая, хрящевая, костная и гемотопоэтические ткани в совокупности с кровью), клетки этих тканей хотя и способны изменять свой мембранный потенциал, но не генерируют ПД при действии наних раздражителя.
Основными физиологическими свойствами возбудимых тканей являются: возбудимость, проводимость, рефрактерность, лабильность.Специфическим свойством мышечной ткани являетсясократимость.
Возбудимость — это свойство некоторых тканей генерировать потенциал действия (ПД) в ответ на раздражение. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. Раздражители вызывающие гиперполяризацию мембран будут приводить к процессу обратному возбуждению – торможению.
Возбудимость может быть охарактеризована кривой потенциала действия, в которой выделяют несколько фаз (рис.1 А). Отметим, что единой терминологии в классификации этих фаз нет, поэтому будем использовать наиболее часто употребляемые названия.
Рис. 1. Изменение мембранного потенциала (А) и возбудимости клетки (Б) в разные фазы потенциала действия.
МВ – фаза местного возбуждения;
Д – фаза деполяризации;
РБ– фаза быстрой реполяризации;
РМ – фаза медленной реполяризации;
Г – фаза следовой гиперполяризации;
Н – период нормальной возбудимости;
РА – период абсолютной рефрактерности;
РО– период относительной рефрактерности;
Н+– период первичной экзальтации;
Н++– период экзальтации;
Н–– период субнормальной возбудимости.
Вначале, под действием раздражителя, развивается местное возбуждение(фаза начальной деполяризации) - процесс медленной деполяризации мембраны от мембранного потенциала до критического уровня деполяризации (КУД). Если этот уровень не будет достигнут – ПД не формируется, а развивается только локальный ответ.
Разность между мембранным потенциалом покоя и критическим уровнем деполяризации называют пороговым потенциалом, его величина определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
Время фазы начальной деполяризации очень короткое, на кривой ПД она регистрируется только при большой развертке, и чаще всего является составной частью общей фазы деполяризации. Эта фаза развивается при достижении КУД, за счет открытия всех потенциалчувствительных Na+ - каналов и лавинообразного входа ионов Na+ в клетку по градиенту концентрации (входящий натриевый ток). В результате, мембранный потенциал очень быстро уменьшается до 0, и даже приобретает положительное значение. Графически – это восходящая часть кривой потенциала действия. В результате инактивации Na+ - каналов и прекращения поступления Na+ в клетку, рост кривой ПД прекращается и начинается ее снижение. Явление изменения знака мембранного потенциала называютреверсиейзаряда мембраны.
По мнению некоторых исследователей, фаза деполяризации заканчивается уже тогда, когда мембранный потенциал становится равным нулю, и весь период, когда величина мембранного потенциала превышает величину 0 мВ, следует считать отдельной фазой реверсии, т.к. ионные токи, определяющие развитие этой части ПД, имеют характерные особенности.
Период времени, в течение которого мембранный потенциал имеет положительное значение, называется овершут.
Нисходящая част кривой ПД – фаза реполяризации. Она определяется выходящим калиевым током. Калий выходит через постоянно открытые каналы утечки, ток через которые резко возрастает из-за изменения электрического градиента вызванного нехваткой снаружи ионов Na+ и через потенциалочувствительные, управляемыеК+- каналы, которые активируются на пике ПД.
Различают быструю и медленную реполяризацию. В начале фазы, когда активны оба типа каналов, реполяризация происходит быстро, к концу фазы, ворота потенциалочувствительных К+- каналов закрываются, интенсивность калиевого тока снижается и реполяризация замедляется. Она прекращается тогда, когда положительный заряд снаружи мембраны вырастет настолько, что окончательно затруднит выход калия из клетки.
Фазу медленной реполяризации называют иногда отрицательным следовым потенциалом, что не совсем верно, так как эта фаза не является потенциалом по определению и не является следовым процессом по механизму.
Фаза следовой гиперполяризации(следовой положительный потенциал) – увеличение мембранного потенциала выше величины потенциала покоя, которое наблюдается у нейронов. Развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+/K+АТФ-азы.
Механизм наблюдаемой иногда следовой деполяризации(следовой отрицательный потенциал) до конца не ясен.
Изменение возбудимости клетки при развитии возбуждения. Рефрактерность.
Возбудимость в различные фазы развития одного цикла возбуждения, вообще является переменной величиной. В ходе развития одного цикла возбуждения возбудимость изменяется в сторону, как повышения, так и понижения. Повышение возбудимости называется экзальтацией, понижение –рефрактерностью.
В изменении возбудимости от момента нанесения раздражения до завершения одиночного цикла возбуждения отмечается несколько периодов (фаз). (Рис.1. Б)
В период развития местного возбуждения наблюдается некоторое повышение возбудимости, которое получило название первичной экзальтации. Каждое нанесенное в это время дополнительное раздражение, по силе даже ниже порогового, ускоряет развитие местного потенциала. Это связано с тем, что пороговый потенциал уменьшается, и открытие воротного механизма Na+ -каналов облегчается.
Как только местное возбуждение достигает критической величины и переходит в потенциал действия (фаза деполяризации), возбудимость начинает быстро снижаться и в точке пика потенциала практически становится равной нулю. Это связано с полной инактивацией Na+ -каналов на пике ПД.
Время, в течение которого происходит это снижение возбудимости называется абсолютной рефрактерной фазой(периодом), а само снижение возбудимости - абсолютной рефрактерностью. Раздражение любой сверхпороговой силы, нанесенное в этот период, практически не может повлиять на развитие текущего возбуждения (потенциала действия).
В фазе реполяризации возбудимость мембраны последовательно восстанавливается, до исходного уровня, за счет постепенного восстановления активности инактивированных Na+ -каналов. Пока активны не все каналы – то этот период называетсяотносительной рефрактерной фазой, а состояние, в котором находится живой объект - относительной рефрактерностью. Эта фаза продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Раздражение, нанесенное в этот период, может вызвать усиление возбуждения только в том случае, если по силе оно будет больше величины порогового потенциала Длительность относительной рефрактерной фазы может быть значительно больше, чем абсолютной.
Вслед за периодом относительной рефрактерности наступает фаза экзальтации (повышенной возбудимости). Это связано с тем что мембранный потенциал снижается до величины КУД, при которой восстанавливается активность большей части Na+ каналов, а разница между величиной мембранного потенциала и КУД – пороговый потенциал – минимальна. В этой фазе может возникнуть повторная волна возбуждения даже на раздражения, которые значительно ниже порогового потенциала. Фаза экзальтации длится до тех пор, пока не восстановится исходная величина мембранного потенциала – потенциал покоя, при этом восстанавливается исходная величина возбудимости.
В фазы следовой гипер- и деполяризации возбудимость меняется незначительно и связана с колебаниями порогового потенциала.
Биологический смысл фазового изменения возбудимости в ходе развития одиночной волны возбуждения заключается в следующем.
Начальная фаза повышения возбудимостиобеспечивает условие, при котором каждый дополнительный раздражитель ускоряет процесс подготовки (местное возбуждение) к специфической (для данной ткани) приспособительной реакции.
Состояние абсолютной рефрактерностипозволяет данной ткани «без помех» осуществлять текущую приспособительную реакцию. Если бы в этих условиях возбудимость была нормальной, то дополнительное раздражение, вызвав дополнительное возбуждение, могло бы исказить эту реакцию, превратив ее в избыточную или недостаточную для данных условий.
Абсолютная рефрактерность защищает ткань от чрезмерных энергетических трат в процессе осуществления текущей приспособительной реакции. Сходную роль играет и относительная рефрактерность, с той разницей, что в данном случае живое образование в состоянии реагировать на раздражения, требующие срочного ответа. Именно поэтому для большинства тканей и органов, работающих непрерывно и не имеющих длительных периодов физиологического покоя (например, сердце), характерна более длительная по сравнению со скелетной мускулатурой рефрактерность.
Кроме того, рефрактерность - один из факторов, определяющих максимальный (предельный) ритм импульсации клетки, что лежит в основе например кодирования и декодирования сигнала структурами нервной системы, регуляции восприятия, сокращения, обеспечении одностороннего проведения возбуждения по нервам и др.
Состояние зкзальтациисоздает условия готовности ткани к ответу на повторное раздражение не только прежней силы, но и более слабой.
Лабильность,или функциональная подвижность, одно из физиологических свойств живых тканей. Это свойство описано в 1892 г. Н. Е. Введенским, который установил, что скорость протекания процесса возбуждения в тканях различна. Каждая возбудимая ткань способна на раздражение отвечать только определенным количеством волн возбуждения. Так, нервное волокно способно воспроизводить до 1000 импульсов в секунду, поперечно-полосатая мышца только 200 250 имп/с.
Мерой лабильности, по Н. Е. Введенскому, является то наибольшее количество волн возбуждения, которое возбудимая ткань может воспроизводить в 1 с в точном соответствии с ритмом наносимых раздражений без явлений трансформации (переделки) ритма, т.е. не уменьшая и не увеличивая его.
Лабильность величина подвижная и может изменяться в достаточно широких пределах. В частности, лабильность широко варьирует в процессе ритмического раздражения. В одних случаях вследствие взаимодействия волн возбуждения лабильность может повыситься, в других понизиться. Повышение лабильности может привести к тому, что недоступные ранее ритмы деятельности станут доступными. На основании этого А. А. Ухтомский сформировал представление об «усвоении ритма», как способности ткани отвечать на раздражение более высоким или более низким ритмом возбуждения по сравнению с его исходным уровнем. Усвоение ритма зависит от текущих изменений обмена веществ в ткани во время ее деятельности
Явление усвоения ритма играет важную роль в процессах врабатывания и тренировки. Снижение лабильности, происходящее в процессе деятельности, приводит к иному результату, способность ткани к ритмической работе уменьшается. Лабильность может быть измерена косвенным путем по величине хронаксии(см. ниже) возбудимых тканей. Чем короче хронаксия, тем выше лабильность. Определение лабильности весьма важно в физиологии труда и спорта.
Проводимость - способность живой ткани проводить возбуждение, которое, возникая в рецепторе, распространяется по нервной системе и является для организма информацией, закодированной в нейроне в виде электрических или химических сигналов. Способностью к проведению возбуждения обладают практически все возбудимые ткани, но наиболее ярко она выражена в нервной ткани, для которой проводимость является одной из функций.
Подробно механизм и закономерности распространения возбуждения по мембранам возбудимых клеток рассмотрен в отдельном занятии.
Законы раздражения.
Процесс возбуждения начинается с действия на возбудимую клетку какого либо раздражителя.
Раздражитель-любое изменение внешней или внутренней среды организма, воспринимаемое клетками и вызывающееответную реакцию. По своей природе раздражители делятфизические (электрические, механические, температурные, световые)и химические.
В зависимости от степени чувствительности клеток к тому или иному раздражителю их подразделяют на адекватные и неадекватные. Адекватный раздражитель — это такой раздражитель, к которому клетка обладает наибольшей чувствительностью вследствие наличия специальных структур, воспринимающих этот раздражитель. Так, адекватным раздражителем для фоторецепторов сетчатки глаза, например, являются световые волны, адекватным раздражителем нейронов являются медиаторы и электрические импульсы.
Неадекватныераздражители в естественных условиях существования организма не воздействуют на возбудимые структуры. Однако, при достаточной силе и продолжительности действия, могут вызвать ответную реакцию со стороны возбудимых тканей, например, удар в глаз при достаточной силе может вызвать ощущение вспышки света.
В условиях физиологического эксперимента в качестве раздражителя чаше всего используют электрический ток. Электрический ток легко дозировать, и он является адекватным раздражителем для возбудимых тканей, так как их функциональная активность всегда сопровождается электрическими явлениями.
Определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани отражают законы раздражения. К законам раздражения относятся:
Закон силы.
Для возникновения возбуждения решающее значение имеет сила раздражителя. Возбуждение возникают только в том случае, если сила действующего раздражителя достигает минимальной, критической величины, которая характеризуется порогом возбуждения. По отношению к этой величине, по своей силе раздражители могут быть подпороговыми, пороговыми и надпороговыми.
Подпороговый раздражитель- это раздражитель такой силы, который не вызывает видимых изменений, но обусловливает возникновение физико-химических сдвигов в возбудимых тканях, например локального ответа. Однако степень этих сдвигов недостаточна для возникновения распространяющегося возбуждения.
Пороговый раздражитель- это раздражитель минимальной силы, который впервые вызывает минимальную измеримую ответную реакцию со стороны возбудимой ткани. Именно эту пороговую силу раздражителя называютпорогомраздражения или возбуждения. Порог раздражения и является мерой возбудимости ткани. Между порогом раздражения и возбудимостью существует обратная зависимость:чем выше порог раздражения, тем ниже возбудимость, чем ниже порог раздражения, тем возбудимость выше.При достижении раздражителем величины порога, возникновение потенциала действия становится неизбежным.
Следует отметить, что порог раздражения показатель достаточно изменчивый и значительно зависит от исходного функционального состояния возбудимой ткани и практически никак не зависит от характеристик самого раздражителя
Надпороговый раздражитель- это раздражитель, сила которого выше, чем сила порогового раздражителя.
Закон силы - характеризует взаимосвязь между силой раздражителя и электрическим ответом, он может быть применен для простых и сложных систем.
Простая возбудимая система– это одна возбудимая клетка, которая реагирует на раздражитель как единое целое. Исключением является сердечная мышца, которая вся реагирует как одна клетка.Закон силы для простых возбудимых систем - подпороговые раздражители не вызывают возбуждения, а пороговые и сверхпороговые раздражители вызывают сразу максимальное возбуждение (Рис. 2).
При подпороговых значениях раздражающего тока возбуждение (электротонический потенциал, локальный ответ) носит местный (не распространяется), градуальный (сила реакции пропорциональная силе действующего стимула) характер. При достижении порога возбуждения возникает ответ максимальной силы (ПД). Амплитуда ответа (амплитуда ПД) не изменяется при дальнейшем увеличении силы раздражителя.
Закон силы для простых возбудимых систем известен как закон «все ли ничего».
Сложная возбудимая система– система, состоящая из множества возбудимых элементов (мышца включает множество двигательных единиц, нерв – множество аксонов). Отдельные элементы (клетки) системы имеют неодинаковые пороги возбуждения.
Закон силы для сложных возбудимых систем - амплитуда ответа пропорциональна силе действующего раздражителя (при значениях силы раздражителя от порога возбуждения самого легковозбудимого элемента до порога возбуждения самого трудновозбудимого элемента) (рис. 3). Амплитуда ответа системы пропорциональна количеству вовлеченных в ответ возбудимых элементов. При возрастании силы раздражителя в реакцию вовлекается все большее число возбудимых элементов.
В случае сложных систем, от силы раздражителя будет зависеть не только электрический, но и физиологический (функциональный) ответ ткани, например сила сокращения. В этом случае закон силы будет звучать следующим образом: чем больше сила раздражителя, тем выше, до определенного предела, ответная реакция со стороны возбудимой ткани. Этот предел будет определяться функциональными возможностями ткани.
Ответ минимальной силы – едва заметное сокращение - возникнет при достижении раздражителем пороговой величины. При этом сократятся мышечные волокна имеющие наименьший порог возбуждения.
Ответная реакция на надпороговый раздражитель будет выше и по мере его увеличения некоторое время также возрастает за счет вовлечения в сокращение все новых мышечных волокон, которые имеют более высокие пороги возбуждения. По достижении определенной величины раздражителя, рост силы сокращения прекратится, значит, в сокращение вовлечены все мышечные волокна. Такую ответную реакцию называют максимальной, астепени силы раздражителя, находящиеся между пороговой и максимальной –субмаксимольными.
| Амплитуда ответа а лЛ И Т У Д а ответа | | Амплитуда ответа | |


Рис. 2. Зависимость силы реакции простой Рис. 3. Зависимость силы реакции сложной
возбудимой системы от силы раздражителя. возбудимой системы от силы раздражителя. ПВ – порог возбуждения. ПВMIN– порог возбуждения самого
легковозбудимого элемента,
ПВMАХ– порог возбуждения самого
трудновозбудимого элемента.
Увеличение силы раздражителя выше максимального какое-то время не сказывается на величине ответной реакции. Такую силу раздражителя называют супермаксимальной. Но при достаточно большом увеличении силы раздражителя, сила ответной реакции начинает снижаться. Такую величину силы раздражителя называютпессимальной.
Пессимальный ответ и есть тот определенный предел, до которого может расти ответная реакция. Превышение этого предела при спортивных, интеллектуальных, эмоциональных и любых других нагрузках не имеет никакого физиологического смысла для получения результата.
Закон силы-времени (силы-длительности)
Эффективность раздражителя зависит не только от силы, но и от времени его действия. Длительность действия раздражителя, способна компенсировать недостаток силы раздражителя и при его недостатке привести, тем не менее, к возникновению распространяющегося потенциала действия, поэтому важно определять не только пороговую силу, но пороговую длительность раздражителя. Учение о хронаксии как пороговом времени необходимом для возникновения возбуждения было создано французским ученым Лапиком.
Связь между силой и временем действия раздражителя характеризует закон силы длительности - сила раздражителя, вызывающего процесс распространяющегося возбуждения, находится в обратной зависимости от длительности его действия, т.е., чем больше сила раздражителя, тем меньше времени он должен действовать для возникновения возбуждения. Зависимость между силой раздражителя и продолжительностью его воздействия, необходимого для возникновения минимальной ответной реакции живой структуры, очень хорошо можно проследить на так называемой кривой силы - времени (кривая Гоорвега - Вейса - Лапика) (Рис 4).
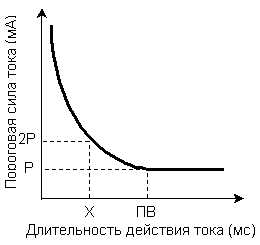
Рис.4.Зависимость пороговой силы раздражителя от времени его действия. Р – реобаза, ПВ – полезное время, Х – хронаксия.
Из кривой следует, что ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и как бы ни велика была сила раздражителя, при недостаточной длительности его воздействия ответной реакции не будет.
Минимальная сила раздражителя, способная, при неограниченном времени действия вызвать возбуждение, была названа Лапиком реобазой.Наименьшая длительность действия раздражителя силой в одну реобазу, достаточная для возникновения ответной реакции называется –полезным временем.
Но определение точки соответствующей на кривой величине полезного времени по техническим причинам затруднено, поэтому Лапиком было предложено измерять не полезное время, а условную величину – хронаксию.
Хронаксия – это наименьший промежуток времени, в течение которого ток, равный по силе удвоенной реобазе, вызывает в ткани возбуждение. Она и есть показатель пороговой длительности раздражения. Хронаксия измеряется в (тысячные доли секунды). По величине хронаксии можно судить о скорости возникновения возбуждения в ткани: чем меньше хронаксия, тем быстрее возникает возбуждение.
Определение хронаксии - хронаксиметрия -получило широкое распространение в физиологии спорта, в клинике, для определения функциональных возможностей и сохранности возбудимых тканей.
Закон крутизны нарастания силы раздражителя (закон аккомодации, закон Дюбуа-Реймона)
Для возникновения возбуждения имеет значение не только сила и время действия тока, но и скорость нарастания силы раздражителя в единицу времени от нулевой величины до величины порога. Степень крутизны, т.е. величина прироста силы раздражения в единицу времени, представляет собой градиент раздражения. Зависимость ответной реакции от градиента раздражения характеризует закон градиента (крутизны нарастания) раздражения - чем выше градиент раздражения, тем сильнее (до определенных пределов) ответная реакция возбудимого образования (рис 5).
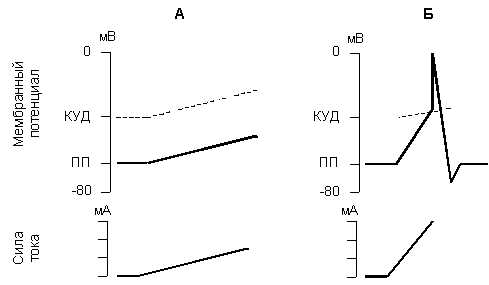
Рис. 5. Изменение мембранного потенциала и критического уровня деполяризации при медленном (А) и быстром (Б) нарастании силы раздражающего тока.
При действии медленно нарастающего раздражителя возбуждение возникает при его гораздо большей силе, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации.Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает.
Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого - окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще.
Способность к аккомодации различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
studfiles.net
§ 2. Потенциал покоя клетки
Потенциал покоя - неизменная во времени разность электрических потенциалов, которая возникает между внутренней и наружной поверхностями клеточной мембраны в состоянии термодинамического равновесия (иначе - в состоянии покоя) вследствие переноса K+ , Na+ и СI- через мембрану.
Формула для значения потенциала покоя была получена Гольдманом, Ходжкиным и Катцем и называется по именам этих авторов:
Потенциал покоя
0 = -
Однако главную роль в возникновении мембранного потенциала покоя играют ионы К+.
Для большинства клеток 0 варьируется в пределах (– 60 – –100) мВ, проницаемости мембраны для ионов Na+, K+, Cl – при формировании 0 соответственно следующему соотношению:
Именно потенциал покоя определяет (поддерживает) то различие концентраций ионов Na+, K+ и Cl- в цитоплазме и межклеточной жидкости, которое соответствует состоянию термодинамического равновесия и определяет нормальную жизнедеятельность клетки.1
6. Закономерности возбуждения тканей электрическим током. Критический потенциал возбуждения.
Нервная, мышечная и железистая ткани организма называются возбудимыми. Такое название этих тканей связано с тем, что их клетки из состояния покоя могут переходить в состояние возбуждения.
Главной особенностью этого состояния является новое распределение электрических зарядов на клеточной мембране (рис. 6). В состоянии возбуждения клетка деполяризована. Переход в состояние возбуждения возможен при действии на клетку возбудимых тканей различных физических или химических факторов. Они называются раздражителями (или стимулами). Одним из раздражителей является электрический ток. Из опытов известно, что возбуждение ткани или клетки электрическим током произойдет только тогда, когда сила тока I будет равна или больше некоторой величины In; Inназывают пороговым током (порогом возбуждения). Пороговый ток определяют из уравнения Вейса-Лапика:
(I In возбуждение).
In= +b (1)
В формуле (1) - время действия тока (миллисекунды), а и b зависят от вида и функционального состояния ткани и ее клетки.
Обычно ток силы In действует на небольшой участок клеточной мембраны. В результате мембранный потенциал покоя 0 на этом участке изменяется на определенную величину. Обозначим это изменение U. Теперь мембранный потенциал становится равным 0 + U и называется критическим потенциалом возбуждения ( кр), то есть кр = 0 + U, причем U>0 и кр > 0 .
7. Генерация (появление) потенциала действия, его характеристики.
Под действием порогового тока In, мембранный потенциал достигает критического значения кр . Это приводит к увеличению проницаемости клеточной мембраны для ионов Na+, концентрация которых снаружи клетки больше, чем внутри (Сe>Сi). Теперь положительные ионы натрия в большом количестве начинают проходить внутрь клетки. Поэтому отрицательный мембранный потенциал быстро повышается до нуля, а затем становится положительным (рис. 7).
Когда для ионов натрия действие градиента концентрации будет уравновешено действием градиента электрического потенциала на мембране, мембранный потенциал станет равным своему максимально возможному положительному значению max (рис.7).
Определенный участок клеточной мембраны окажется возбужденным и деполяризованным. Его внутренняя сторона будет иметь положительный, а внешняя отрицательный электрический заряд (см. рис.6 ). Этот процесс изменения мембранного потенциала от потенциала покоя до max называется деполяризацией.
Теперь диффузия ионов К+ из клетки наружу будет изменять мембранный потенциал до тех пор, пока он не вернется к значению потенциала покоя 0 Этот процесс называется реполяризацией (рис.7).
Длительность фазы деполяризации мала и для нервных и мышечных клеток составляет 0,5 - 1 мс (мс - миллисекунда). Длительность реполяризации зависит от вида клеток: для нервных клеток и скелетных мышц она равна 0,5 - 1 мс, для клеток сердечной мышцы ~ 300 мс.
Полное изменение мембранного потенциала во времени, которое происходит при возбуждении клетки, называется потенциалом действия.
8. Распространение потенциала действия по аксону (нервному волокну) не покрытому миелиновой оболочкой.
Аксон или нервное волокно - это длинный цилиндрический отросток, который отходит от каждой периферической нервной клетки.
Возбуждение аксона на каком-то участке приводит к деполяризации мембраны в этом месте (рис.8) и повышению мембранного потенциала до max. На соседнем, невозбужденном участке, потенциал равен 0. Под действие разности потенциалов max - 0 между возбужденным и невозбужденным участками аксона возникают локальные токи. Они показаны стрелками на рис.8. Эти токи заменяют пороговый ток и приводят к образованию потенциала действия на невозбужденном участке аксона. Затем по той же причине потенциал действия возникает на следующем участке волокна и так далее. Иначе говоря, происходит его распространение со скоростью до 25 м/с.
Не покрытые миелиновой оболочкой аксоны строят нервную систему беспозвоночных животных. Аксон позвоночных покрыт миелиновой оболочкой, которая разделяется промежутками - перехватами Ранвье. Такое строение аксона увеличивает скорость распространения потенциала действия во много раз.
C помощью потенциала действия в живом организме передается информация от рецепторов к нервным клеткам (нейронам) мозга и от них к мышцам и железистым органам.
В результате многократной генерации потенциала действия концентрация ионов Na+ в межклеточной жидкости и K+ в цитоплазме может резко измениться. Восстановление нормальной концентрации этих ионов обеспечивает работа Na+-K+-насоса.
1 Распределение электрических зарядов на мембране в состоянии покоя показано на рис.5 (состояние поляризации клетки)
studfiles.net
Электрические явления в нервах и мышцах
При определенных условиях в нервах и мышцах возникают электрические токи. Различают: 1) токи, или потенциалы, покоя и повреждения и 2) токи, или потенциал, действия – акционные точки.
Токи покоя
При отсутствии раздражения и, следовательно, возбуждения, существует разность потенциалов между наружной поверхностью цитоплазмы живой клетки (мембраной) и её внутренним содержимым.
Поверхностный слой цитоплазмы живой клетки, находиться в состоянии покоя, имеет положительный электрический заряд, а цитоплазма внутри клетки заряжена электроотрицательно. Поэтому, введя в клетку микроэлектрод и приложив другой электрод к поверхности клетки, можно отвести ток к регистрирующему прибору и измерить потенциал покоя, мембранный или клеточный потенциал.
Токи покоя, или клеточные потенциалы, обнаруживаются в неповрежденных живых клетках. Современными методами отведения электрического тока и его регистрации установлено, что токи покоя отдельных клеток животных, например гигантских нервных волокон кальмара, — 50-60 мв (милливольт), миелиновых нервных волокон амфибий – 70 мв, теплокровных – 90 мв, большинства тел нейронов – 80-100 мв, поперечнополосатых мышц амфибий – 80 мв, эпителия амфибий – 20 мв, волокон сердечной мышцы животных с постоянной температурой тела – 95 мв.
Токи повреждения
При повреждении живой ткани открывается доступ к внутреннему содержимому клеток, которое заряжено электроотрицательно. Поэтому если приложить электрод к неповрежденному участку ткани, находящемуся в состоянии покоя и, следовательно, заряженному электроположительно, а другой электрод приложить к поврежденному или перерожденному участку, заряженному электроотрицательно, то при соединении обоих электродов проводником тока с регистрирующим прибором можно измерить разность потенциалов.
Токи повреждения можно обнаружить на нервно-мышечном препарате. Если разрезать мышцу или любым способом повредить ее в одном участке (прижечь, раздавить и т. д.) и затем, приложив неполяризующиеся электроды, соединить срез мышцы или участок повреждения с целой поверхностью мышцы, не подвергающейся раздражению, то от целой поверхности к участку разреза или повреждения направится электрический ток. Сильно поврежденный участок всегда электроотрицателен по отношению к электроположительному неповрежденному участку. Следовательно, биоэлектрический ток повреждения, или альтерационный ток, возникает на границе целой и поврежденной частей ткани. Он обнаруживается и на некотором расстоянии от места повреждения, уменьшаясь с увеличением расстояния от места повреждения. Поэтому по месту происхождения его называют также демаркационным током. Однако ток повреждения появляется только тогда, когда отрицательно заряженный поврежденный участок соединяется проводником тока с положительно заряженным целым участком.
Величина тока повреждения меньше, чем клеточного потенциала, так как межклеточная и внеклеточная жидкости производят шунтирование (укорочение) при его измерении.
Разность потенциалов между целым и поврежденным участками скелетных мышц лягушки сразу же после повреждения равна 25-50 мв, наибольшая разность доходит до 80. В скелетных мышцах кролика токи повреждения равны 40 мв. В гладких мышцах токи повреждения меньше и быстрее уменьшаются до нуля. Разность потенциалов между целым и поврежденным участками седалищного нерва лягушки достигает 20-30 мв, седалищного нерва лошади — 6-16 мв, а седалищного нерва человека и обезьяны — 6-7 мв. Следовательно, токи повреждения нервов меньше, чем токи повреждения мышц. Особенно малы токи повреждения нервов человека и высших животных. Постепенно разность потенциалов уменьшается.
Когда нерв второго нервно-мышечного препарата набрасывается на первый нервно-мышечный препарат таким образом, чтобы он одновременно коснулся неповрежденного поврежденного участков мышцы, электрическая цепь замыкается. При этом мышца второго препарата сокращается в результате раздражения нерва током, возникающим в момент его замыкания между положительно заряженным нормальным участком мышцы первого препарата и отрицательно заряженным поврежденным ее участком.
Токи действия
Акционные токи, или токи (потенциалы) действия, возникают в раздражаемых тканях. Их появление характеризует возбуждение.
Для обнаружения тока действия нужно приложить один электрод к неповрежденному участку живой ткани, который находится в состоящих покоя и поэтому заряжен электроположительно, а другой — к раздражаемому участку и присоединить оба электрода к регистрирующему прибору. Раздражаемый участок при приложении к нему раздражителя немедленно или через несколько сотых миллисекунды становится электроотрицательным. Это падение потенциала’ доходит до максимума, а затем данный участок снова становится электроположительным. Различают однофазные и двухфазные токи действия.
Однофазный ток действия
При прикладывании одного из неполяризующихся электродов к неповрежденному участку нерва или мышцы, а другого — к поврежденному участку возникает электрический ток. Этот ток регистрируется при включении в цепь гальванометра или другого более чувствительною прибора.
Стрелка гальванометра отклонится в одну сторону вследствие электроотрицательного заряда поврежденного участка и электроположительного заряда нормального участка. Но если нанести раздражение по соседству с нормальным участком, то волна возбуждения, которая возникнет в месте раздражения, на некоторое время превратит заряд под нормальным участком в электроотрицательный. В этот момент тока не будет, так как под обоими электродами заряд станет электроотрицательным и стрелка гальванометра станет на нулевую линию. Когда волна возбуждения продвинется дальше и будет находиться между электродами, нормальный участок снова станет электроположительным и стрелка гальванометра снова отклонится в ту же сторону.
Когда волна возбуждения дойдет до поврежденного участка, положение стрелки гальванометра не изменится, так как под вторым электродом в месте повреждения уже имеется электроотрицательный заряд. Таким образом, стрелка гальванометра движется только в одну сторону, а запись этого движения регистрирует однофазный, или монофазный ток действия.
Это явление Дюбуа-Реймон назвал отрицательным колебанием тока покоя, которое возникает каждый раз, когда волна возбуждения проходит под электродом, расположенным на неповрежденном участке ткани.
Однофазный ток действия можно зарегистрировать также при раздражении ткани во время отведения клеточного потенциала. При внутриклеточном введении одного электрода и приложении другого электрода к поверхности клетки установлено, что клеточный потенциал не только исчезает в течение тысячных долей секунды, но что в течение этого промежутка времени амплитуда тока действия на 30-50 мв больше клеточного потенциала. Это объясняется тем, что в пункте прохождения волны возбуждения наружная поверхность мембраны становится электроотрицательной, а внутренняя — электроположительной. Продолжительность тока действия в нервных волокнах и клетках скелетных мышц составляет 0,1-5 мс.
Однофазный ток действия представляет собой местный электроотрицательный потенциал, мгновенно возникающий при очень слабых, подпороговых одиночных раздражениях и ограниченный несколькими миллиметрами от пункта раздражения. Величина местного низковольтного электроотрицательного заряда убывает по мере удаления от пункта раздражения. Чем больше подпороговое раздражение, тем больше возникающее возбуждение и амплитуда или отклонение однофазного тока действия.
Следовательно, местное возбуждение или местный сдвиг обмена веществ отличается градуальностью — возрастанием возбуждения по мере увеличения силы раздражения. Местное возбуждение наступает сразу после раздражения, т. е. без латентного периода. В участке слабого местного нераспространяющегося возбуждения возбудимость сохраняется, рефрактерность отсутствует. Если бы при местном возбуждении наблюдалась рефрактерность, то оно не могло бы перейти в пороговое, распространяющееся возбуждение.
Местный однофазный ток действия предшествует распространяющейся волне возбуждения и подготавливает ее. При одиночном пороговом раздражении волна возбуждения возрастает, достигает критического уровня и превращается из местной в распространяющуюся. Поэтому распространяющееся возбуждение наступает после латентного периода, продолжительность которого равна длительности местного нераспространяющегося возбуждения. В отличие от местного возбуждения распространяющееся возбуждение не градуально и сопровождается фазами изменения возбудимости. Потенциал действия, возникший в раздражаемом участке мембраны, вызывает возбуждение соседних участков мембраны и таким образом возбуждение распространяется по нервным и мышечным волокнам.
О прохождении волны возбуждения по возбудимой ткани судят прежде всего по токам действия. Однофазное колебание тока действия распространяющейся волны возбуждения в отдельных нервных и мышечных клетках состоит из нескольких составляющих: 1) высоковольтного потенциала, пика, 2) отрицательного следового потенциала и 3) положительного следового потенциала.
По сравнению со следовыми потенциалами пик имеет наибольшую высоту и наименьшую продолжительность. Пик имеет приблизительно одинаковую высоту на всем протяжении возбудимых тканей животных с постоянной температурой тела. Это указывает на то, что по мере распространения возбуждения вольтаж потенциала не падает. Этот факт свидетельствует, что химические вещества, необходимые для возникновения возбуждения, равномерно распределены по возбудимой ткани и что распространение возбуждения происходит самостоятельно, автоматически. Раздражение при достаточной интенсивности вызывает возбуждение только в том участке, который раздражается, а дальше возбуждение движется по ткани, вовлекая в сдвиг обмена веществ вещества, имеющиеся в возбудимой ткани.
Пик играет ведущую роль в распространении возбуждения. Высота пика различна у нервных волокон разных групп, имеющих неодинаковый диаметр, и изменяется в зависимости от физиологического состояния волокна и скорости проведения возбуждения. Чем толще нервное миелиновое волокно, тем меньше длительность пика. По мышечному волокну человека возбуждение проходит за 4-5 м/с.
В волокнах скелетных мышц однофазные токи действия различаются по амплитуде и по продолжительности. В волокнах сердечной мышцы они отличаются большей продолжительностью пика, который после начального крутого снижения начинает снижаться постепенно, а затем снова круто падает.
Токи действия в гигантских нервных волокнах кальмара достигают 90-110 мв, в миелиновых нервных волокнах амфибий 110 мв, в волокнах поперечнополосатых мышц амфибий — 120 мв, а в волокнах сердечной мышцы животных с постоянной темпера турой тела — 135 мв. Чем больше частота тока действия, тем меньше его вольтаж. Гладкие мышечные волокна отличаются тем, что их токи действия не превосходят мембранных потенциалов (не больше 70-80 мв).
Условия, улучшающие обмен веществ (повышение темпера туры, отдых), уменьшают продолжительность волны возбуждения, а условия, ухудшающие обмен веществ (утомление), увеличивают ее. С понижением температуры амплитуда пика уменьшается, а продолжительность его возрастает.
При температуре тела продолжительность подъема высоковольтного потенциала нерва составляет примерно 1/3 а продолжительность снижения — 2/3 его общей продолжительности. Предполагается, что крутой подъем пика обусловлен быстрым движением ионов натрия внутрь клетки.
Отрицательный следовой потенциал соответствует остаточным сдвигам распределения ионов калия и натрия. В отличие от пика он неустойчив и изменяется от условий среды. Он достигает 0,05 потенциала пика. Положительный низковольтный следовой потенциал также связан с перемещением ионов и равен примерно 0,002 потенциала пика.
Тепло и отдых уменьшают, а охлаждение и истощение увеличивают продолжительность следовой электроотрицательности. В нерве, лишенном кислорода, при действии наркоза и солей калия следовая низковольтная электроотрицательность отсутствует, а начальная высоковольтная электроотрицательность сохраняется, и при известных концентрациях наркоза даже не наступает заметных изменений её величины. Это указывает на то, что пик может возникать и при отсутствии кислорода, а следовая электроотрицательность связана с восстановительными процессами, которые совершаются в ткани вслед за возбуждением, и для её появления необходим кислород. Нарушение обмена веществ, утомление увеличивают продолжительность следовых потенциалов. Чем больше лабильность, тем меньше выражены следовые потенциалы. В высоколабильных мякотных волокнах человека и высших животных они почти не проявляются.
Электрические явления в нервах и мышцах развиваются в следующей последовательности. Сначала во все время раздражения отмечается электротоническое колебание, вызванное выходом катионов на поверхность клетки. Затем возникает местная низковольтная электроотрццательность. которая длится до начала появления высоковольтного потенциала. Это время от начала раздражения до начала пика, или латентный период, равно долям миллисекунды. После этого появляется пик; время»от начала его до вершины равно одной или нескольким миллисекундам. Продолжительность низковольтного отрицательного следового потенциала составляет несколько десятков миллисекунд и может доходить до 0,1 с и больше. Особенно велика продолжительность низковольтного положительного следового потенциала, которая равна десятым долям секунды.
Двухфазный ток действия
В естественных условиях одиночные волны возбуждения встречаются чрезвычайно редко; В организме в нервах и мышцах передаются серии волн возбуждения, взаимно влияющих друг на друга. Возбуждение, которое достигает порога или несколько превышает его, носит ритмический характер. В других случаях возбуждение в естественных условиях является градуальным, длительным, стойким, переходящим в торможение. В последнем случае при чрезмерно большой силе или частоте раздражения, превышающей меру лабильности, полностью угнетается высоковольтная электрическая активность.
Волновое распространяющееся возбуждение возникает из неволнового, градуального. Такая же эволюция возбуждения происходила в филогенезе. Представление о возникновении волнового возбуждения дает следующий простой опыт.
После приложения обоих неполяризующихся электродов к нормальным целым участкам возбудимой ткани и включения этой ткани в цепь гальванометра его стрелка останется на нуле, так как все участки нормальной ткани, находящейся в состоянии покоя, имеют одинаковый заряд. Но когда ткань раздражают у одного из электродов, то возникает возбуждение, и заряд ткани под этим электродом становится отрицательным. При этом стрелка гальванометра отклоняется в одну сторону. Передвигаясь далее, волна возбуждения оказывается между электродами, и в это время заряды под электродами вновь становятся одинаковыми и стрелка гальванометра возвращается к нулю. Но когда волна возбуждения доходит до второго электрода, то гальванометр опять отмечает разность потенциалов, так как под вторым электродом заряд становится отрицательным, а в то же самое время под первым электродом заряд положительный, так как под ним нег возбуждения. Теперь стрелка гальванометра отклоняется в противоположную сторону, а затем, после прекращения возбуждения, возвращается к нулю. Следовательно, когда по ткани проходит волна возбуждения, то стрелка гальванометра отклоняется сначала в одну сторону, потом в другую. Запись этого движения стрелки гальванометра регистрирует двухфазный ток действия.
Форма регистрации токов, или потенциалов, действия зависит от места возникновения возбуждения, от используемых приборов и от расположения электродов. В настоящее время их записывают посредством катодных осциллографов, не имеющих инерции, в отличие от гальванометров. Современные катодные осциллографы регистрируют биопотенциалы в миллионные доли вольта, продолжающиеся одну стомиллионную или даже миллиардную долю секунды. Эти токи отличаются сложностью, и их нередко расшифровывают при помощи электронных устройств.
Биопотенциалы каждой возбудимой ткани, например сердца, имеют характерный вид и изменяются в зависимости от ее функционального состояния. Записываются биопотенциалы кожи, эпителия, соединительной ткани, рецепторов, сетчатки глаза, скелетных мышц, пищеварительных желез, гладкой мускулатуры пищеварительного канала и других органов, разных отделов центральной нервной системы и даже одного нейрона. Первые исследования биопотенциалов больших полушарий головного мозга провели В. Я. Данилевский, Р. Кетон (1875), продолговатого мозга — И. М. Сеченов (1882).
Н. В. Введенский (1883, 1884) соединил мышцу проводами с телефонной трубкой и установил важнейшие законы, по которым протекает в ней возбуждение. Он же впервые применил телефон для изучения явлений возбуждения в нерве.
www.polnaja-jenciklopedija.ru
11 вопрос
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в виде кратковременного изменения мембранного потенциала на небольшом участке возбудимой клетки (нейрона иликардиомиоцита), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, в то время, как в покое она заряжена положительно. Потенциал действия является физиологической основой нервного импульса.
Благодаря работе «калий-натриевого насоса» концентрация ионов натрия в цитоплазме клетки очень мала по сравнению с окружающей средой. При проведении потенциала действия открываются потенциал-зависимые натриевые каналы и положительно заряженные ионы натрия поступают в цитоплазму по градиенту концентрации, пока он не будет уравновешен положительным электрическим зарядом. Вслед за этим потенциал-зависимые каналы инактивируются и отрицательный потенциал покоя восстанавливается за счёт диффузии в клетку отрицательно заряженных ионов хлора, концентрация которых в окружающей среде также значительно выше внутриклеточной.
Фазы потенциала действия
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
Отрицательный следовой потенциал — увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
Положительный следовой потенциал — от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
Общие положения
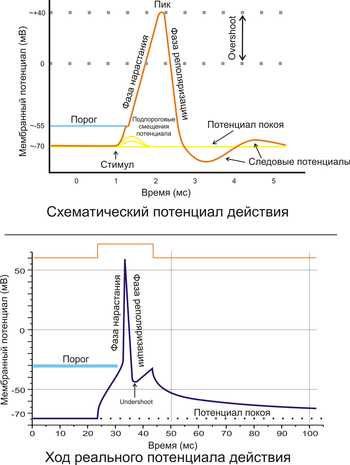
Рис. 2. A. Схематичное изображение идеализированного потенциала действия. B. Реальный потенциал действия пирамидного нейрона гиппокампакрысы. Форма реального потенциала действия обычно отличается от идеализированной.
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и хлора, внутри — ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны — в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий черезсинапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе — например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны — аксонном холмике, так что потенциал действия не распространяется на дендриты).
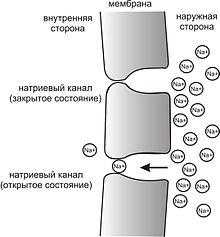
Рис. 3. Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственно
Это обусловлено тем, что на мембране клетки находятся ионные каналы — белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны — натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентомконцентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова — один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
Распространение потенциала действия
По немиелинизированным волокнам
По немиелинизированному волокну потенциал действия распространяется непрерывно. Проведение нервного импульса начинается с распространениемэлектрического поля. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые потенциалы. Сам потенциал действия не перемещается, он исчезает там же, где возник. Главную роль в возникновении нового потенциал действия играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то потенциал действия будет распространяться в обоих направлениях. Обычно же потенциал действия распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник потенциал. Одностороннее проведение потенциала действия обеспечивается свойствами натриевых каналов — после открытия они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» потенциал действия, он не возникает.
При прочих равных условиях распространение потенциала действия по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара потенциал действия может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
По миелинизированным волокнам
По миелинизированному волокну потенциал действия распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых потенциал действия, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2-й, 3-ий, 4-й и даже 5-й, посколькуэлектроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля.
«Скачкообразное распространение» увеличивает скорость распространения потенциала действия по миелинизированным волокнам по сравнению с немиелинизированными. Кроме того, миелинизированные волокна толще, а электрическое сопротивление более толстых волокон меньше, что тоже увеличивает скорость проведения импульса по миелинизированным волокнам. Другим преимуществом сальтаторного проведения является его экономичность в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых меньше 1 % мембраны, и, следовательно, необходимо значительно меньше энергии для восстановления трансмембранных градиентов Na+ и K+, расходующихся в результате возникновения потенциал действия, что может иметь значение при высокой частоте разрядов, идущих по нервному волокну.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения за счёт миелиновой оболочки, достаточно сравнить скорость распространения импульса по немиелинизированным и миелинизированным участкам нервной системы человека. При диаметре волокна около 2 µм и отсутствии миелиновой оболочки скорость проведения будет составлять ~1 м/с, а при наличии даже слабой миелинизации при том же диаметре волокна — 15—20 м/с. В волокнах большего диаметра, обладающих толстой миелинововой оболочкой, скорость проведения может достигать 120 м/с.
Скорость распространения потенциала действия по мембране отдельно взятого нервного волокна не является постоянной величиной — в зависимости от различных условий, эта скорость может очень значительно уменьшаться и, соответственно, увеличиваться, возвращаясь к некоему исходному уровню.
Активные свойства мембраны
Схема строения мембраны клетки.
Активные свойства мембраны, обеспечивающие возникновение потенциала действия, основываются главным образом на поведении потенциалзависимых натриевых (Na+) и калиевых (K+) каналов. Начальная фаза ПД формируется входящим натриевым током, позже открываются калиевые каналы и выходящий K+-ток возвращает потенциал мембраны к исходному уровню. Исходную концентрацию ионов затем восстанавливает натрий-калиевый насос.
По ходу ПД каналы переходят из состояния в состояние: у Na+ каналов основных состояний три — закрытое, открытое и инактивированное (в реальности дело сложнее, но этих трёх достаточно для описания), у K+ каналов два — закрытое и открытое.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и высчиляется черезкоэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Ходжкином и Хаксли.[1][2]
Потенциал действия (ПД) - это кратковременные высоко амплитуды и изменения МПС, возникающих при возбуждении. Основной причиной ПД является изменение проницаемости мембраны для ионов. Рассмотрим развитие ПД на примере нервного волокна. Регистрировать ПД можно при введении одного из электродов в волокно или располагая оба электрода на его поверхности. Проследим процесс формирования ПД при внутриклеточном методе. 1. В состоянии покоя мембрана поляризована и МПС равен 90 мВ. 2. Как только начинается возбуждение, величина этого потенциала уменьшается (это уменьшение называется деполяризацией). В ряде случаев потенциал сторон мембраны меняется на противоположный (так называемый овершут). Это первая стадия ПД - деполяризация. 3. Стадия реполяризации, при которой величина разности потенциалов падает почти до первоначального уровня. Эти две фазы в пиком ПД. 4. После пика наблюдаются следовые потенциалы - следовая деполяризация и следовая гиперполяризация (гиперполяризация - увеличение разности потенциалов между сторонами мембраны). Например, было 90 мВ, а становится 100 мВ. Развивается ПД очень быстро - за несколько миллисекунд. Параметры ПД: 1) переменный характер, поскольку меняется направление движения тока, 2) величина, которая благодаря овершута может превышать МПС; 3) время, в течение которого развиваются ПД и отдельные его стадии - деполяризация, реполяризация, следовая гиперполяризация. Как формируется ПД. В состоянии покоя «ворота» потенциалзависимых Na +-каналов закрыты. Закрытые также «ворота» потенциалзависимых К +-каналов. 1. Во время фазы деполяризации происходит активизация Na +-Kaналив. При этом изменяется конформационный состояние белков, входящих в состав «ворот». Эти «ворота» открываются, и проницаемость мембраны для Na + увеличивается в несколько тысяч раз. Na + лавообразные входит в волокно нерва. В настоящее время К +-каналы открываются очень медленно. Так, в волокно поступает значительно больше Na +, чем выводится из него К +. 2. Реполяризация характеризуется закрытием Na +-каналов. «Ворота» на внутренней поверхности мембраны закрываются - наблюдается инактивация каналов под воздействием электрических потенциалов. Инактивация происходит медленнее, чем активация. В настоящее время ускоряется активация К +-каналов и растет диффузия К + наружу. Таким образом, деполяризация связана преимущественно со входом Na + в волокно, а реполяризация - с выходом из него К +. Соотношение между входом Na + и выходом К + изменяется в процессе раз «витке ПД: в начале ПД входит Na + в несколько тысяч раз больше, чем получается К +, а затем выходит более К +, чем входит Na +. Причиной следовых потенциалов дальнейшие изменения соотношения между этими двумя процессами. Во время следовой гиперполяризации много К +-каналов еще остаются открытыми и К + продолжает выходить наружу. Восстановление ионных градиентов после ПД. Одиночные ПД изменяют разность концентраций ионов в нервном волокне и за его пределами очень мало. Но в тех случаях, когда проходит значительное количество импульсов, эта разница может быть весьма существенной. Восстановление ионных градиентов происходит тогда за счет усиления работы Na + / K +-HacociB - в большей степени нарушается этот градиент, тем интенсивнее работают насосы. При этом используется энергия АТФ. Часть ее выделяется в виде тепла, поэтому в этих случаях наблюдается кратковременное повышение температуры волокна. Условия необходимые для возникновения ПД. ПД возникает лишь при определенных условиях. Раздражители, действующие на волокну, могут быть разными. Чаще используется постоянный электрический ток. Он легко дозируется, мало травмирует ткань и ближайший тех раздражителей, которые существуют в живых организмах. При каких условиях постоянный ток может зумовитК появление ПД? Ток должен быть достаточно сильным, действовать определенное время, его нарастание должно быть быстрым. Наконец, имеет значение и направление тока (действие анода или катода). В зависимости от силы различают подпороговый (недостаточный для возникновения возбуждения), пороговый (достаточный) и надпороговый (чрезмерный) ток. Несмотря на то что подпороговый ток не вызывает возбуждение, он все же деполяризует мембрану, и эта деполяризация тем больше, чем выше его напряжение. Деполяризация, развивающийся при этом, называется локальным ответом и является видом местного возбуждения. Оно характеризуется тем, что не распространяется, величина его зависит от силы раздражения (закрном силовых отношений: чем больше сила раздражения, тем активнее ответ). При локальной ответа возбудимость ткани повышается. Возбудимость - это способность отвечать на раздражение и переходить в состояние возбуждения. Если сила раздражителя достаточное (пороговая), то деполяризация достигает определенной величины, называется критическим уровнем деполяризации (Ек). Для нервного волокна, покрытого миелином, Ек составляет около 65 мВ. Таким образом, разница между МПС (Е0), равный в данном случае 90 мВ, и Ек составляет 25 мВ. Эта величина (ДЕ = Е0-Ек) очень важна для характеристики возбудимости ткани. Когда Е0 увеличивается при деполяризации, возбудимость выше и, наоборот, снижение Е0 при гиперполяризации приводит к ее уменьшению. ГДЕ может зависеть не только от величины Е0, но и от критического уровня деполяризации (Ек). При пороговой силе раздражителя возникает ПД. Это уже не местное возбуждение, оно способно распространяться на дальние расстояния, подлежит закону «все или ничего» (при увеличении силы раздражителя амплитуда ПД не растет). Возбудимость при развитии ПД отсутствует или значительно снижена. ПД является одним из показателей возбуждения - активного физиологического процесса, которым живые клетки (нервные, мышечные, железистые) отвечают на раздражение. Во время возбуждения меняются метаболизм, температура клеток, нарушается ионная равновесие между цитоплазмой и внешней средой, происходит ряд других процессов. Кроме силы постоянного тока, возникновения ПД зависит также от длительности его действия. Существует обратная пропорциональная зависимость между силой тока и длительностью его действия. Подпороговый ток даже при очень длительном воздействии не приведет возбуждения. Надпороговый ток при слишком короткой действия также не приведет к возбуждению. Для возникновения возбуждения требуется также определенная скорость (крутизна) нарастание силы тока. Если повышать силу тока очень медленно, то при этом будет меняться Ек и Е0 может не достичь его уровня. Имеет значение и направление тока: ПД возникает при замыкании тока только тогда, когда катод помещается на внешней поверхности мембраны, а анод - в клетке или волокне. При прохождении тока изменяется МП. Если на поверхности лежит катод, то развивается деполяризация (возбудимость повышается), а если анод - гиперполяризация (возбудимость снижается). Знание механизмов действия электрического тока на живые объекты крайне необходимо для разработки и применения в клинике методов физиотерапии (диатермия, УВЧ, гипергидроз и др.).. Изменение возбудимости при ПД. При локальной ответа возбудимость повышается (ДЭ уменьшается). Изменения возбудимости во время самого ПД можно заметить, если раздражать повторно в разные стадии развития ПД. Оказывается, что во время пика даже очень сильное повторное раздражение остается без ответа (период абсолютной рефрактерности). Затем возбудимость постепенно нормализуется, но она все же ниже, чем начальная (период относительной рефрактерности). При выраженной следовой деполяризации возбудимость выше, чем первоначальная, а при положительной следового потенциала возбудимость снова снижается. Абсолютная рефрактерность объясняется инактивизациею Na + каналов и повышением проводимости К + - каналов. При относительной рефрактерности вновь активизируются Na + - каналы и снижается правиднисть К + - каналов. Двухфазный характер ПД. Обычно в условиях, когда микроэлектрод содержится внутри клетки или волокна, наблюдается однофазный ПД. Иная картина бывает в тех случаях, когда оба электрода лежат на внешней поверхности мембраны - биполярная регистрация. Возбуждение, которое представляет собой волну электроотрицательности, перемещаясь по мембране, доходит сначала до одного электрода, затем помещается между электродами, наконец достигает второго электрода, а затем распространяется дальше. В этих условиях ПД имеет двухфазный характер. Регистрация ПД широко используется в клинике для диагностики заболеваний сердца, мозга, опорно-двигательного аппарата, желудка и др.
Потенциал действия. Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя, в этом участке возникает возбуждение, одним из наиболее важных проявлений которого служит быстрое колебание МП, называемое потенциалом действия (ПД)
При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями секунды, становится заряженным электроотрицательно по отношению к соседнему, покоящемуся участку, т.е. при возбуждении происходит т.н. "перезарядка мембраны". Точные измерения показали, что амплитуда ПД на 30-50 мв превышает величину МП. Причина этого состоит в том, что при возбуждении происходит не просто исчезновение ПП, а возникает разность потенциалов обратного знака, в результате чего наружная поверхность мембраны становится заряженной отрицательно по отношению у ее внутренней стороне.
В ПД принято различать его пик (т.н. спайк - spike) и следовые потенциалы. Пик ПД имеет восходящую и нисходящую фазы. Перед восходящей фазой регистрируется более или менее выраженный т.н. местный потенциал , или локальный ответ. Поскольку во время восходящей фазы исчезает исходная поляризация мембраны, ее называют фазой деполяризации; соответственно нисходящую фазу, в течение которой поляризация мембраны возвращается к исходному уровню, называется фазой реполяризации. Продолжительность пика ПД в нервных и скелетных мышечных волокнах варьирует в пределах 0,4-5,0 мсек. При этом фаза реполяризации всегда продолжительнее.
Кроме пика, в ПД различают два следовых потенциала - следовую деполяризацию и следовую гиперполяризацию. Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность варьирует от нескольких десятков до сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, развивающимися в мышцах и нерве после окончания возбуждения.Следовые потенциалы непостоянны, и в разных тканях могут проявляться только следовой деполяризацией или только следовой гиперполяризацией, последовательность их проявления тоже может быть различной.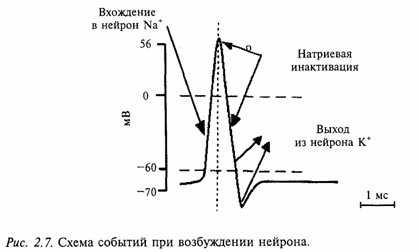
Причиной возникновения ПД является изменение ионной проницаемости мембраны. В состоянии покоя, как уже говорилось, проницаемость мембраны для К+ превышает натриевую проницаемость. Вследствие этого поток положительно заряженных ионов из протоплазмы наружу превышает противоположный поток Na+. Поэтому мембрана в покое снаружи заряжена положительно.
При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко повышается, и в конечном итоге становится примерно в 20 раз больше проницаемости для К+ Поэтому поток ионов Na+ в клетку начинает значительно превышать направленный наружу поток К+. Ток Na+ достигает величины +150 мв. Одновременно несколько уменьшается выход К+ из клетки. Все это приводит к извращению (реверсии) МП, и наружная поверхность мембраны становится заряженной электро отрицательно по отношению к внутренней поверхности. Указанный сдвиг и регистрируется в виде восходящей ветви пика ПД (фаза деполяризации).
Повышение проницаемости мембраны для ионов Na+ продолжается в нервных клетках очень короткое время. Связано оно с кратковременным открытием т.н. Na+-каналов (точнее, заслонок М в этих каналах), которое затем сменяется срочным закрытием Na+-пор с помощью т.н. Н-ворот. Этот процесс называется натриевой инактивацией. В результате поток Na в клетку прекращается.
В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы во внешний раствор. В итоге этих двух процессов происходит восстановление поляризованного состояния мембраны (реполяризация) , и наружная ее поверхность вновь приобретает положительный заряд. В дальнейшем происходят процессы восстановления нормального ионного состава клетки и необходимого градиента концентрации ионов за счет активизации деятельности Na-К-насоса.
Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости мембраны возбудимой клетки. Однако, возбуждение возможно лишь при условии, если действующий на мембрану агент имеет некоторую минимальную (пороговую) величину, способную изменить мембранный потенциал (МП, или Ео ) до некоторого критического уровня (Ек, критический уровень деполяризации). Стимулы, сила которых ниже пороговой величины, называются подпороговыми, выше - надпороговыми. Показано, что пороговая сила, необходимая для возникновения возбуждения при внутриклеточном микроэлектроде равна 10 -7 - 10-9 А.
Таким образом, главным условием для возникновения ПД является следующее: мембранный потенциал должен стать равным или меньше критического уровня деполяризации ( Ео <= Eк)
Закон "все или ничего". ПД подчиняется закону "все или ничего". При изучении зависимости эффектов раздражения от силы приложенного стимула был установлен т.н. закон "все или ничего". Согласно этому закону, подпороговые раздражения не вызывают возбуждения ("ничего"), при пороговых же стимулах возбуждение сразу приобретает максимальную величину ("все"), и уже не возрастает при дальнейшем усилении раздражителя.
Эта закономерность первоначально была открыта Боудичем при исследовании сердца, а в дальнейшем подтверждена и на других возбудимых тканях. Долгое время закон "все или ничего" неправильно интерпретировался как общий принцип реагирования возбудимых тканей. Предполагали, что "ничего" означает полное отсутствие ответа на под пороговый стимул, а "все" рассматривалось как проявление полного исчерпания возбудимым субстратом его потенциальных возможностей. Дальнейшие исследования, в особенности микроэлектродные, показали, что эта точка зрения не соответствует действительности. Выяснилось, что при под пороговых силах возникает местное не распространяющееся возбуждение (локальный ответ). Вместе с тем оказалось, что "все" также не характеризует того максимума, которого может достигнуть ПД. В живой клетке существуют процессы, активно приостанавливающие деполяризацию мембраны. Если каким-либо воздействием на нервное волокно, например, наркотиками, ядами, ослабить входящий Na-ток, обеспечивающий генерацию ПД, то он перестает подчиняться правилу "все или ничего" - его амплитуда начинает градуально зависеть от силы стимула. Поэтому "все или ничего" рассматривается сейчас не как всеобщий закон реагирования возбудимого субстрата на раздражитель, а лишь как правило, характеризующее особенности возникновения ПД в данных конкретных условиях.
studfiles.net
ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
225. К признакам, отличающим возбуждение от раздражения, относятся увеличение уровня потребления кислорода и обменных процессов изменение функции, электрических процессов и энергозатрат.
226. По силе действия раздражители подразделяются на подпороговые, пороговые, надпороговые.
227. По месту возникновения (для клетки или организма) все раздражители делятся на внешние и внутренние.
228. Увеличение мембранного потенциала называется гиперполяризацией.
229. Подпороговый раздражитель оказывает ряд физико-химических сдвигов без видимых эффектов (локальный ответ).
230. Воздействие порогового раздражителя вызывает минимальную видимую ответную реакцию с формированием потенциала действия.
231. Надпороговый раздражитель при формирование потенциала действия согласно закону «все или ничего» вызывает ответную реакцию, однотипную с пороговой.
232. Порог раздражения является способом оценки возбудимости.
233. Большей возбудимостью обладает ткань, для возникновения возбуждения которой необходимо приложить минимальный по силе пороговый раздражитель.
234. Реобазой называется сила раздражителя величиною в 1 порог.
235. Полезное время - это минимальная длительность раздражения возбудимой ткани раздражителем силою в 1 порог, приводящая к развитию потенциала действия.
236. Минимальная сила постоянного тока, вызывающая возбуждение при неограниченно долгом действии, называется реобазой.
237. Минимальное время, в течение которого должен действовать ток двойной реобазы, чтобы вызвать возбуждение называется хронаксией.
238. Сила возбуждения прямо пропорциональна крутизне нарастания силы раздражающего тока.
239. Гидрофильные концы мембрано-образующих молекул обращены наружу.
240. Белки, фиксированные на наружной поверхности мембраны выполняют рецепторную, ферментативную функции, а также функцию ионных каналов утечки и активного транспорта.
241. Суммарный ионный ток через мембрану определяется количеством открытых каналов.
242. Для клеточных мембран высших млекопитающих и человека характерны натриевые, калиевые, кальциевые и другие виды ионных каналов.
243. Электрические явления возбудимых тканей обусловлены электрическими свойствами клеточных мембран.
244. Мембранный потенциал различных возбудимых тканей колеблется в пределах (- 95) - (–70) Мв.
245. Пассивный транспорт ионов через мембрану по градиенту концентрации формирует диффузионные токи.
246. Движение ионов через мембрану по градиенту концентрации, не требующее затраты энергии, называется пассивным транспортом.
247. Движение ионов через мембрану против концентрационного градиента, требующее затраты энергии, называется активным транспортом.
248. Встроенная в клеточную мембрану белковая молекула, обеспечивающая избирательный переход ионов через мембрану с затратой энергии АТФ - это специфический ионный канал.
249. Потенциал мембраны вне состояния возбуждения ткани называется потенциалом покоя.
250. Потенциал мембраны в состоянии возбуждения ткани называется потенциалом действия.
251. Натриевые каналы имеют быстрые активационные и медленные инактивационные «ворота».
252. При действии постоянного тока под катодом происходит деполяризация мембраны, при которой возбудимость мембраны под катодом увеличивается.
253. Разность потенциалов между цитоплазмой и окружающим клетку раствором называется мембранным потенциалом.
254. Внутренняя поверхность мембраны возбудимой клетки по отношению к наружной в состоянии физиологического покоя заряжена отрицательно.
255. Наружная поверхность возбужденного участка клетки (ткани) по отношению к невозбужденному заряжена отрицательно.
256. Уровень деполяризации мембраны, при котором возникает потенциал действия, называется критическим уровнем.
257. Биологический процесс, характеризующийся временной деполяризацией мембран клеток и изменением обменных процессов, называется возбуждением.
258. Уменьшение величины мембранного потенциала покоя при действии раздражителя называется деполяризацией.
259. Увеличение мембранного потенциала покоя называется гиперполяризацией.
260. Концентрация ионов калия в цитоплазме в 30-50 раз больше, чем концентрация снаружи.
261. Концентрация ионов натрия в цитоплазме в 10-20 раз меньше, чем концентрация снаружи.
262. Причиной поляризации являются ионная асимметрия, различная степень диффузных токов, деятельность К-Nа- насоса.
263. Диффузионные токи калия увеличивают величину мембранного потенциала.
264. Величина диффузного тока определяется значением концентрационного градиента, размерами ионов, размерами и структурой мембраны.
265. При увеличении концентрации калия снаружи клетки мембранный потенциал уменьшится.
266. При увеличении концентрации натрия снаружи клетки мембранный потенциал уменьшится.
267. При увеличении концентрации калия внутри клетки мембранный потенциал увеличится.
268. Если поток натрия внутрь клетки увеличится, а поток калия останется прежним мембранный потенциал уменьшится.
269. Гидролиз одной молекулы АТФ для энергетического обеспечения работы NA-K-насоса обеспечивает трансмембранный транспорт против градиента концентрации 2-х ионов натрия и 3-х ионов калия.
270. Воздействие адекватного подпорогового раздражителя приведет к увеличению натриевого тока в клетку.
271. Величина локального ответа в зависимости от силы подпорогового раздражителя подчиняется закону градуальности.
272. Локальный ответ распространяется декрементно (с затуханием).
273. Локальный ответ способен к суммации.
274. Первая фаза потенциала действия называется фазой деполяризации.
275. Вторая фаза потенциала действия называется фазой реполяризации.
276. Первая фаза потенциала действия разовьется в том случае, если уменьшение мембранного потенциала достигнет критического уровня.
277. Снижение мембранного потенциала до критического уровня приводит к лавинообразному натриевому току внутрь клетки.
278. Потенциалу действия предшествует локальный ответ.
279. Величина потенциала действия в нервном волокне равняется 110-120 мВ.
280. Продолжительность основного зубца потенциала действия в нервном волокне равна 0,5-2 мс.
281. Фазы потенциала действия, наступающая за основным зубцом потенциала действия называется следовыми потенциалами.
282. Фаза следовой электроположительности характеризуется тем, что восстанавливающийся мембранный потенциал превосходит величину исходного мембранного потенциала покоя.
283. Формирование следовой электроположительности обуславливает ток ионов калия.
284. В отличие от локального ответа потенциал действия имеет абсолютный рефрактерный период.
285. Период генерации потенциала действия сопровождается выделением 2-3 % тепла в результате сопровождающих процесс возбуждения биохимических процессов.
286. В фазу запаздывающего теплообразования выделяется 97-98 % тепла биохимических процессов, обусловленных процессом возбуждения.
287. Возбудимость ткани в период абсолютной рефрактерности отсутствует.
288. Фаза относительной рефрактерности характеризуется тем, что возникновение потенциала действия в этот период возможно лишь в случае воздействия раздражителя надпороговой величины.
289. Фаза повышенной возбудимости характеризуется тем, что возникновение потенциала действия в этот период возбудимости возможно в случае воздействия раздражителя подпороговой величины.
290. Период повышенной возбудимости в фазу следовой деполяризации называется экзальтацией.
291. Фаза пониженной возбудимости соответствует следовой электроположительности потенциала действия.
292. Физиологическая характеристика возбудимой ткани, отражающая ее способность к воспроизводству максимального количества импульсов в единицу времени называется лабильностью.
293. Лабильность и рефрактерный период друг от друга находятся в обратной зависимости.
294. Лабильность возбудимой ткани по мере развития утомления уменьшается.
295. В условиях физиологического эксперимента с возбудимыми тканями чаще всего используют электрический ток.
296. Свойства электрического тока, которые позволяют применять его в качестве раздражителя для возбудимых тканей, это его близость к естественному раздражителю, возможность дозированного воздействия и высокая воспроизводимость.
297. Полярный закон действия электрического тока проявляется в возбуждение ткани под катодом при замыкании, под анодом при размыкании.
298. Локальный заряд наружной поверхности мембраны возбудимой ткани в случае приложения к ней анода замкнутой электрической цепи увеличится (произойдет гиперполяризация).
299. Способность живой ткани реагировать на любые виды раздражителей носит название раздражимость.
300. Способность клеток под влиянием раздражения избирательно менять проницаемость мембраны для ионов натрия, калия, хлора носит название возбудимость.
301. Минимальная сила раздражителя, необходимая и достаточная для вызова ответной реакции называется пороговой.
302. Амплитуда сокращения одиночного мышечного волокна при увеличении силы раздражения выше пороговой остается без изменений.
303. Закону силы подчиняется целая скелетная мышца.
304. Закону "все или ничего" подчиняется сердечная мышца.
305. Раздражитель, к восприятию которого в процессе эволюции специализировался данный рецептор и вызывающий возбуждение при минимальных величинах раздражения, называется адекватным.
306. Порог раздражения является способом оценки возбудимости ткани.
307. Приспособление ткани к медленно нарастающему по силе раздражителю называется аккомодацией.
308. Потенциал действия в нейроне в естественных условиях возникает в начальном сегменте аксона.
309. Рефрактерность – это понижение или исчезновение возбудимости.
310. Открытый участок мембраны осевого цилиндра шириной около 1мкм, в котором миелиновая оболочка прерывается, носит название перехвата Ранвье.
311. Изолирующую и трофическую функцию в миелинизированном нервном волокне выполняет миелиновая оболочка.
312. Возбуждение в безмиелиновых нервных волокнах распространяется непрерывно вдоль всей мембраны от возбужденного к невозбужденному участку.
313. Возбуждение в миелинизированных нервных волокнах распространяется скачкообразно от перехвата к перехвату.
314. Нервное волокно практически не утомляется.
315. Мембрана клетки очень тонкая, но достаточно прочная оболочка, она состоит из белков, липидов и мукополисахаридов.
316. Бимолекулярный слой липидов является матриксом мембраны.
317. Натриевые, калиевые, кальциевые и хлорные каналы относят к специфическим, потому что эти каналы избирательно пропускают одноименные ионы.
318. Суммарная проводимость для того или иного иона определяется числом одновременно открытых каналов. Канал состоит из транспортной системы и воротного механизма.
319. Ионные каналы подразделяют на специфические и неспецифические.
320. Концентрация ионов натрия на наружной стороне мембраны во много раз больше концентрации натрия в цитоплазме.
321. Неспецифические каналы не меняют свое состояние при электрических воздействиях на мембрану, потому что неспецифические каналы не имеют воротных механизмов и всегда открыты.
322. Специфические каналы не всегда открыты, потому что они имеют воротные механизмы.
323. Поверхностная мембрана возбудимых клеток в покое электрически поляризована, потому что поверхностная мембрана имеет разный электрический потенциал наружной и внутренней поверхности.
324. Ионная асимметрия поддерживается системами активного транспорта.
325. Перенос ионов против их градиентов концентрации осуществляется активным транспортом.
326. Утечка ионов калия увеличивает разность потенциала между средой и аксоплазмой.
327. При соприкосновении тушки лягушки с металлической подставкой (1-й опыт Гальвани) происходит сокращение мышц лапок. потому что соприкосновение приводит к возникновению возбуждения в спинном мозге.
328. Второй опыт Гальвани доказывает существование биопотенциала, потому что постановка опыта исключала использование металлических предметов.
329. Опыт Маттеучи подтверждает существование «животного» электричества в тканях, потому что в одноименном опыте использование второго нервно-мышечного препарата лягушки позволяло продемонстрировать вторичные сокращения мышцы.
330. Локальный ответ способен к суммации. Локальный ответ распространяется декрементно.
331. Причиной формирования потенциала покоя является ионная асимметрия, потому что избыток ионов калия стремится покинуть клетку и удерживается анионами, а избыток натрия снаружи клетки стремится попасть внутрь ее, но удерживается анионами хлора.
332. Периодическое закрытие быстрых натриевых каналов необходимо для формирования потенциалов действия. Сенсор напряжения улавливает степень перезарядки мембраны и включает инактивационную систему. (?)
333. Локальный ответ распространяется декрементно. Величина возникающего локального ответа подчиняется закону градуальности.
infopedia.su
15. ПОРОГОВЫЕ ПОТЕНЦИАЛ, СИЛА, ВРЕМЯ
4.6. ОЦЕНКА ВОЗБУДИМОСТИ КЛЕТКИ. АККОМОДАЦИЯ. ИСПОЛЬЗОВАНИЕ ЭЛЕКТРИЧЕСКОГО ТОКА В МЕДИЦИНЕ
Возбудимость клетки изменяется не только в процессе возбуждения, но и при изменении химического состава внеклеточной жидкости, например, в результате длительной высокой активности клеток, отклонения показателей внутренней среды в патологических случаях. При снижении концентрации ионов Na+ вне клетки этот ион в меньшем количестве входит в клетку, в результате чего снижается ее возбудимость из-за гиперполяризации клетки. Это наблюдается, например, при бессолевой диете, при этом может развиваться мышечная слабость. Повышение внеклеточной концентрации Na+ вызывает противоположный эффект, например усиление тонуса сосудов вследствие возрастания возбудимости нервно-мышечных элементов. Возбудимость различных тканей сама по себе различна — у нервных клеток выше, чем у мышечных, что используется в клинической практике, например, при выяснении причины двигательных нарушений. Показателями состояния возбудимости ткани являются пороговый потенциал, пороговая сила, пороговое время.
А. Пороговый потенциал (AV) — это минимальная величина, на которую надо уменьшить мембранный потенциал покоя, чтобы вызвать возбуждение (ПД). AV и возбудимость клеток находятся в обратных соотношениях: небольшая величина AV свидетельствует о высокой возбудимости клетки. Если, например, уменьшение мембранного потенциала (частичная деполяризация) на 5—
10 мВ вызывает возникновение ПД, то возбудимость клетки высока. Напротив, большой AV (30—40 мВ) свидетельствует о более низкой возбудимости клетки. Однако во всех случаях ПД возникает только при достижении критического уровня деполяризации клеточной мембраны (Екр).
Критический уровень деполяризации Екр, (КУД) — это минимальный уровень деполяризации клеточной мембраны, при которой возникает ПД. Дальнейшее раздражение клетки и искусственное снижение ПП ничего не изменяют в процессе возникновения ПД, поскольку деполяризация клетки, достигнув критического уровня, сама по себе ведет к открытию потенциалзависимых т-ворот Na-каналов, в результате чего Na+ устремляется в клетку, ускоряя деполяризацию независимо от действия раздражителя. Критический уровень деполяризации клеточной мембраны обычно составляет около —50 мВ. При величине ПП, например, —60 мВ (Е0) деполяризация — уменьшение ПП на 10 мВ приведет к достижению Екр (—50 мВ) и возникнет ПД. Если ПП равен —90 мВ, то для вызова ПД надо снизить ПП на 40 мВ. В последнем случае возбудимость клетки значительно ниже.
Таким образом:
ДУ = Е0 - Екр.
Соотношения между AV, Е0 и Е™ показаны на рис. 4.7 — наибольшая возбудимость при наименьшем AV, наименьшая возбудимость при наибольшем AV. AV мало зависит от критического уровня деполяризации (Екр), но существенно — от ПП клетки (Е0), поскольку Екр, как отмечалось выше, — величина довольно постоянная.
Величина ПП изменяется в различных условиях деятельности клетки, вследствие этого колеблется и ее возбудимость, например при изменении концентрации Са2+, рН среды. Когда концентрация Са2+ в среде повышается, клетка становится менее возбудимой, поскольку возрастает мембранный потенциал, вследствие чего Е0 удаляется от Екр, а когда концентрация Са2+ снижается, возбудимость клетки возрастает, так как мембранный потенциал уменьшается, Е0 приближается к Екр. Такое повышение возбудимости лежит в основе синдрома тетании, связанного с дефицитом Са2+ в крови (см. раздел 4.3.1). Изменения содержания ионов Н+ в среде действуют на возбудимость нейронов так же, как изменения концентрации Са2+, что в обоих случаях объясняется изменением
величины Е0. Однако если мембранный потенциал снижается медленно ниже Екр (—50 мВ), например, в условиях гипоксии, при действии миорелаксантов типа сукцинилхо-лина, то клетка становится невозбудимой вследствие инактивации №+-каналов и невозможности достичь Екр.
Несмотря на то что AV является наиболее точным показателям состояния возбудимости клетки, используется он в эксперименте из-за сложности процедуры реже, чем другие показатели. Чаще всего возбудимость оценивается по пороговой силе раздражителя.
Б. Пороговая сила — это наименьшая сила раздражителя, способная вызвать возбуждение (ПД) при неограничении ее действия во времени (рис. 4.8). Сила раздражителя — понятие собирательное, оно отражает степень выраженности раздражающего воздействия стимула на ткань. Например, сила электрического тока выражается в амперах (А), температура среды — в градусах Цельсия (°С), концентрация химического вещества — в миллимолях на 1 л (ммоль/л), сила звука — в децибелах (дБ) и т.д. При использовании в качестве раздражителя электрического тока предложенное определение пороговой силы совпадает с понятием «реобаза». Реобаза — наименьшая сила тока, способная вызвать импульсное возбуждение. Если возбудимость ткани высока, пороговая сила раздражителя мала. Чем выше возбудимость, тем ниже пороговая сила. Большая пороговая сила свидетельствует о низкой возбудимости ткани. При внутриклеточном раздражении пороговая сила электрического тока для различных клеток равна 1(Г7— КГ9 А. При медленно нарастающей силе раздражителя возбуждение может не возникнуть даже при достижении большой
его силы, значительно превосходящей пороговую. Это свидетельствует о том, что возбудимость ткани в таких условиях уменьшается — возникает явление аккомодации.
Аккомодация — это понижение возбудимости ткани и амплитуды ПД вплоть до полного его отсутствия при медленно нарастающем стимуле (малая крутизна). Главной причиной аккомодации является инактивация Na-каналов, возникающая при медленной деполяризации клеточной мембраны — в течение 1 с и более. Клетка теряет возбудимость, если закрывается около 50 % инакти-вационных h-ворот (в покое h-ворота в основном открыты, закрыто около 20 %). Меньшую роль играет активация К-каналов. Возникающая частичная деполяризация клетки при медленно нарастающей силе стимула ведет к уменьшению мембранного потенциала. Поэтому если и возникает ПД при
дальнейшем резком увеличении силы раздражителя, то его амплитуда мала. Аккомодация развивается, например, в клетках ЦНС, когда они деполяризуются при суммации медленно нарастающих синаптических потенциалов. Скорость развития аккомодации у разных тканей различна, она зависит, как и скорость возникновения ПД, от скорости активации и инактивации ионных каналов, в первую очередь инактивации Na-каналов. Таким образом, аккомодация характеризует не возбудимость ткани, а изменение возбудимости ткани при действии плавно нарастающего раздражителя. Поэтому при определении возбудимости ткани в качестве раздражителя необходимо использовать прямоугольные импульсы. В этом случае скорость нарастания стимула и активация Na-каналов опережают скорость аккомодации (инактивации Na-каналов), что и приводит к возникновению ПД. Важным условием, обеспечивающим возникновение возбуждения при действии раздражителя, является его длительность. Поэтому для оценки свойств возбудимой ткани вводится еще одно понятие — пороговое время.
В. Пороговое время — это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение (см. рис. 4.8 — проекция точки А на абсциссу). Пороговое время называют также полезным временем, так как раздражитель обеспечивает деполяризацию только до критического уровня (Екр.). Далее ПД развивается независимо от действия раздражителя, дальнейшее раздражение уже становится ненужным — бесполезным. В эксперименте в клинической практике для оценки свойств возбудимой ткани чаще используют не пороговое время, а хронаксию. Это связано с тем, что определение порогового времени затруднено (пологая часть кривой, см. рис. 4.8). Хронаксия — наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение (см. рис. 4.8 — проекция точки Б на абсциссе). Хронаксия соответствует более крутой части кривой сила — время, она колеблется от сотых долей до сотен миллисекунд. Измерение хронаксии в клинической практике позволяет уточнить характер повреждений мышцы при травмах. В норме определяется фактически хронаксия нервных волокон, так как возбудимость их выше. В случае повреждения нерва и его перерождения определяют истинную хронаксию мышцы, которая намного превышает таковую до травмы.
Взаимозависимость между сверхпороговой силой раздражителя и временем его действия,
необходимым для вызова возбуждения, показана также на рис. 4.8. Кривая в виде гиперболы (кривая Гоорвега—Вейса—Лапика) демонстрирует, что с увеличением сверхпороговой силы раздражителя время его действия, необходимое для вызова возбуждения, уменьшается, и наоборот. Из графика (правая часть) также следует, что, если для получения возбуждения использовать раздражитель по амплитуде меньше реобазы, возбуждение ткани не возникнет даже в том случае, если время его действия будет бесконечно большим. С другой стороны, если для получения возбуждения использовать раздражитель, длительность которого будет меньше некоторого критического интервала (левая часть графика), возбуждение ткани также не будет получено даже в том случае, если сила раздражителя будет бесконечно большой. Поэтому высокочастотный переменный ток (>10 кГц) опасности для организма не несет: при сверхкоротком воздействии на ткань импульс электрического тока дает лишь тепловой эффект, что используется в клинической практике для глубокого прогревания тканей при различных патологических процессах. Электрический ток с частотой от 0,5 до 1 мГц также может использоваться в лечебных целях, в основном для прогревания тканей. Низкочастотный переменный синусоидальный ток (50 Гц) стимулирует возбудимые ткани. Стимулы синусоидального тока частотой 50 Гц большого напряжения опасны для жизни: они могут вызвать фибрилляцию сердца с летальным исходом (относительный рефрактерный период миокарда представляет собой в этом случае фазу повышенной уязвимости).
В учебниках и руководствах по физиологии нередко встречаются термины «порог раздражения» и «порог возбуждения», под которым понимают минимальную интенсивность или минимальную энергию раздражения, способную вызвать возбуждение. Следует заметить, что понятие «интенсивность раздражения» не определено в количественном отношении. Понятие «порог раздражения» или «порог возбуждения» как минимальная энергия без указания времени действия раздражителя тоже не позволяет определить возбудимость ткани, поскольку при длительном действии слабого (подпороговой силы) раздражителя можно израсходовать большое количество энергии, но возбуждения ткани так и не вызвать. В этом случае деполяризация не достигнет критического уровня (EKp.). Кроме того, возникнет явление аккомодации — инактивация Na-каналов, которое, как известно, развивается быстро. Необоснованно также рассматривать термин «пороговая сила» как величину, зависящую от времени ее действия, что нередко допускается. Пороговая сила не может за-
висеть от времени действия — она зависит только от возбудимости ткани. При определении пороговой силы время ее действия не ограничивают. Сверхпороговая сила действительно связана со временем ее действия: чем она больше, тем короче время ее действия, необходимое для вызова возбуждения.
studfiles.net
БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ
Физиологическое состояние живой ткани характеризуется электрическими токами, которые от нее отводятся; это биоэлектрические токи, или биопотенциалы. Различают биотоки трех типов: 1) токи покоя, или клеточные потенциалы, 2) токи, или потенциалы повреждения и 3) токи действия, или акционные токи.
Токи покоя, или клеточные потенциалы отводятся от живых неповрежденных клеток. Когда нет раздражения, а значит и возбуждения, имеется разность потенциалов между наружной поверхностью оболочки (мембраны) живой клетки, заряженной положительно, и внутренней ее поверхностью, или цитоплазмой, заряженной отрицательно. Для отведения потенциала покоя внутрь клетки вводится микроэлектрод, имеющий внешний диаметр менее 0,5 мкм (микрометра, или микрона), а на наружную поверхность клетки накладывается микроэлектрод, имеющий внешний диаметр в 1—4 мкм.
Величина токов покоя в разных клетках различна и достигает 100 же (милливольт) и больше.
Токи, или потенциалы повреждения отводятся от поврежденного и неповрежденного участков живой ткани, так как поврежденный участок заряжен отрицательно, а неповрежденный в отсутствии возбуждения— положительно. Они являются реакцией живой протоплазмы на повреждение и изменяются во времени и пространстве, снижаясь сразу после повреждения от нескольких десятков милливольт до нескольких милливольт.
Токи действия, или акционные токи имеют большое значение для величины возбуждения и его распространения. Различают однофазные и двухфазные токи действия.
Однофазный ток действия отводится от ограниченного участка ткани при очень слабом подпороговом раздражении. Он является низковольтным, местным, нераспространяющимся. Чем больше сила подпорогового раздражителя, тем больше величина однофазного тока действия. Следовательно, однофазный ток действия характеризует очень слабое местное возбуждение с сохранением возбудимости.
Двухфазный ток действия отводится от ткани, раздражаемой пороговыми и надпороговыми раздражителями. При усилении раздражителя от подпорогового до порогового возбуждение возрастает, достигает критической величины, которая в дальнейшем не изменяется и начинает волнообразно распространяться в обе стороны от раздражаемого участка. При этом отводится высоковольтный потенциал, который в нервных волокнах в 10 раз выше порога возбудимости.
Высоковольтный потенциал распространяющегося тока действия, или пик, имеет почти одинаковую высоту (амплитуду) на всем протяжении возбудимой ткани, но высота пика различна у разных тканей и у нервных волокон разного строения и диаметра. Продолжительность отдельного пика также одинакова на всем протяжении возбудимой ткани. Амплитуда и длительность пика изменяются в зависимости от физиологического состояния ткани и скорости проведения возбуждения.
На высоте пика живая ткань находится в состоянии максимального возбуждения, и если нанести в это время повторное раздражение, то оно не вызовет нового возбуждения и не произведет никакого изменения деятельности ткани или органа. Этот период отсутствия эффекта обозначается как абсолютная рефрактерная фаза. Продолжительность этой фазы в нервных волокнах от 0,4 до 2 мсек. После пика регистрируется низковольтный отрицательный следовой потенциал, который отличается от пика неустойчивостью. Во время протекания низковольтного отрицательного потенциала, достигающего всего 0,05 потенциала пика и продолжающегося в нервных волокнах от 1 до 10 мсек, возбудимость постепенно восстанавливается. Это фаза относительной рефрактерности, в которой ткань или орган реагируют на раздражение, но ниже обычного уровня. И, наконец, фаза относительной рефрактерности переходит в фазу повышенной возбудимости, которая обозначается как супернормальный период, или экзальтационная фаза, во время которой орган реагирует на раздражение значительно выше исходного уровня. Эта фаза совпадает с низковольтным положительным следовым потенциалом, составляющим приблизительно 0,002 потенциала пика.Рефрактерность — свойство всех возбудимых тканей, но в разных тканях ее продолжительность различна, например, в скелетной мышце она продолжается больше, чем в нервах, а в сердечной мышце она продолжается еще дольше. Благодаря абсолютной и относительной рефрактерным фазам возбуждение проводится ритмически, отдельными волнами, или импульсами, так как в продолжение абсолютной рефрактерной фазы возбудимость отсутствует. В этой фазе возбуждение переходит в торможение. Следовательно, нерв может проводить не больше 2000 импульсов в секунду, но в естественных условиях при температуре тела он проводит не больше 200 импульсов в секунду.
Лабильность. Н. Е. Введенский обнаружил, что живые ткани обладают свойством лабилыюсти, или функциональной подвижности. В одних тканях возбуждение быстро возникает и быстро прекращается, в других — медленно возникает, медленно протекает и сравнительно долго не прекращается. Лабильность измеряется числом волн возбуждения в одну секунду.
Следовательно, в тканях, обладающих большой или высокой лабильностью, в течение одной секунды проводится значительно больше волн возбуждения, чем в тканях, которым свойственна малая или низкая лабильность.
Торможение. Когда возбуждение, проявляющееся в деятельности ткани или органа, возрастает до критической величины, то при определенных условиях оно переходит в торможение, проявляющееся во временном активном задерживании деятельности ткани или органа. Н. Е. Введенский обнаружил, что переход возбуждения в торможение происходит постепенно, через несколько стадий. Вначале наблюдается уравнительная стадия, во время которой слабые и сильные раздражители вызывают одинаковую по величине деятельность органа или ткани, например, одинаковой высоты сокращение скелетной мышцы. По мере усиления торможения возникает вторая стадия — парадоксальная. В этой стадии сильные раздражители тормозят работу органа, а слабые вызывают возбуждение и, следовательно, работу органа. И, наконец, наступает последняя стадия — тормозная, в которой и слабые и сильные раздражители вызывают торможение. При переходе торможения в возбуждение эти стадии развиваются в обратном порядке.
Явление перехода возбуждения в торможеиие и наоборот Н. Е. Введенский назвал парабиозом.Единство возбуждения и торможения. Возбуждение и торможение, как следует из сказанного, не являются процессами, существующими независимо друг от друга, а представляют собой две взаимосвязанные фазы единого нервного процесса. При определенных условиях раздражения и состояния лабильности ткани возникает возбуждение, а при других условиях раздражения и состояния лабильности ткани возбуждение переходит в торможение. Для каждой ткани существуют наиболее благоприятные ритм, частота и сила раздражения, вызывающие максимально возможное ее возбуждение и деятельность. Н. Е. Введенский назвал наиболее благоприятную частоту раздражения оптимумом частоты, наиболее благоприятную силу раздражения — оптимумом силы, а чрезмерную частоту или силу раздражения, при которой возникает-торможение, он назвал пессимумом частоты и пессимумом силы. В настоящее время различают несколько других видов торможения в нервной системе.
nauka03.ru



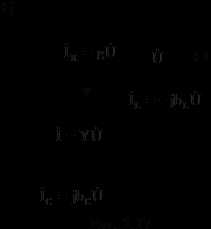





Видеоматериалы
Опыт пилотных регионов, где соцнормы на электроэнергию уже введены, показывает: граждане платить стали меньше
Подробнее...С начала года из ветхого и аварийного жилья в республике были переселены десятки семей
Подробнее...Более 10-ти миллионов рублей направлено на капитальный ремонт многоквартирных домов в Лескенском районе
Подробнее...Актуальные темы
ОТЧЕТ о деятельности министерства энергетики, ЖКХ и тарифной политики Кабардино-Балкарской Республики в сфере государственного регулирования и контроля цен и тарифов в 2012 году и об основных задачах на 2013 год
Подробнее...Предложения организаций, осуществляющих регулируемую деятельность о размере подлежащих государственному регулированию цен (тарифов) на 2013 год
Подробнее...
КОНТАКТЫ
360051, КБР, г. Нальчик
ул. Горького, 4
тел: 8 (8662) 40-93-82
факс: 8 (8662) 47-31-81
e-mail:
Этот адрес электронной почты защищен от спам-ботов. У вас должен быть включен JavaScript для просмотра.
![]()
![]()






