
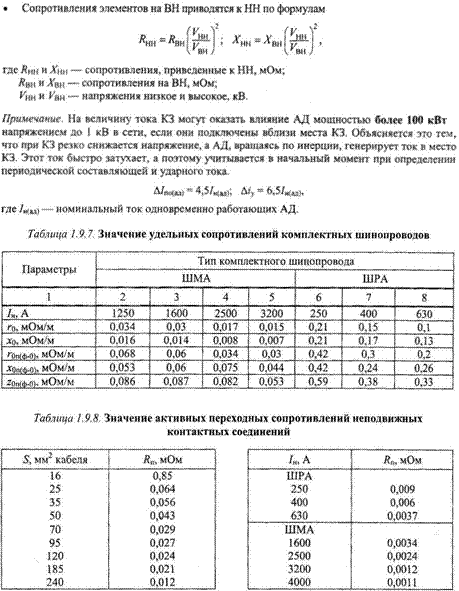
Активные и индуктивные сопротивления проводов
В данной статье представлены справочные таблицы активных и индуктивных сопротивлений воздушных линий с проводами из меди, алюминия и стали взятые из ГОСТ, РД, электротехнических справочников и каталогов производителей.
Активные сопротивления проводов
Значения активных сопротивлений проводов марок М, А, АКП, АН, АЖ, А1, А2, АС, АСца, АСКС, АС КП. АСК АТ1С, АТЗС, АТ4С приведены в ГОСТ 839 – 2019 «Провода неизолированные для воздушных линий электропередач» приложение А, таблицы А1 – А8. Для ознакомления, я приведу лишь несколько таблиц из данного ГОСТа, остальные таблицы вы сможете найти непосредственно в самом ГОСТе.
Значения активных сопротивлений стальных проводов марок ПСТ и ПС приведены в книге «Электроснабжение сельского хозяйства. Будзко А.И. 2000 г.» страница 508.
Индуктивные сопротивления проводов
Значения индуктивных сопротивлений для воздушных линий с проводами из меди, алюминия и стали приведены в РД 153-34.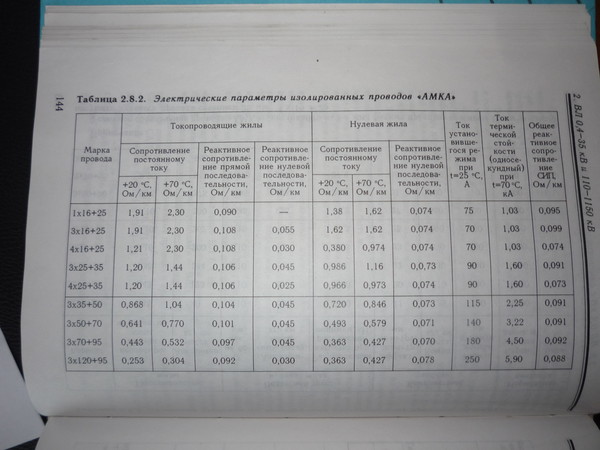 0-20.527-98 «Руководящие указания по расчету токов короткого замыкания и выбору электрооборудования» таблицы П1, П2.
0-20.527-98 «Руководящие указания по расчету токов короткого замыкания и выбору электрооборудования» таблицы П1, П2.
Значения индуктивных сопротивлений стальных проводов марок ПСТ и ПС приведены в книге «Электроснабжение сельского хозяйства. Будзко А.И. 2000 г.» страница 511.
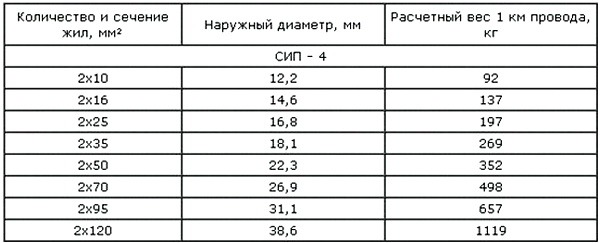
Активные и индуктивные сопротивления проводов СИП-1, СИП-2, СИП-4
Значения активных и индуктивных сопротивлений для проводов СИП-1, СИП-2 и СИП-4 приведены в ТУ 16-705.500-2006 «Провода самонесущие изолированные и защищенные для воздушных линий электропередач» таблицы Б.1, Б.2.
Активные и индуктивные сопротивления проводов СИП-3
Значения активных и индуктивных сопротивлений для проводов СИП-3(SAX-W) приведены в «Пособии по проектированию воздушных линий электропередачи напряжением 0,38 – 20 кВ с СИП. Книга 4» от компании «ENSTO» таблицы 2.6 и 2.7.
Также значение активных сопротивлений для проводов СИП-3 указаны в ГОСТ 31946—2012 таблица 3.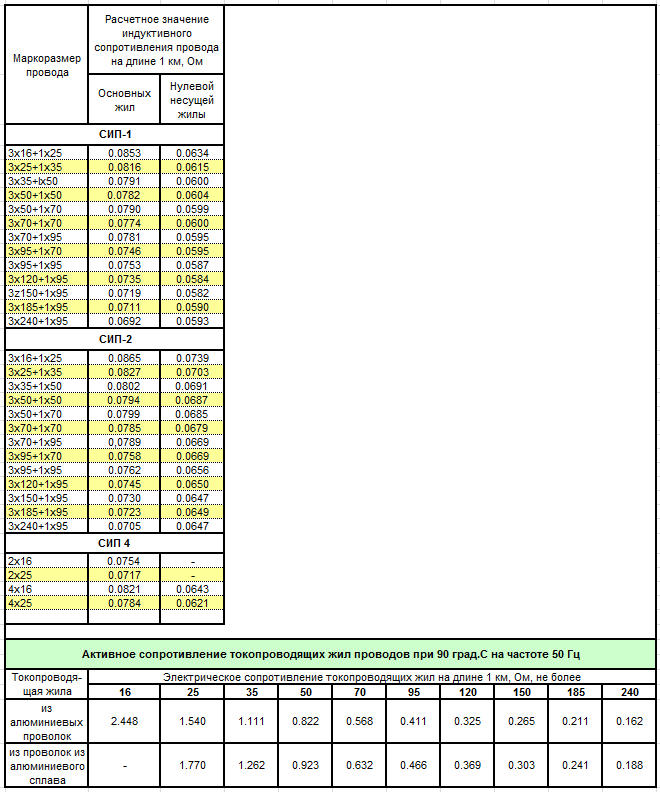 В данной таблице электрическое сопротивление нулевой несущей жилы и токопроводящей жилы указаны при температуре 20 °С.
В данной таблице электрическое сопротивление нулевой несущей жилы и токопроводящей жилы указаны при температуре 20 °С.
Как мы видим значения сопротивлений из пособия компании «ENSTO» таблица 2.6 совпадают с ГОСТ 31946—2012 таблица 3.
Значения индуктивных сопротивлений, приведённые в таблице 2.7 указаны для проводов СИП-3 на напряжение 20 кВ с междуфазным расстоянием 400 мм (данное расстояние указано на установочных чертежах в каталоге).
Соответственно если у вас расстояние между проводами не 400 мм и провода используются свыше напряжения 20 кВ, то применять сопротивления из таблицы 2.7 – я не рекомендую.
В этом случае, ориентировочно индуктивное сопротивление можно рассчитать, по формуле [Л1, с.19]:
где:
- Dср. – среднее геометрическое расстояние между проводами, мм;
- D1-2 — расстояние между проводами первой и второй фазы;
- D2-3 — расстояние между проводами второй и третей фазой;
- D1-3 — расстояние между первой и третей фазой.


Если провода расположены в вершинах равностороннего треугольника со стороной D, имеем Dср = D. Для проводов же, расположенных в одной горизонтальной плоскости и удаленных друг от друга на расстояние D, действительно равенство:
- dр – расчетный диаметр токопроводящей жилы провода без учета изоляции (мм), определяется по ТУ 16-705.500-2006;
Пример
Определить индуктивное сопротивление для проводов марки СИП-3 1х50-20, расположенных в одной горизонтальной плоскости и удаленных друг от друга на расстояние D = 400 мм.
Решение:
1. Определяем среднее геометрическое расстояние между проводами:
где: D = 400 мм – расстояние между проводами.
2. Определяем индуктивное сопротивление для проводов марки СИП-3 1х50-20:
где: dр = 10,7 мм – расчетный диаметр токопроводящей жилы провода без учета изоляции.
Более подробно с самой методикой расчета можно ознакомиться в статье: «Определение активных и индуктивных сопротивлений проводов» .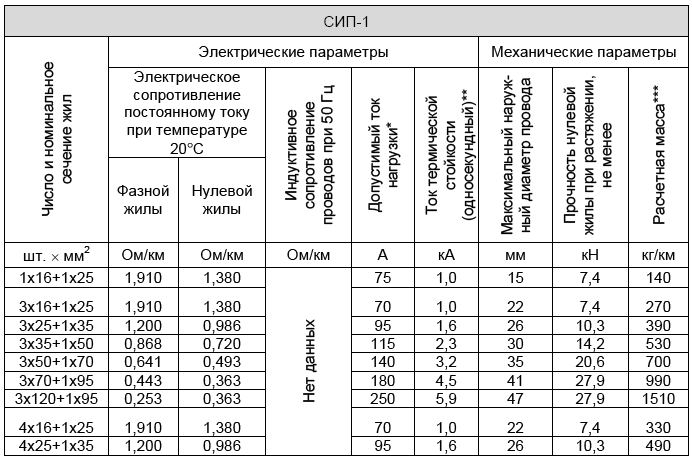
Также рекомендую ознакомиться со статьей: «Пример определения индуктивного сопротивления ВЛ 10 кВ» .
Чтобы уменьшить время на постоянные расчеты индуктивного сопротивления проводов СИП-3, используя формулу, приведенную выше, я предварительно выполнил расчеты для наиболее часто используемых расстояний между проводами 400 – 6000 мм и для всех сечений проводов СИП-3 от 1х35 до 1х240 мм2. Полученные значения индуктивных сопротивлений, я свел в таблицы 1 и 2.
Таблица 1 – Индуктивное сопротивление проводов СИП-3-20 кВ
| Индуктивное сопротивление проводов СИП-3-20 кВ, Ом/км | ||||||||
|---|---|---|---|---|---|---|---|---|
| Среднее геометрическое расстояние между проводами (Dср.), мм | Число и номинальное сечение фазных жил | |||||||
| 1×35 | 1×50 | 1×70 | 1×95 | 1×120 | 1×150 | 1×185 | 1×240 | |
Расчетный наружный диаметр провода, мм (ТУ 16-705. 500-2006 — Таблица 2) 500-2006 — Таблица 2) | ||||||||
| 12 | 13 | 15 | 16 | 18 | 19 | 21 | 24 | |
| ТУ 16-705.500-2006 — Номинальная толщина защитной изоляции защищенных проводов на номинальное напряжение 20 кВ — 2,3 мм, на номинальное напряжение 35 кВ — 3,5 мм. | ||||||||
| Расчетный диаметр токопроводящей жилы без учета изоляции (dр), мм | ||||||||
| 9,7 | 10,7 | 12,7 | 13,7 | 15,7 | 16,7 | 18,7 | 21,7 | |
| 400 | 0,293 | 0,286 | 0,276 | 0,271 | 0,262 | 0,259 | 0,251 | 0,242 |
| 450 | 0,300 | 0,294 | 0,283 | 0,278 | 0,270 | 0,266 | 0,259 | 0,249 |
| 500 | 0,307 | 0,300 | 0,290 | 0,285 | 0,276 | 0,273 | 0,265 | 0,256 |
| 550 | 0,313 | 0,306 | 0,296 | 0,291 | 0,282 | 0,278 | 0,271 | 0,262 |
| 600 | 0,318 | 0,312 | 0,301 | 0,296 | 0,288 | 0,284 | 0,277 | 0,268 |
| 700 | 0,328 | 0,322 | 0,311 | 0,306 | 0,298 | 0,294 | 0,287 | 0,277 |
| 800 | 0,336 | 0,330 | 0,319 | 0,314 | 0,306 | 0,302 | 0,295 | 0,286 |
| 900 | 0,343 | 0,337 | 0,327 | 0,322 | 0,313 | 0,309 | 0,302 | 0,293 |
| 1000 | 0,350 | 0,344 | 0,333 | 0,328 | 0,320 | 0,316 | 0,309 | 0,300 |
| 1250 | 0,364 | 0,358 | 0,347 | 0,342 | 0,334 | 0,330 | 0,323 | 0,314 |
| 1500 | 0,376 | 0,369 | 0,359 | 0,354 | 0,345 | 0,341 | 0,334 | 0,325 |
| 2000 | 0,394 | 0,387 | 0,377 | 0,372 | 0,363 | 0,360 | 0,352 | 0,343 |
| 2500 | 0,408 | 0,401 | 0,391 | 0,386 | 0,377 | 0,374 | 0,366 | 0,357 |
| 3000 | 0,419 | 0,413 | 0,402 | 0,397 | 0,389 | 0,385 | 0,378 | 0,369 |
| 3500 | 0,429 | 0,423 | 0,412 | 0,407 | 0,399 | 0,395 | 0,388 | 0,378 |
| 4000 | 0,437 | 0,431 | 0,420 | 0,415 | 0,407 | 0,403 | 0,396 | 0,387 |
| 4500 | − | − | 0,428 | 0,423 | 0,414 | 0,410 | 0,403 | 0,394 |
| 5000 | − | − | 0,434 | 0,429 | 0,421 | 0,417 | 0,410 | 0,401 |
| 5500 | − | − | − | − | 0,427 | 0,423 | 0,416 | 0,407 |
| 6000 | − | − | − | − | − | − | − | 0,412 |
Как мы видим значение индуктивного сопротивления проводов СИП-3 1х50-20 из расчетной таблицы 1 практически совпало со значением из таблицы 2.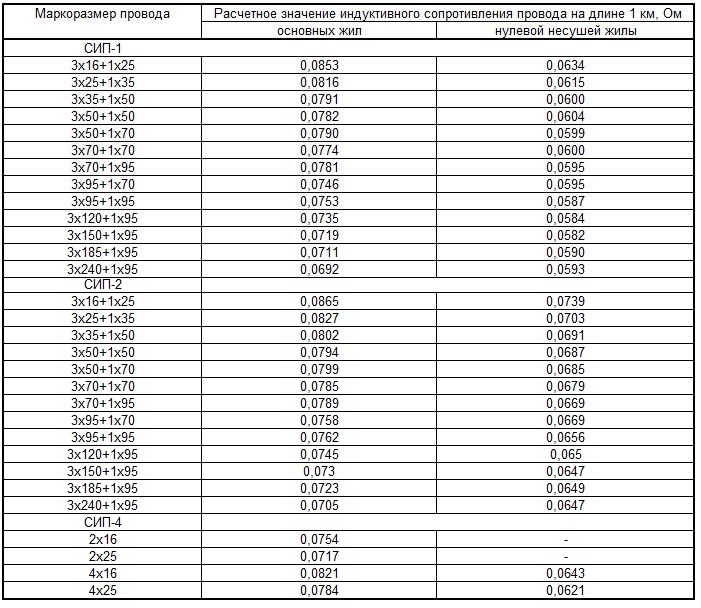 7 компании «ENSTO».
7 компании «ENSTO».
Таблица 2 — Индуктивное сопротивление проводов СИП-3-35 кВ
| Индуктивное сопротивление проводов СИП-3-35 кВ, Ом/км | ||||||||
|---|---|---|---|---|---|---|---|---|
| Среднее геометрическое расстояние между проводами (Dср.), мм | Число и номинальное сечение фазных жил | |||||||
| 1×35 | 1×50 | 1×70 | 1×95 | 1×120 | 1×150 | 1×185 | 1×240 | |
| Расчетный наружный диаметр провода, мм (ТУ 16-705.500-2006 — Таблица 2) | ||||||||
| 14 | 16 | 17 | 19 | 20 | 22 | 24 | 26 | |
ТУ 16-705.500-2006 — Номинальная толщина защитной изоляции защищенных проводов на номинальное напряжение 20 кВ — 2,3 мм, на номинальное напряжение 35 кВ — 3,5 мм.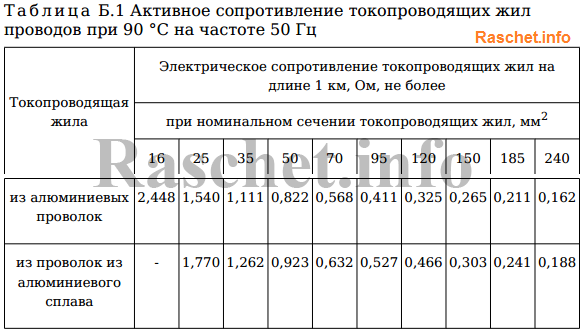 | ||||||||
| Расчетный диаметр токопроводящей жилы без учета изоляции (dр), мм | ||||||||
| 10,5 | 12,5 | 13,5 | 15,5 | 16,5 | 18,5 | 20,5 | 22,5 | |
| 400 | 0,288 | 0,277 | 0,272 | 0,263 | 0,259 | 0,252 | 0,246 | 0,240 |
| 450 | 0,295 | 0,284 | 0,279 | 0,271 | 0,267 | 0,259 | 0,253 | 0,247 |
| 500 | 0,302 | 0,291 | 0,286 | 0,277 | 0,273 | 0,266 | 0,260 | 0,254 |
| 550 | 0,308 | 0,297 | 0,292 | 0,283 | 0,279 | 0,272 | 0,266 | 0,260 |
| 600 | 0,313 | 0,302 | 0,297 | 0,289 | 0,285 | 0,278 | 0,271 | 0,265 |
| 700 | 0,323 | 0,312 | 0,307 | 0,298 | 0,294 | 0,287 | 0,281 | 0,275 |
| 800 | 0,331 | 0,320 | 0,315 | 0,307 | 0,303 | 0,296 | 0,289 | 0,283 |
| 900 | 0,339 | 0,328 | 0,323 | 0,314 | 0,310 | 0,303 | 0,297 | 0,291 |
| 1000 | 0,345 | 0,334 | 0,329 | 0,321 | 0,317 | 0,310 | 0,303 | 0,297 |
| 1250 | 0,359 | 0,348 | 0,343 | 0,335 | 0,331 | 0,324 | 0,317 | 0,311 |
| 1500 | 0,371 | 0,360 | 0,355 | 0,346 | 0,342 | 0,335 | 0,329 | 0,323 |
| 2000 | 0,389 | 0,378 | 0,373 | 0,364 | 0,360 | 0,353 | 0,347 | 0,341 |
| 2500 | 0,403 | 0,392 | 0,387 | 0,378 | 0,374 | 0,367 | 0,361 | 0,355 |
| 3000 | 0,414 | 0,403 | 0,398 | 0,390 | 0,386 | 0,379 | 0,372 | 0,366 |
| 3500 | 0,424 | 0,413 | 0,408 | 0,399 | 0,395 | 0,388 | 0,382 | 0,376 |
| 4000 | 0,432 | 0,421 | 0,416 | 0,408 | 0,404 | 0,397 | 0,390 | 0,384 |
| 4500 | − | − | 0,424 | 0,415 | 0,411 | 0,404 | 0,398 | 0,392 |
| 5000 | − | − | 0,430 | 0,422 | 0,418 | 0,411 | 0,404 | 0,398 |
| 5500 | − | − | − | − | 0,424 | 0,417 | 0,410 | 0,404 |
| 6000 | − | − | − | − | − | − | − | 0,410 |
Литература:
1. Расчет токов короткого замыкания в электросетях 0,4-35 кВ, Голубев М.Л. 1980 г.
Расчет токов короткого замыкания в электросетях 0,4-35 кВ, Голубев М.Л. 1980 г.
Всего наилучшего! До новых встреч на сайте Raschet.info.
Активные и реактивные сопротивления кабелей
|
|
Активные и реактивные сопротивления кабелей
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 448
448 303
303 0816
0816 0746
0746 0865
0865 0669
0669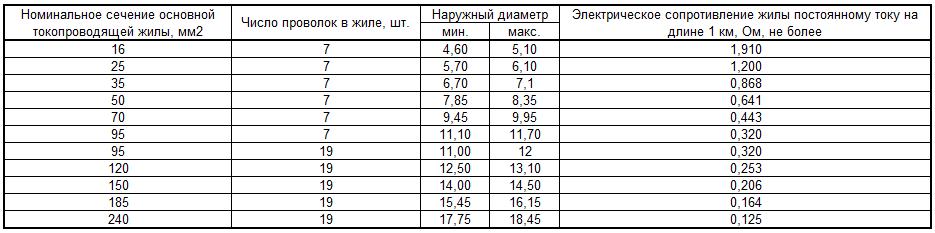 0705
0705Сопротивление цепи фаза – ноль
Таблица 1
Сечение фазных жил мм2 | Сечение нулевой жилы мм2 | Полное сопротивление цепи фаза – ноль, Ом/км при температуре жил кабеля +65 градусов | |||||
Материал жилы: | |||||||
|
| Алюминий | Медь | ||||
|
| R фазы | R нуля | Z цепи (кабеля) | R фазы | R нуля | Z цепи (кабеля) |
1,5 | 1,5 | — | — | — | 14,55 | 14,55 | 29,1 |
2,5 | 2,5 | 14,75 | 14,75 | 29,5 | 8,73 | 8,73 | 17,46 |
4 | 4 | 9,2 | 9,2 | 18,4 | 5,47 | 5,47 | 10,94 |
6 | 6 | 6,15 | 6,15 | 12,3 | 3,64 | 3,64 | 7,28 |
10 | 10 | 3,68 | 3,68 | 7,36 | 2,17 | 2,17 | 4,34 |
16 | 16 | 2,3 | 2,3 | 4,6 | 1,37 | 1,37 | 2,74 |
25 | 25 | 1,47 | 1,47 | 2,94 | 0,873 | 0,873 | 1,746 |
35 | 35 | 1,05 | 1,05 | 2,1 | 0,625 | 0,625 | 1,25 |
50 | 25 | 0,74 | 1,47 | 2,21 | 0,436 | 0,873 | 1,309 |
50 | 50 | 0,74 | 0,74 | 1,48 | 0,436 | 0,436 | 0,872 |
70 | 35 | 0,527 | 1,05 | 1,577 | 0,313 | 0,625 | 0,938 |
70 | 70 | 0,527 | 0,527 | 1,054 | 0,313 | 0,313 | 0,626 |
95 | 50 | 0,388 | 0,74 | 1,128 | 0,23 | 0,436 | 0,666 |
95 | 95 | 0,388 | 0,388 | 0,776 | 0,23 | 0,23 | 0,46 |
120 | 35 | 0,308 | 1,05 | 1,358 | 0,181 | 0,625 | 0,806 |
120 | 70 | 0,308 | 0,527 | 0,527 | 0,181 | 0,313 | 0,494 |
120 | 120 | 0,308 | 0,308 | 0,616 | 0,181 | 0,181 | 0,362 |
150 | 50 | 0,246 | 0,74 | 0,986 | 0,146 | 0,436 | 0,582 |
150 | 150 | 0,246 | 0,246 | 0,492 | 0,146 | 0,146 | 0,292 |
185 | 50 | 0,20 | 0,74 | 0,94 | 0,122 | 0,436 | 0,558 |
185 | 185 | 0. | 0,20 | 0,40 | 0,122 | 0,122 | 0,244 |
240 | 240 | 0,153 | 0,153 | 0,306 | 0,090 | 0,090 | 0,18 |
 20
20
Таблица 2
Мощность трансформатора, кВ∙А | 25 | 40 | 69 | 100 | 160 | 250 | 400 | 630 | 1000 |
Сопротивление трансформатора, Zт/3, Ом (Δ/Υ) | 0,30 | 0,19 | 0,12 | 0,075 | 0,047 | 0,03 | 0,019 | 0,014 | 0,009 |
Таблица 3
I ном. | 1 | 2 | 6 | 10 | 13 | 16 | 20 | 25 | 32-40 | 50 и более |
R авт., Ом | 1,44 | 0,46 | 0,061 | 0,014 | 0,013 | 0,01 | 0,007 | 0,0056 | 0,004 | 0,001 |
 авт. выкл, А
авт. выкл, А
Таблица 4
R цепи, Ом | 0,05 | 0,1 | 0,2 | 0,3 | 0,4 | 0,5 | 0,6 | 0,8 | 1,0 | 1,5 | 2 и более |
Rдуги, Ом | 0,015 | 0,022 | 0,032 | 0,04 | 0,045 | 0,053 | 0,058 | 0,075 | 0,09 | 0,12 | 0,15 |
При проектировании групповой сети, если питающая и распределительная сеть уже проложены, целесообразно выполнить измерение сопротивления цепи фаза – ноль от трансформатора до шин группового щита. Это может значительно уменьшить вероятность ошибок при расчетах групповой сети. В этом случае сопротивление рассчитываем по формуле:
Это может значительно уменьшить вероятность ошибок при расчетах групповой сети. В этом случае сопротивление рассчитываем по формуле:
RL—N= Rрасп + Rпер.гр + Rавт.гр+ Rnгр∙Lnгр +Rдуги (2)
где, Rрасп – измеренное сопротивление цепи фаза – ноль линии, подключаемой к вводному автоматическому выключателю группового щитка, Ом; Rпер.гр – сопротивление переходных контактов в групповой линии, Ом; Rавт.гр – суммарное сопротивление автоматических выключателей – вводного группового щита и отходящей групповой линии, Ом; Rnгр – удельное сопротивление кабеля n-й групповой линии (по таблице 1), Ом/км; Lnгр – длина n-й групповой линии, км.
Рассмотрим процесс вычисления сопротивления цепи фаза – ноль схемы, показанной на Рис.1 при однофазном коротком замыкании фазы на ноль в конце групповой линии.
Исходные данные:
— трансформатор мощностью 630 кВ∙А подключен по схеме «треугольник – звезда» — по таблице 2 находим Zт/3=0,014 Ом;
— питающая сеть – кабель с алюминиевыми жилами длиной 80 метров имеет фазный проводник 150 мм2 и нулевой – 50 мм2. По таблице 1 находим удельное сопротивление кабеля 0,986 Ом/км. Вычисляем его сопротивление (длины кабелей выражаем в километрах): 0,986 Ом/км∙0,08 км=0,079 Ом;
По таблице 1 находим удельное сопротивление кабеля 0,986 Ом/км. Вычисляем его сопротивление (длины кабелей выражаем в километрах): 0,986 Ом/км∙0,08 км=0,079 Ом;
— распределительная сеть – кабель с медными жилами длиной 50 метров и сечением жил 35 мм2. По таблице 1 находим удельное сопротивление кабеля 1,25 Ом/км. Вычисляем его сопротивление:
1,25 Ом/км∙0,05 км=0,0625 Ом;
— групповая сеть – кабель с медными жилами длиной 35 метров и сечением жил 2,5 мм2. По таблице 1 находим удельное сопротивление кабеля 17,46 Ом/км. Вычисляем его сопротивление:
17,46 Ом/км∙0,035 км=0,61 Ом;
— автоматический выключатель отходящий линии – 16 Ампер (с характеристикой срабатывания «С»), вводной автоматический выключатель группового щитка 32 Ампера, остальные автоматические выключатели в линии имеют номинальный ток более 50 Ампер. Вычисляем их сопротивление (по таблице 3) 0,01 Ом+0,004 Ом+3∙0,001 Ом=0,017 Ом;
— переходные сопротивления контактов учтем только в групповой линии (точки подключения кабеля групповой линии к щитку и к нагрузке).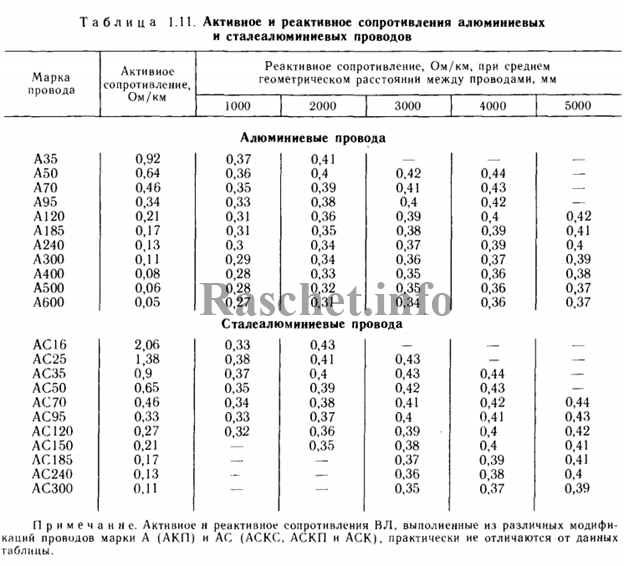 Получаем 2∙0,01 Ом=0,02 Ом.
Получаем 2∙0,01 Ом=0,02 Ом.
Суммируем все полученные значения и получаем сопротивление цепи фаза – ноль без учета сопротивления дуги RL—N=0,014+0,079+0,0625+0,61+0,017+0,02=0,80 Ом.
Из таблицы 4 берем сопротивление дуги 0,075 Ом, и получаем окончательное значение искомой величины RL—N=0,80 Ом+0,075 Ом=0,875 Ом.
В Правилах устройства электроустановок (ПУЭ) задано наибольшее время отключения цепей при коротком замыкании в сетях с глухозаземленной нейтралью 0,2 секунды при напряжении 380 В и 0,4 секунды при напряжении 220В.
Для обеспечения заданного времени срабатывания защиты необходимо, что бы при коротком замыкании в защищаемой линии возникал ток, превышающий не менее чем в 3 раза номинальный ток плавкой вставки ближайшего предохранителя (для взрывоопасных помещений не менее чем в 4 раза) и не менее чем в 3 раза ток расцепителя автоматического выключателя, имеющего обратнозависимую от тока характеристику (для взрывоопасных помещений не менее чем в 6 раз). Для автоматических выключателей с комбинированным расцепителем (имеющим тепловой расцепитель для защиты от перегрузок и электромагнитный расцепитель для защиты от токов коротких замыканий) ток короткого замыкания должен превысить ток срабатывания электромагнитного расцепителя не менее, чем в 1,2 – 1,25 раза.
Для автоматических выключателей с комбинированным расцепителем (имеющим тепловой расцепитель для защиты от перегрузок и электромагнитный расцепитель для защиты от токов коротких замыканий) ток короткого замыкания должен превысить ток срабатывания электромагнитного расцепителя не менее, чем в 1,2 – 1,25 раза.
В настоящее время используются автоматические выключатели с различной кратностью токов срабатывания электромагнитного расцепителя к тепловому. Автоматические выключатели группы «В» имеют кратность в пределах от 3 до 5, группы «С» от 5 до 10, группы «D» от 10 до 20, группы «K» от 10 до 15 и группы «Z» от 2 до 3. При расчетах всегда берется максимальное значение кратности токов срабатывания расцепителей. Например для автоматического выключателя С16, ток короткого замыкания должен быть не менее 16 А∙10∙1,2=192 А (для автоматического выключателя С10 не менее10А∙10∙1,2=120 А и для С25 не менее 25 А∙10∙1,2=300 А). В приведенном выше примере мы получили сопротивление цепи фаза – ноль 0,875 Ом. При таком сопротивлении цепи ток короткого замыкания Iкз составит величину
При таком сопротивлении цепи ток короткого замыкания Iкз составит величину
Uф/ RL—N=220В/0,875 Ом=251 А. Следовательно групповая линия в приведенном примере защищена от токов коротких замыканий.
Максимальное сопротивление цепи фаза – ноль для автоматического выключателя С16 составит величину 220 В/192А=1,14 Ом. В приведенном примере сети (Рис. 1) сопротивление цепи от трансформатора до шин группового щита составит 0, 875 Ом — 0,61 Ом=0.265 Ом. Следовательно максимально возможное сопротивление кабеля групповой линии будет равно 1,14 Ом – 0, 265 Ом=0,875 Ом. Его максимальную длину L при сечении жил кабелей 2,5 мм2 определим при помощи таблицы 1.
L, км=0,875 Ом/(17,46 Ом/км)=0,050 км.
Всегда, когда есть возможность, следует рассчитывать групповую сеть с максимальным запасом по сопротивлению цепи фаза – ноль, особенно розеточную сеть. Часто нагрузки (утюг, чайник и другие бытовые приборы), в которых часто происходят замыкания, подключают к розетке через удлинитель. Начиная с определенной длины провода удлинителя, нарушается согласование параметров цепи с характеристиками аппаратов защиты, то есть ток короткого замыкания оказывается недостаточным для мгновенного отключения сети. Отключение аварийного участка осуществится только тепловым расцепителем через сравнительно большой промежуток времени (несколько секунд), в результате чего кабели могут нагреться до недопустимо высоких температур вплоть до воспламенения изоляции.
Начиная с определенной длины провода удлинителя, нарушается согласование параметров цепи с характеристиками аппаратов защиты, то есть ток короткого замыкания оказывается недостаточным для мгновенного отключения сети. Отключение аварийного участка осуществится только тепловым расцепителем через сравнительно большой промежуток времени (несколько секунд), в результате чего кабели могут нагреться до недопустимо высоких температур вплоть до воспламенения изоляции.
Проект электропроводки должен быть выполнен таким образом, что бы даже в случае воспламенения изоляции кабеля при коротком замыкании это не приводило к пожару. Именно поэтому возникли требования к прокладке скрытой электропроводки в стальных трубах в зданиях со строительными конструкциями, выполненными из горючих материалов. Во взрывоопасных зданиях целесообразно использовать более сложную защиту кабелей от воздействия токов короткого замыкания.
9 марта 2013 г.
К ОГЛАВЛЕНИЮ
Активные и индуктивные сопротивления проводов
В данной статье представлены справочные таблицы активных и индуктивных сопротивлений воздушных линий с проводами из меди, алюминия и стали взятые из ГОСТ, РД, электротехнических справочников и каталогов производителей.
Активные сопротивления проводов
Значения активных сопротивлений проводов марок М, А, АКП, АН, АЖ, А1, А2, АС, АСца, АСКС, АС КП. АСК АТ1С, АТЗС, АТ4С приведены в ГОСТ 839 – 2019 «Провода неизолированные для воздушных линий электропередач» приложение А, таблицы А1 – А8. Для ознакомления, я приведу лишь несколько таблиц из данного ГОСТа, остальные таблицы вы сможете найти непосредственно в самом ГОСТе.
Значения активных сопротивлений стальных проводов марок ПСТ и ПС приведены в книге «Электроснабжение сельского хозяйства. Будзко А.И. 2000 г.» страница 508.
Индуктивные сопротивления проводов
Значения индуктивных сопротивлений для воздушных линий с проводами из меди, алюминия и стали приведены в РД 153-34.0-20.527-98 «Руководящие указания по расчету токов короткого замыкания и выбору электрооборудования» таблицы П1, П2.
Значения индуктивных сопротивлений стальных проводов марок ПСТ и ПС приведены в книге «Электроснабжение сельского хозяйства. Будзко А.И. 2000 г.» страница 511.
Будзко А.И. 2000 г.» страница 511.
Активные и индуктивные сопротивления проводов СИП-1, СИП-2, СИП-4
Значения активных и индуктивных сопротивлений для проводов СИП-1, СИП-2 и СИП-4 приведены в ТУ 16-705.500-2006 «Провода самонесущие изолированные и защищенные для воздушных линий электропередач» таблицы Б.1, Б.2.
Активные и индуктивные сопротивления проводов СИП-3
Значения активных и индуктивных сопротивлений для проводов СИП-3(SAX-W) приведены в «Пособии по проектированию воздушных линий электропередачи напряжением 0,38 – 20 кВ с СИП. Книга 4» от компании «ENSTO» таблицы 2.6 и 2.7.
Также значение активных сопротивлений для проводов СИП-3 указаны в ГОСТ 31946—2012 таблица 3. В данной таблице электрическое сопротивление нулевой несущей жилы и токопроводящей жилы указаны при температуре 20 °С.
Как мы видим значения сопротивлений из пособия компании «ENSTO» таблица 2.6 совпадают с ГОСТ 31946—2012 таблица 3.
Значения индуктивных сопротивлений, приведённые в таблице 2. 7 указаны для проводов СИП-3 на напряжение 20 кВ с междуфазным расстоянием 400 мм (данное расстояние указано на установочных чертежах в каталоге).
7 указаны для проводов СИП-3 на напряжение 20 кВ с междуфазным расстоянием 400 мм (данное расстояние указано на установочных чертежах в каталоге).
Соответственно если у вас расстояние между проводами не 400 мм и провода используются свыше напряжения 20 кВ, то применять сопротивления из таблицы 2.7 – я не рекомендую.
В этом случае, ориентировочно индуктивное сопротивление можно рассчитать, по формуле [Л1, с.19]:
где:
- Dср. – среднее геометрическое расстояние между проводами, мм;
- D1-2 — расстояние между проводами первой и второй фазы;
- D2-3 — расстояние между проводами второй и третей фазой;
- D1-3 — расстояние между первой и третей фазой.
Если провода расположены в вершинах равностороннего треугольника со стороной D, имеем Dср = D. Для проводов же, расположенных в одной горизонтальной плоскости и удаленных друг от друга на расстояние D, действительно равенство:
- dр – расчетный диаметр токопроводящей жилы провода без учета изоляции (мм), определяется по ТУ 16-705. 500-2006;
 500-2006;
500-2006;Пример
Определить индуктивное сопротивление для проводов марки СИП-3 1х50-20, расположенных в одной горизонтальной плоскости и удаленных друг от друга на расстояние D = 400 мм.
Решение:
1. Определяем среднее геометрическое расстояние между проводами:
где: D = 400 мм – расстояние между проводами.
2. Определяем индуктивное сопротивление для проводов марки СИП-3 1х50-20:
где: dр = 10,7 мм – расчетный диаметр токопроводящей жилы провода без учета изоляции.
Чтобы уменьшить время на постоянные расчеты индуктивного сопротивления проводов СИП-3, используя формулу, приведенную выше, я предварительно выполнил расчеты для наиболее часто используемых расстояний между проводами 400 – 6000 мм и для всех сечений проводов СИП-3 от 1х35 до 1х240 мм2. Полученные значения индуктивных сопротивлений, я свел в таблицы 1 и 2.
Таблица 1 – Индуктивное сопротивление проводов СИП-3-20 кВ
| Индуктивное сопротивление проводов СИП-3-20 кВ, Ом/км | ||||||||
|---|---|---|---|---|---|---|---|---|
Среднее геометрическое расстояние между проводами (Dср. ), мм ), мм | Число и номинальное сечение фазных жил | |||||||
| 1×35 | 1×50 | 1×70 | 1×95 | 1×120 | 1×150 | 1×185 | 1×240 | |
| Расчетный наружный диаметр провода, мм (ТУ 16-705.500-2006 — Таблица 2) | ||||||||
| 12 | 13 | 15 | 16 | 18 | 19 | 21 | 24 | |
| ТУ 16-705.500-2006 — Номинальная толщина защитной изоляции защищенных проводов на номинальное напряжение 20 кВ — 2,3 мм, на номинальное напряжение 35 кВ — 3,5 мм. | ||||||||
| Расчетный диаметр токопроводящей жилы без учета изоляции (dр), мм | ||||||||
| 9,7 | 10,7 | 12,7 | 13,7 | 15,7 | 16,7 | 18,7 | 21,7 | |
| 400 | 0,293 | 0,286 | 0,276 | 0,271 | 0,262 | 0,259 | 0,251 | 0,242 |
| 450 | 0,300 | 0,294 | 0,283 | 0,278 | 0,270 | 0,266 | 0,259 | 0,249 |
| 500 | 0,307 | 0,300 | 0,290 | 0,285 | 0,276 | 0,273 | 0,265 | 0,256 |
| 550 | 0,313 | 0,306 | 0,296 | 0,291 | 0,282 | 0,278 | 0,271 | 0,262 |
| 600 | 0,318 | 0,312 | 0,301 | 0,296 | 0,288 | 0,284 | 0,277 | 0,268 |
| 700 | 0,328 | 0,322 | 0,311 | 0,306 | 0,298 | 0,294 | 0,287 | 0,277 |
| 800 | 0,336 | 0,330 | 0,319 | 0,314 | 0,306 | 0,302 | 0,295 | 0,286 |
| 900 | 0,343 | 0,337 | 0,327 | 0,322 | 0,313 | 0,309 | 0,302 | 0,293 |
| 1000 | 0,350 | 0,344 | 0,333 | 0,328 | 0,320 | 0,316 | 0,309 | 0,300 |
| 1250 | 0,364 | 0,358 | 0,347 | 0,342 | 0,334 | 0,330 | 0,323 | 0,314 |
| 1500 | 0,376 | 0,369 | 0,359 | 0,354 | 0,345 | 0,341 | 0,334 | 0,325 |
| 2000 | 0,394 | 0,387 | 0,377 | 0,372 | 0,363 | 0,360 | 0,352 | 0,343 |
| 2500 | 0,408 | 0,401 | 0,391 | 0,386 | 0,377 | 0,374 | 0,366 | 0,357 |
| 3000 | 0,419 | 0,413 | 0,402 | 0,397 | 0,389 | 0,385 | 0,378 | 0,369 |
| 3500 | 0,429 | 0,423 | 0,412 | 0,407 | 0,399 | 0,395 | 0,388 | 0,378 |
| 4000 | 0,437 | 0,431 | 0,420 | 0,415 | 0,407 | 0,403 | 0,396 | 0,387 |
| 4500 | − | − | 0,428 | 0,423 | 0,414 | 0,410 | 0,403 | 0,394 |
| 5000 | − | − | 0,434 | 0,429 | 0,421 | 0,417 | 0,410 | 0,401 |
| 5500 | − | − | − | − | 0,427 | 0,423 | 0,416 | 0,407 |
| 6000 | − | − | − | − | − | − | − | 0,412 |
Как мы видим значение индуктивного сопротивления проводов СИП-3 1х50-20 из расчетной таблицы 1 практически совпало со значением из таблицы 2.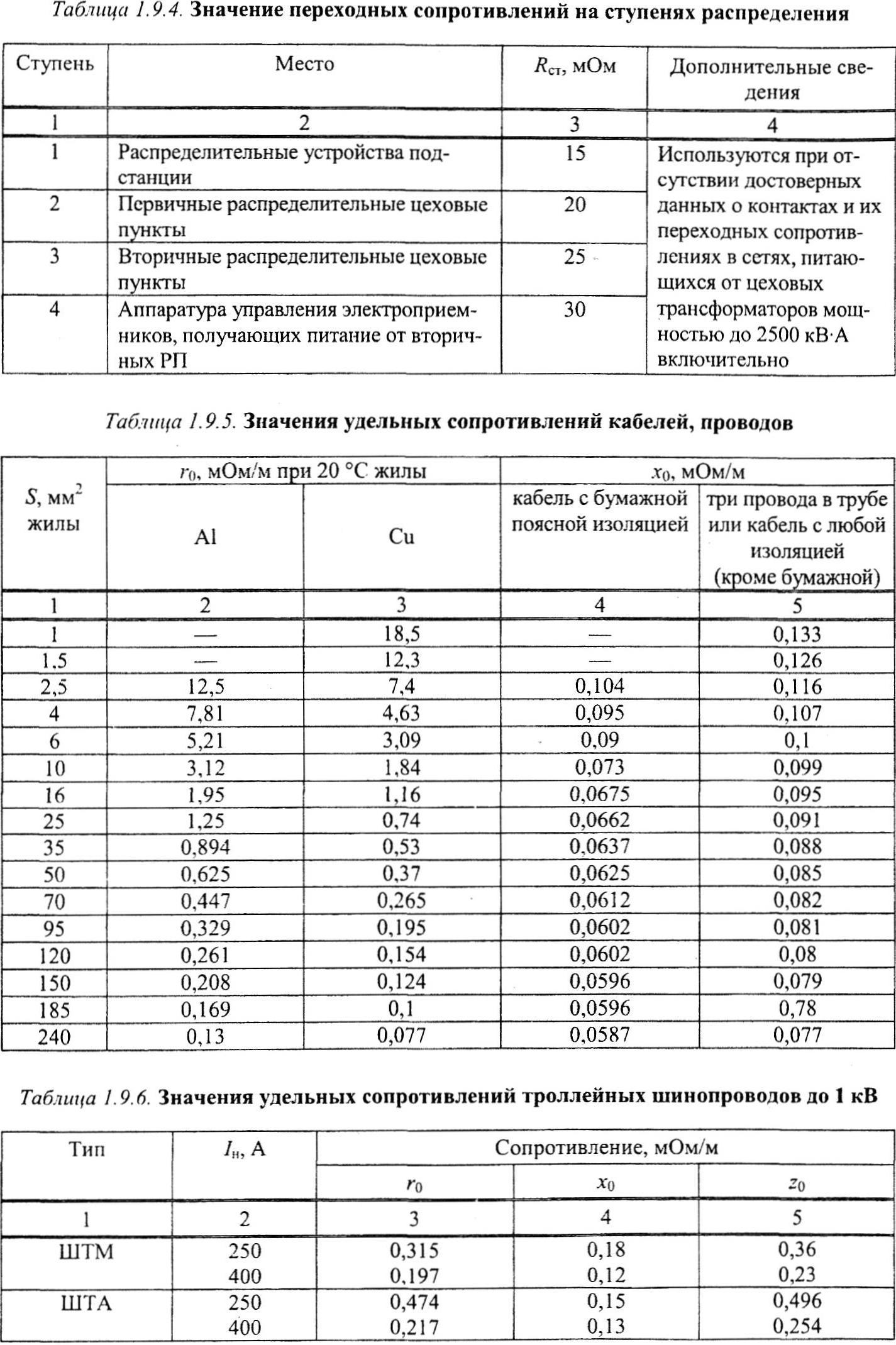 7 компании «ENSTO».
7 компании «ENSTO».
Таблица 2 — Индуктивное сопротивление проводов СИП-3-35 кВ
| Индуктивное сопротивление проводов СИП-3-35 кВ, Ом/км | ||||||||
|---|---|---|---|---|---|---|---|---|
| Среднее геометрическое расстояние между проводами (Dср.), мм | Число и номинальное сечение фазных жил | |||||||
| 1×35 | 1×50 | 1×70 | 1×95 | 1×120 | 1×150 | 1×185 | 1×240 | |
| Расчетный наружный диаметр провода, мм (ТУ 16-705.500-2006 — Таблица 2) | ||||||||
| 14 | 16 | 17 | 19 | 20 | 22 | 24 | 26 | |
| ТУ 16-705.500-2006 — Номинальная толщина защитной изоляции защищенных проводов на номинальное напряжение 20 кВ — 2,3 мм, на номинальное напряжение 35 кВ — 3,5 мм. | ||||||||
| Расчетный диаметр токопроводящей жилы без учета изоляции (dр), мм | ||||||||
| 10,5 | 12,5 | 13,5 | 15,5 | 16,5 | 18,5 | 20,5 | 22,5 | |
| 400 | 0,288 | 0,277 | 0,272 | 0,263 | 0,259 | 0,252 | 0,246 | 0,240 |
| 450 | 0,295 | 0,284 | 0,279 | 0,271 | 0,267 | 0,259 | 0,253 | 0,247 |
| 500 | 0,302 | 0,291 | 0,286 | 0,277 | 0,273 | 0,266 | 0,260 | 0,254 |
| 550 | 0,308 | 0,297 | 0,292 | 0,283 | 0,279 | 0,272 | 0,266 | 0,260 |
| 600 | 0,313 | 0,302 | 0,297 | 0,289 | 0,285 | 0,278 | 0,271 | 0,265 |
| 700 | 0,323 | 0,312 | 0,307 | 0,298 | 0,294 | 0,287 | 0,281 | 0,275 |
| 800 | 0,331 | 0,320 | 0,315 | 0,307 | 0,303 | 0,296 | 0,289 | 0,283 |
| 900 | 0,339 | 0,328 | 0,323 | 0,314 | 0,310 | 0,303 | 0,297 | 0,291 |
| 1000 | 0,345 | 0,334 | 0,329 | 0,321 | 0,317 | 0,310 | 0,303 | 0,297 |
| 1250 | 0,359 | 0,348 | 0,343 | 0,335 | 0,331 | 0,324 | 0,317 | 0,311 |
| 1500 | 0,371 | 0,360 | 0,355 | 0,346 | 0,342 | 0,335 | 0,329 | 0,323 |
| 2000 | 0,389 | 0,378 | 0,373 | 0,364 | 0,360 | 0,353 | 0,347 | 0,341 |
| 2500 | 0,403 | 0,392 | 0,387 | 0,378 | 0,374 | 0,367 | 0,361 | 0,355 |
| 3000 | 0,414 | 0,403 | 0,398 | 0,390 | 0,386 | 0,379 | 0,372 | 0,366 |
| 3500 | 0,424 | 0,413 | 0,408 | 0,399 | 0,395 | 0,388 | 0,382 | 0,376 |
| 4000 | 0,432 | 0,421 | 0,416 | 0,408 | 0,404 | 0,397 | 0,390 | 0,384 |
| 4500 | − | − | 0,424 | 0,415 | 0,411 | 0,404 | 0,398 | 0,392 |
| 5000 | − | − | 0,430 | 0,422 | 0,418 | 0,411 | 0,404 | 0,398 |
| 5500 | − | − | − | − | 0,424 | 0,417 | 0,410 | 0,404 |
| 6000 | − | − | − | − | − | − | − | 0,410 |
Литература:
1. Расчет токов короткого замыкания в электросетях 0,4-35 кВ, Голубев М.Л. 1980 г.
Расчет токов короткого замыкания в электросетях 0,4-35 кВ, Голубев М.Л. 1980 г.
Источник: https://raschet.info
Соединение проводов СИП между собой
В настоящее время частное строительство находится на стадии активного развития. Многие жители нашей страны все чаще предпочитают выполнять строительные и сопутствующие работы самостоятельно, в том числе и проводить электросеть с последующим ее подключением. В связи с этим вопрос о том, как соединить два СИП провода, является актуальным.
СИП – это самонесущий силовой изолированный провод, который часто применяется для монтажа воздушных электрических сетей. В отличие от аналогичных проводов, такой кабель обладает рядом преимуществ:
- безопасность монтажа. СИП имеет уникальную конструкцию и многослойную защиту, благодаря которой достигается абсолютная безопасность при монтаже. Но если вы не имеете специального образования и ряда навыков, то самостоятельный монтаж должен осуществляться со строгим соблюдением технических требований
- надежность. Напряжение передается по кабелю в любых погодных условиях, даже неблагоприятных
- вариативность. Все типы СИП различаются не только размером сечения, но и количеством жил, а также способом их изоляции
- устойчивость к воздействию внешних факторов. Провода изготовлены из специальных материалов, устойчивых к влиянию атмосферных явлений. Даже при очень низкой температуре воздуха они остаются гибкими и мягкими
 Напряжение передается по кабелю в любых погодных условиях, даже неблагоприятных
Напряжение передается по кабелю в любых погодных условиях, даже неблагоприятных
Различные виды СИП могут отличаться размером сечения. Чтобы правильно соединить два провода между собой, необходимо знать их основные характеристики.
Виды СИП проводов
Огромным плюсом использования проводов данного типа является возможность соединения между собой двух и более кабелей различного сечения. Однако специалисты не рекомендуют соединять провода кардинально различных диаметров. Хороший контакт возможен только при соединении двух и более проводов либо одинакового, либо соседнего сечения. Различают СИП провода следующих видов:
- СИП-1 – провода данного типа отличаются тем, что в них все медные жилы, кроме нулевой, изолированы специальным материалом
- СИП-2 – в этих проводах также изолированы все жилы, кроме нулевой. Отличие заключается в способе изоляции – здесь используется материал со специальными молекулярными поперечными сечениями
- СИП-3 – рекомендуется для монтажа электросети в частных домах. Отличительная особенность заключается в конструкции провода – здесь всего одна жила из алюминия, которая изолирована материалом со светостабилизированными частицами. При его покупке специалист рекомендуют спрашивать у продавца сертификат качества
- СИП-4 – характеризуется отсутствием нулевой жилы. Все жилы тщательно изолированы специальным материалом, благодаря чему достигается абсолютная герметичность
- СИП-5 – провода такого типа используются в основном для прокладки электрических сетей на производствах, в городах или между населенными пунктами. Отличительная особенность – каждая жила помещается в специальную коробочку. Самостоятельно соединить провода данного типа практически невозможно
 Отличие заключается в способе изоляции – здесь используется материал со специальными молекулярными поперечными сечениями
Отличие заключается в способе изоляции – здесь используется материал со специальными молекулярными поперечными сечениями
Если вам необходимо соединить два СИП провода между собой, причем вы стремитесь сделать надежный и безопасный во всех отношениях контакт – перед тем, как приступить к действию, проконсультируйтесь со специалистами. Далеко не все провода можно соединять между собой. К примеру, толстый провод с сечением 4 мм очень сложно правильно соединить с кабелем, который имеет сечение 1,5 мм. Здесь необходимо соблюдать определенную последовательность, а любое отступление от технических рекомендаций может привести к проблемам в дальнейшей эксплуатации электросети.
Далеко не все провода можно соединять между собой. К примеру, толстый провод с сечением 4 мм очень сложно правильно соединить с кабелем, который имеет сечение 1,5 мм. Здесь необходимо соблюдать определенную последовательность, а любое отступление от технических рекомендаций может привести к проблемам в дальнейшей эксплуатации электросети.
Способы соединения СИП кабеля
Существует несколько способов соединения двух СИП кабелей разного или одинакового сечения. Самый простой из них – скручивание с последующей спайкой. Чтобы соединить провода между собой, выполните следующие действия:
- зачистите медные жилы обоих проводов (достаточно будет двух-трех см)
- приложите их друг к другу и тщательно скрутите. Обратите внимание, что расстояние между жилами должно быть минимальным. При образовании больших зазоров контакт будет не достаточно качественным
- при помощи специального аппарата осуществите сварку и спайку двух концов
- тщательно изолируйте место соединения
В качестве изоляции можно использовать специальную клейкую ленту или колпачки, которые предназначены специально для этой цели.
Соединить два СИП кабеля между собой можно также с помощью простых болтов. Стоит отметить, что созданное таким способом соединение, по мнению профессиональных электриков, является самым качественным, надежным и долговечным. Процесс соединения отличается простотой – соединить два провода можно самостоятельно в короткие сроки. Для этого необходимо выполнить ряд процедур:
- зачистить жилы обоих проводов. В данном случае длина зачищаемого участка зависит от объема болта
- зачищенную жилу загнуть в форме петли
- петлю надеть на болт
- сверху расположить промежуточную шайбу
Далее на шайбу надевается гайка, которую необходимо тщательно затянуть, а затем проверить прочность соединения.
Есть и другие способы соединения двух СИП проводов, но все они требуют наличия определенных навыков и специальных инструментов, поэтому используют их крайне редко, тем более для прокладки частной электросети.
СИП ПАНЕЛИ
КУПИТЬ СЕРТИФИЦИРОВАННЫЕ СИП ПАНЕЛИ С СИП ЗАВОДА
Свойства и характеристики СИП панелей
Канадские дома очень популярны во многих странах мира. В России тоже все больше людей предпочитают строить дома по SIP технологии. Такой интерес к канадским домам наблюдается благодаря их многочисленным достоинствам и относительно невысокой цене. Основным элементом конструкции таких домов являются СИП панели, и именно их свойства в большой степени определяют достоинства дома. СИП (SIP) панель состоит из двух ОСП (OSB) плит и вклеенного между ними пенополистирола, который выступает в качестве утеплителя. Несложная конструкция и не дорогие материалы, тем не менее, дают строительный материал с уникальным набором свойств. Хорошая теплоизоляция, прочность и легкость панелей вместе с каркасной конструкцией, делают канадские дома очень прочными, теплыми, долговечными и недорогими в строительстве.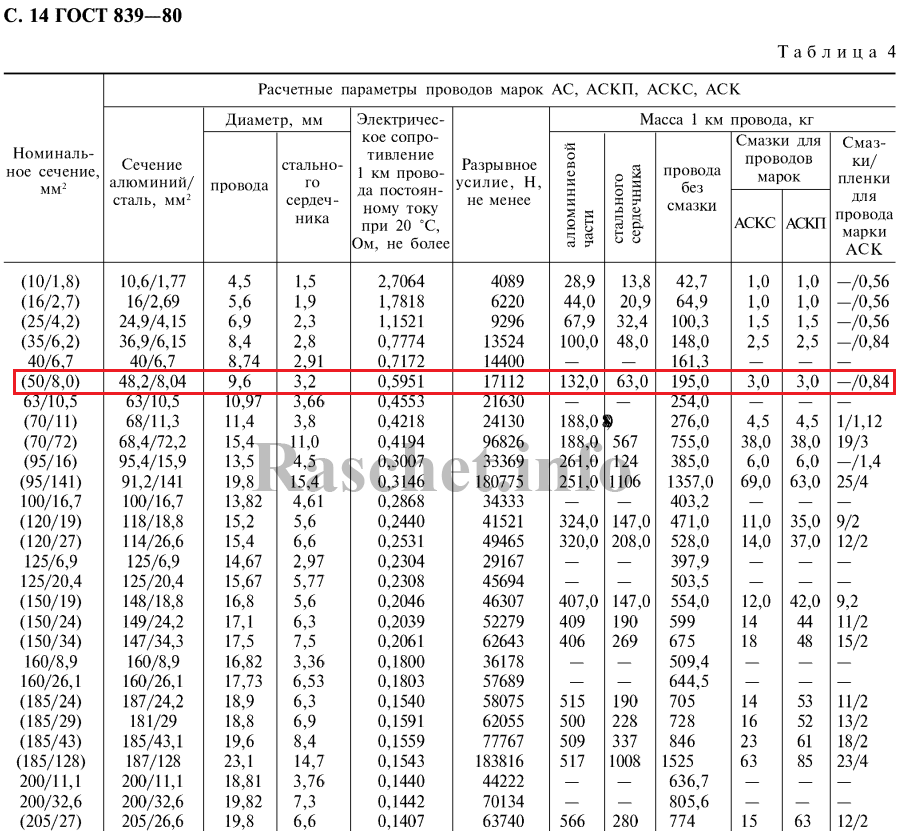
Теплоизоляционные свойства.
Коэффициент теплопроводность пенополистирола составляет 0,037-0,042 Вт/(м·К)). Для примера коэффициент теплопроводности дерева 0,12 Вт/(м·К), пенобетона 0,29 Вт/(м·К), кирпича – 0,7 Вт/(м·К), бетона – 1,51 Вт/(м·К). То есть стена из пенополистирола толщиной 10 см по теплопроводности соответствует стене из дерева толщиной 30 см. Или стене из пенобетона толщиной 73 см или кирпичной стене толщиной 175 см.
Чем меньше теплопроводность тем меньше энергии будет тратиться на обогрев дома и соответственно меньше затрат на энергоносители. В среднем затраты на отопление дома из SIP панелей в 6–8 раз меньше чем на отопление обычных домов (деревянных или кирпичных с утеплением). Сделаем расчет теплового сопротивления СИП панели толщиной 174 мм. (Тепловое сопротивление – величина обратная теплопроводности и используется для тепловых расчетов). Коэффициент теплопроводности пенополистиролва марки ПСБ-С-25 равен 0,039 Вт/(м·К). При толщине 0,15 м его теплопроводность будет составлять 0,039 Вт/(м·К) / 0,15 м = 0,26 Вт/К. Тепловое сопротивление (R) пенополистирола толщиной 15 см будет 1 / 0,26 Вт/К = 3,85 К/Вт. Коэффициент теплопроводности плит OSB-3 равен 0,13 Вт/(м·К). При толщине 24 мм (две плиты по 12мм) теплопроводность будет 0,13 Вт/(м·К) / 0,024м = 5,42 Вт/К, тепловое сопротивление будет 1/ 5,42 Вт/К = 0,18 К/Вт. Тепловое сопротивление СИП панели будет равно R (утеплителя) + R (плит) = 3,85 К/Вт + 0,18 К/Вт = 4,03 К/Вт. Чем больше значение теплового сопротивления, тем меньше происходит теплопотерь, тем “теплее” стены. Согласно строительным нормам для Москвы и области тепловое сопротивление ограждающих конструкций для дома постоянного проживания должно быть не менее 3,13 К/Вт, для административных зданий не менее 2,55 К/Вт. Видно, что СИП панель толщиной 174 мм вполне отвечает требованиям, более того существенно их превосходит.
При толщине 0,15 м его теплопроводность будет составлять 0,039 Вт/(м·К) / 0,15 м = 0,26 Вт/К. Тепловое сопротивление (R) пенополистирола толщиной 15 см будет 1 / 0,26 Вт/К = 3,85 К/Вт. Коэффициент теплопроводности плит OSB-3 равен 0,13 Вт/(м·К). При толщине 24 мм (две плиты по 12мм) теплопроводность будет 0,13 Вт/(м·К) / 0,024м = 5,42 Вт/К, тепловое сопротивление будет 1/ 5,42 Вт/К = 0,18 К/Вт. Тепловое сопротивление СИП панели будет равно R (утеплителя) + R (плит) = 3,85 К/Вт + 0,18 К/Вт = 4,03 К/Вт. Чем больше значение теплового сопротивления, тем меньше происходит теплопотерь, тем “теплее” стены. Согласно строительным нормам для Москвы и области тепловое сопротивление ограждающих конструкций для дома постоянного проживания должно быть не менее 3,13 К/Вт, для административных зданий не менее 2,55 К/Вт. Видно, что СИП панель толщиной 174 мм вполне отвечает требованиям, более того существенно их превосходит.
Физико-механические свойства.
Хотя СИП панели сделаны из древесных плит и пенопласта, тем не менее, они выдерживают достаточно большие нагрузки. Например, SIP панель размерами 2,50х1,25 м и толщиной 174 мм выдерживает поперечную нагрузку 2 тонны. Она прогибается но не разрушается. Такая нагрузка соответствует нагрузке 640 кг/м2. Для сравнения, железобетонные плиты перекрытий, которые используются в строительстве выдерживаю нагрузку порядка 300 – 400 кг/м2. Согласно СНиП-ам расчетная нагрузка на эксплуатируемое перекрытия составляет – 150 кг/м2, расчетная максимальная снеговая нагрузка – 180 кг/м2. Очевидно, что характеристики СИП панели удовлетворяют этим требованиям.
При продольных нагрузках SIP панели выдерживают до 11 тонн на 1 м.п. То есть если взять дом с размерами 10 на 10 метров, то на эти стены можно нагрузить до 440 тонн (периметр стен 40 м.п. * 11 тонн). Такие характеристики с большим запасом перекрывают строительные нормы по нагрузкам на конструкции зданий. Сразу возникает вопрос, откуда такая прочность, ведь панели сделаны из деревоплиты и, смешно сказать, из пенопласта. Да, сама по себе OSB плита достаточно гибкая и даже при не очень больших нагрузках прогибается. Про пенопласт, даже толщиной 150 мм, говорить не приходится. Но при склеивании этих разных по своим механическим свойствам материалов, появляется материал, который по прочности в несколько раз превосходит материалы, из которых он сделан. Структурно СИП можно сравнить с двутавровой балкой. Пенная сердцевина работает как поперечная перегородка, а наружные твердые слои, как аналоги фланцев балки. Чем толще сердцевина, тем лучше панель противостоит поперечному изгибу. При продольном сжатии пенополистирол приклеенный к плитам не позволяет им выгибаться. При поперечном изгибе благодаря пенополистиролу одна из плит SIP панели сжимается, а другая растягивается обеспечивая всей конструкции высокую прочность на изгиб.
Сразу возникает вопрос, откуда такая прочность, ведь панели сделаны из деревоплиты и, смешно сказать, из пенопласта. Да, сама по себе OSB плита достаточно гибкая и даже при не очень больших нагрузках прогибается. Про пенопласт, даже толщиной 150 мм, говорить не приходится. Но при склеивании этих разных по своим механическим свойствам материалов, появляется материал, который по прочности в несколько раз превосходит материалы, из которых он сделан. Структурно СИП можно сравнить с двутавровой балкой. Пенная сердцевина работает как поперечная перегородка, а наружные твердые слои, как аналоги фланцев балки. Чем толще сердцевина, тем лучше панель противостоит поперечному изгибу. При продольном сжатии пенополистирол приклеенный к плитам не позволяет им выгибаться. При поперечном изгибе благодаря пенополистиролу одна из плит SIP панели сжимается, а другая растягивается обеспечивая всей конструкции высокую прочность на изгиб.
Допустимый прогиб панели измеряется следующим образом: длина пролета делится на коэффициент отклонения (L/480, L/360, L/240, L/180, где L – длина пролета).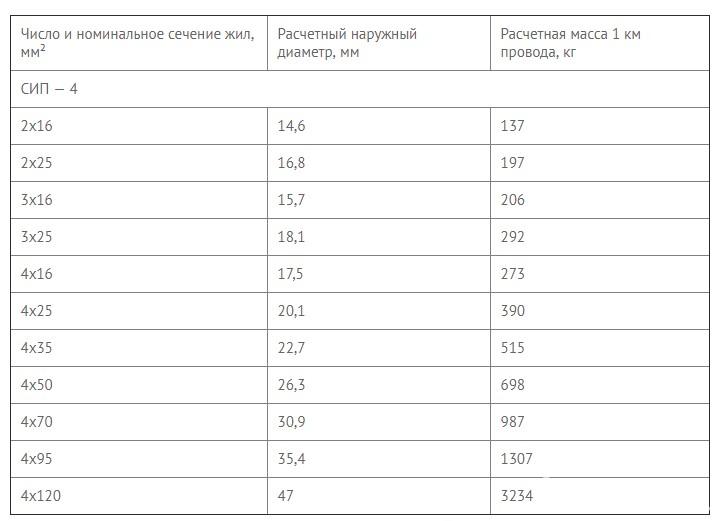 Чем больше знаменатель в этом уравнении, тем меньше прогиб и тем жестче конструкция. Обычно, коэффициент L/360 применяется для расчета несущих способностей перекрытия, а L/240 – для несущих способностей кровли.
Чем больше знаменатель в этом уравнении, тем меньше прогиб и тем жестче конструкция. Обычно, коэффициент L/360 применяется для расчета несущих способностей перекрытия, а L/240 – для несущих способностей кровли.
В таблице приведены данные по нагрузкам для СИП панели толщиной 174 мм.
Прогиб | Длинна пролета | ||||
3050 мм | 3660 мм | 4270 мм | 4880 мм | 5490 мм | |
L/360 | 340 кг/м2 | 236 кг/м2 | 172 кг/м2 | 122 кг/м2 | 95 кг/м2 |
L/240 | 454 кг/м2 | 358 кг/м2 | 259 кг/м2 | 186 кг/м2 | 141 кг/м2 |
Что это означает? Например, если взять пролет 3050 мм, положить на него панель шириной 1 метр, и на каждый квадратный метр плиты положить по 340 кг, то плита прогнется на 8,5 мм ( 3050мм / 360 = 8,5мм)
Особо следует отметить, что в СИП панелях сочетается прочность и одновременно гибкость. Это позволяет домам, построенным из таких панелей выдерживать даже сильные ветровые нагрузки и землетрясения без повреждения конструкции.
Это позволяет домам, построенным из таких панелей выдерживать даже сильные ветровые нагрузки и землетрясения без повреждения конструкции.
Вес панелей.
Не маловажным является, и малый вес СИП панелей. Плотность пенопласта используемого в панелях 25 кг/м3, плотность OSB-3 550 – 650 кг/м3. Поэтому 1 м2 СИП панели толщиной 174 мм весит всего 19 кг. К примеру вес 1 м2 стены из кирпича (толщиной в один кирпич – 25 см) будет весить от 300 до 475 кг. Получается что дом, построенный из СИП панелей приблизительно в 20 раз легче кирпичного. Небольшой общий вес канадских домов позволяет применять облегченные типы фундаментов, а это существенно снижает стоимость строительства. Кроме того вес стандартной панели около 60 кг, а это значит что строительство можно проводить без использования тяжелой строительной техники. Малый вес плиты вместе с ее большими размерами позволяет быстро возводить дома. Все это существенно снижает затраты на строительство.
Пожаростойкость.
Дома, построенные по SIP-технологии, обладают третьей степенью огнестойкости, которая обеспечивает сдерживание огня на протяжении одного часа. SIP-панели обладают свойством самозатухания благодаря входящему в их состав антипирену. Герметичность дома и плотные стыки панелей препятствуют задымлению помещений и распространению огня. При горении SIP-панели выделяют в 7 раз меньше тепловой энергии, чем древесина, и на порядок меньше отравляющих веществ, что снижает опасность удушья в случае нахождения внутри дома.
Экологичность.
СИП панели – это новый для России строительный материал. И у нас до сих пор еще не проводились серьезные и всесторонние испытания этого материала. А отсутствие достоверной информации (на русском языке) приводит к недоверию и многочисленным домыслам, в том числе и о вредности плит OSB и пенополистрола. SIP панели состоят из материалов, удовлетворяющих самым высоким требованиям экологической безопасности. Плита ОСП на 90 % состоит из древесной стружки, остальные 10 % – натуральные смолы, наполнители и отвердители, синтетический воск и соль борной кислоты, которая усиливает защитные свойства ОСП. Обратите внимание, здесь нет фенола, формальдегида, бутанола. Пенополистриол производится из стирола – экологически безопасного вещества, использование которого не ограничено санитарно-гигиеническими нормативами. Этот материал гипоалергенен и в мировом производстве используется даже для изготовления пищевой упаковки и посуды. Следует помнить, что технология производства плит OSB и самих SIP панелей пришла к нам из Северной Америки и Европы, где проблемами экологии серьезно озаботились намного раньше, чем нам стало известно само слово “экология”. Научные учреждения Европы и Америки проводили многочисленные и всесторонние исследования SIP технологии, и все материалы этих исследований опубликованы и доступны, правда, на английском языке. СИП панели не выделяют в атмосферу вредные вещества, поэтому во всём мире их используют для строительства социальных и медицинских учреждений.
Плита ОСП на 90 % состоит из древесной стружки, остальные 10 % – натуральные смолы, наполнители и отвердители, синтетический воск и соль борной кислоты, которая усиливает защитные свойства ОСП. Обратите внимание, здесь нет фенола, формальдегида, бутанола. Пенополистриол производится из стирола – экологически безопасного вещества, использование которого не ограничено санитарно-гигиеническими нормативами. Этот материал гипоалергенен и в мировом производстве используется даже для изготовления пищевой упаковки и посуды. Следует помнить, что технология производства плит OSB и самих SIP панелей пришла к нам из Северной Америки и Европы, где проблемами экологии серьезно озаботились намного раньше, чем нам стало известно само слово “экология”. Научные учреждения Европы и Америки проводили многочисленные и всесторонние исследования SIP технологии, и все материалы этих исследований опубликованы и доступны, правда, на английском языке. СИП панели не выделяют в атмосферу вредные вещества, поэтому во всём мире их используют для строительства социальных и медицинских учреждений.
КУПИТЬ СЕРТИФИЦИРОВАННЫЕ СИП ПАНЕЛИ С СИП ЗАВОДА
R-значений в реальном мире
Изоляция — один из ключевых компонентов любого энергоэффективного дома или коммерческого здания. Поскольку на отопление и охлаждение приходится 50 процентов энергии в среднем доме, выбранный вами тип изоляции может сэкономить тысячи долларов на счетах за коммунальные услуги в течение всего срока службы вашего дома. Изоляция оценивается по R-значению, которое измеряет термическое сопротивление материала. Изоляционный материал с более высоким значением сопротивления R создает более эффективный тепловой барьер между внешней температурой и кондиционируемым пространством внутри дома.
Расчетные значения R для SIP
Но R-значение не раскрывает всей картины. Лабораторные тесты, определяющие значение R, мало похожи на то, как на самом деле работает изоляция в доме. Когда присутствуют реальные факторы, такие как инфильтрация воздуха, экстремальные температуры и тепловые мосты, изоляция из стекловолокна, устанавливаемая на месте, может потерять более половины своего R-значения.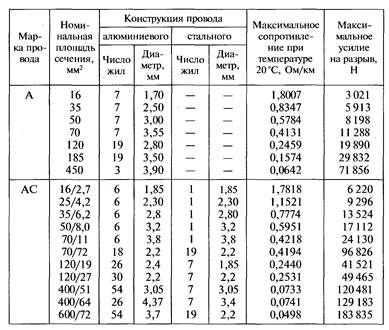 Исследования неоднократно показывали, что SIP обеспечивают непрерывную изоляцию, которая будет поддерживать заявленное значение R в течение всего срока службы дома и каждый раз превосходит изоляцию из стекловолокна.
Исследования неоднократно показывали, что SIP обеспечивают непрерывную изоляцию, которая будет поддерживать заявленное значение R в течение всего срока службы дома и каждый раз превосходит изоляцию из стекловолокна.
Тепловые мосты
Когда в лаборатории измеряется изоляция, установленная на месте, в ходе испытания измеряется только сама изоляция, а не другие компоненты, составляющие систему стен или крыши. Дома с деревянным каркасом полагаются на размерные пиломатериалы, называемые шпильками, через равные промежутки времени, чтобы обеспечить структурную поддержку. Древесина является очень плохим изолятором и образует мост из внешней части дома во внутреннюю часть дома, где тепло может проходить через теплопроводность. Этот процесс известен как тепловой мост.
Еще одна проблема с изоляцией, устанавливаемой на месте, — это сама установка. Стекловолокно необходимо установить между стойками и вырезать, чтобы оно подходило к оконным проемам и проводке. Этот процесс никогда не может быть идеальным и оставляет зазоры там, где отсутствует изоляция.
Национальная лаборатория Ок-Ридж при Министерстве энергетики возглавила строительную отрасль в определении R-значения для всей стены, который становится все более популярным показателем, который проверяет тепловое сопротивление всей секции стены. Стена 2х6 с утеплителем стеклопластик Р-19 получается Р-13.7, когда учитывается тепловое соединение шпилек через каждые 24 дюйма.
Здание с SIP не требует установки какой-либо изоляции в полевых условиях. Для строительства дома из SIP требуется очень мало габаритных пиломатериалов, потому что SIP конструктивно достаточно. Тесты ORNL доказывают, что SIP сохраняют свое полное R-значение при тестировании всей стены.
Эффект инфильтрации воздуха
Усилиям по поддержанию постоянной и комфортной температуры в вашем доме препятствуют две силы: теплопроводность и конвекция.Электропроводность — это передача тепла через твердый материал, для предотвращения чего и предназначена изоляция. Конвекция — это перенос воздуха через щели в стенах и крыше дома.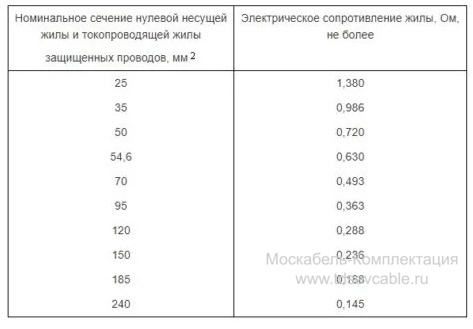 Утечка наружного воздуха в дом или проникновение воздуха является причиной 40 процентов потерь тепла или холода в среднем доме.
Утечка наружного воздуха в дом или проникновение воздуха является причиной 40 процентов потерь тепла или холода в среднем доме.
Изоляция из стекловолокна не защищает от проникновения воздуха, как показали сравнительные испытания, проведенные Национальной лабораторией Окриджа. Исследователи построили два одинаковых объекта площадью 2600 кв.футов домов, один из которых сделан из SIP, а другой — с деревянным каркасом и изоляцией из стекловолокна. Дом для исследования SIP был в 15 раз более воздухонепроницаемым, чем комната с деревянным каркасом, если судить по тесту на дверь с вентилятором.
SIP служат не только как материал для каркаса и изоляции, но и как воздушный барьер, соответствующий нормам. Дома SIP обычно проходят испытания на воздухонепроницаемость в два-три раза больше, чем дома с деревянным каркасом и изоляцией из стекловолокна.
Проникновение воздуха может вызвать больше проблем, чем просто более высокие счета за коммунальные услуги.Воздух, проходящий через изоляцию из стекловолокна, часто переносит влагу.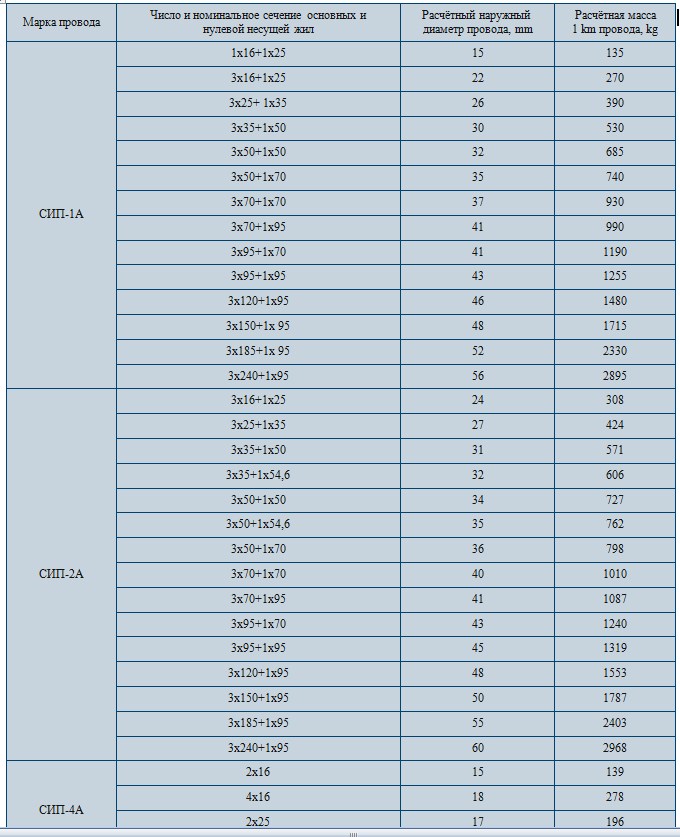 Это может вызвать невидимый рост плесени в полостях стен и общее плохое качество воздуха в помещении, что может привести к проблемам со здоровьем у пассажиров.
Это может вызвать невидимый рост плесени в полостях стен и общее плохое качество воздуха в помещении, что может привести к проблемам со здоровьем у пассажиров.
Лабораторные и реальные условия
В США R-значение изоляции определяется с помощью стандартного метода тестирования, называемого испытанием защищенной горячей пластиной. Этот тест проводится в контролируемой среде, где нет движения воздуха, при температуре 75 ° F.
Исследования, проведенные Окриджской национальной лабораторией Министерства энергетики США, показывают, что по мере того, как наружные температуры становятся холоднее, коэффициент сопротивления стекловолоконной изоляции снижается.Используя полномасштабный климатический симулятор, ORNL проверила изоляцию чердака из стекловолокна с сыпучим заполнением, рассчитанную на R-19, при различных температурах. Когда наружная температура опускалась до -8 ° F, изоляция R-19 работала по R-9,2. Что еще более удивительно, так это то, что инфракрасное изображение показало конвективные токи внутри стекловолоконной изоляции. Теплый воздух изнутри дома будет подниматься через изоляцию, терять тепло, соприкасаясь с холодной температурой чердака, и опускаться обратно через изоляцию, образуя конвективную петлю с постоянной потерей энергии.
Теплый воздух изнутри дома будет подниматься через изоляцию, терять тепло, соприкасаясь с холодной температурой чердака, и опускаться обратно через изоляцию, образуя конвективную петлю с постоянной потерей энергии.
Напротив, изоляция из жесткого пенопласта, используемая в SIP, на самом деле лучше работает при более низких температурах. Пенополистирол с заявленным значением R 3,9 на дюйм при 75 ° F был испытан при R-4,2 на дюйм при 50 ° F и R-4,4 на дюйм при 25 ° F. Что еще более важно, поскольку все типы СИП имеют твердую изоляцию, полностью закрытую деревянной обшивкой, они не подвержены каким-либо конвективным токам, как изоляция из стекловолокна.
Следующее исследование EPSIA подтверждает, что изоляция EPS, обычно используемая в SIP, не теряет R-ценность со временем.
EPSIA R-Value Bulletin
Преимущества SIPs
Из всего, чем должен быть ваш бревенчатый дом — прочным, уединенным, модно обставленным, есть один элемент, который никогда не должен попадать в список: холод. Когда дело доходит до комфорта и энергоэффективности, современные дома из дерева лидируют, во многом благодаря использованию структурных изолированных панелей (SIP), которые закрывают каркас и делают его непроницаемым для погодных условий. Мы поговорили со Стивеном Манном, генеральным директором Insulspan, ведущего производителя структурных изоляционных панелей, чтобы узнать больше о преимуществах SIP.
Когда дело доходит до комфорта и энергоэффективности, современные дома из дерева лидируют, во многом благодаря использованию структурных изолированных панелей (SIP), которые закрывают каркас и делают его непроницаемым для погодных условий. Мы поговорили со Стивеном Манном, генеральным директором Insulspan, ведущего производителя структурных изоляционных панелей, чтобы узнать больше о преимуществах SIP.
THL: Как работают SIP?
SM: SIP создаются путем ламинирования двух кусков OSB2 с высокими эксплуатационными характеристиками (предназначенных для применения в строительстве) вокруг непрерывной сердцевины из пенополистирола (пенополистирола). Мы можем предоставить панели размером до 8 на 24 фута, что позволяет нам покрыть большую площадь одной панелью и уменьшает количество стоек, необходимых для строительства. Это также снижает потери тепла, тепловые мосты и утечку воздуха. Еще одна важная вещь, которую мы делаем с SIP, — это использование герметика на каждом из стыков, который уменьшает воздушный поток. Вы не можете сделать это с помощью стекловолокна или выдувной изоляции.
Вы не можете сделать это с помощью стекловолокна или выдувной изоляции.
THL: Чем SIP по сравнению с другими типами изоляции? Какие преимущества у них перед альтернативами?
SM: Баттовая изоляция имеет R-значение 8,8, но с SIP мы можем получить эффективное R-значение около 22,6. Важным фактором при этом является количество древесины, которое используется для создания стены или крыши. Из-за структурного аспекта SIP требуется не так много пиломатериалов.Пиломатериалы не обладают таким же термическим сопротивлением, как SIP.
Что касается изоляции из напыляемой пены, одно из преимуществ состоит в том, что мы можем производить SIP толщиной до 12,25 дюйма, что обеспечивает R-значение до 45. Если вы действительно хотите увеличить R-значение, вы можете добавить еще 20 процентов. с помощью пенополистирола с усиленным графитом. Вы не можете получить такую густую аэрозольную пену или такую высокую R-ценность. Дело в том, что распыляемая пена создается в результате химической реакции, которая выделяет газ и со временем разрушается, снижая ее R-ценность. EPS — это жесткий пенопласт с закрытыми порами, сделанный из пластика и воздуха, который не разлагается.
EPS — это жесткий пенопласт с закрытыми порами, сделанный из пластика и воздуха, который не разлагается.
THL: Сколько энергии можно сэкономить, используя SIP поверх изоляции из стекловолокна?
SM: Количество сэкономленной энергии будет зависеть от размера здания. Для каркасной стены 2 на 6 с изоляцией из войлока R-20 эффективное значение R составляет около 16,8, а с SIP толщиной 6 1/2 дюйма мы получаем эффективное значение R около 22,6. Но на самом деле это зависит от размера конструкции и толщины стен или поверхностей крыши, которые вы используете.
THL: Сколько стоят SIP по сравнению с другими продуктами, особенно с точки зрения рентабельности инвестиций?
SM: SIP имеют немного более высокие начальные инвестиционные затраты, но если учесть скорость строительства и вам понадобится только один торговый подрядчик вместо двух (каркас и изоляция), покупатели действительно могут сэкономить деньги во время установки. процесс. Однако настоящая экономия — это то, что владельцы увидят в течение всего срока службы дома. При использовании SIP в доме требуется меньшая установка HVAC из-за более высокого значения R и меньшего расхода воздуха, поэтому оборудование стоит меньше.Тогда вы со временем еще больше сэкономите на расходах на электроэнергию благодаря меньшему количеству воздухообмена. Это долгосрочная экономия, которая делает SIP стоящими.
процесс. Однако настоящая экономия — это то, что владельцы увидят в течение всего срока службы дома. При использовании SIP в доме требуется меньшая установка HVAC из-за более высокого значения R и меньшего расхода воздуха, поэтому оборудование стоит меньше.Тогда вы со временем еще больше сэкономите на расходах на электроэнергию благодаря меньшему количеству воздухообмена. Это долгосрочная экономия, которая делает SIP стоящими.
Узнать больше о
Timber Home Living :
Набор толстопленочных резисторов TANCAP TECHNOLOGY SIP-08A 10 кОм ± 2% 200 В 0,125 Вт, шаг 2,54 мм (TANCAP TECHNOLOGY SIP-A08-103G)
Записи с 1 по 6 из 6 Обратите внимание на нашу минимальную сумму заказа 200,00 евро. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SIP-A08-100G TANCAP TECHNOLOGY массив толстопленочных резисторов SIP-08A 10 Ом ± 2% 200 В 0.125 Вт шаг 2,54 мм |
| на складе: 0 шт. 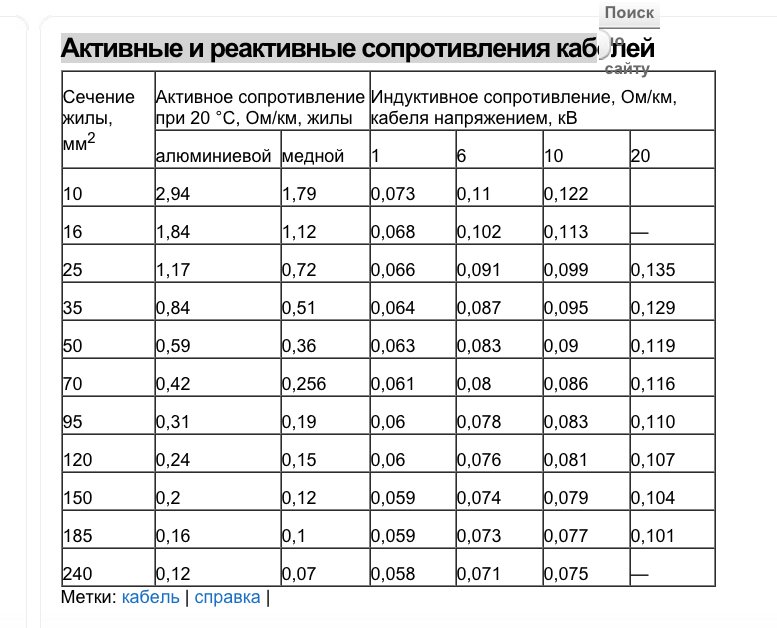 |
| в спецификациюновая спецификация | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SIP-A08-220G TANCAP TECHNOLOGY массив толстопленочных резисторов SIP-08A 22 Ом ± 2% 200 В 0. |
| на складе: 0 шт. |
| в спецификациюновая спецификация | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SIP-A08-102G TANCAP TECHNOLOGY массив толстопленочных резисторов SIP-08A 1 кОм ± 2% 200 В 0. |
| на складе: 0 шт. |
| в спецификациюновая спецификация | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SIP-A08-222G TANCAP TECHNOLOGY массив толстопленочных резисторов СИП-08А 2. |
| на складе: 0 шт. |
| в спецификациюновая спецификация | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SIP-A08-332G TANCAP TECHNOLOGY массив толстопленочных резисторов СИП-08А 3. |
| на складе: 0 шт. |
| в спецификациюновая спецификация | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SIP-A08-104G TANCAP TECHNOLOGY массив толстопленочных резисторов SIP-08A 100 кОм ± 2% 200 В 0. |
| общий запас: 970 шт. |
| в спецификациюновая спецификация | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 125 Вт шаг 2,54 мм
125 Вт шаг 2,54 мм  125 Вт шаг 2,54 мм
125 Вт шаг 2,54 мм 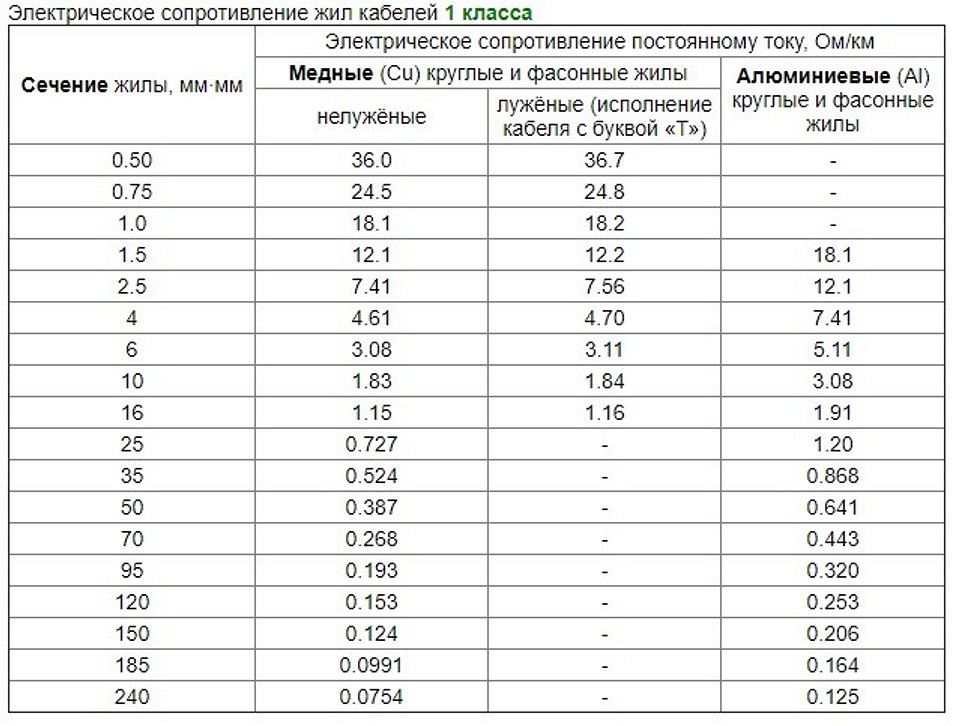 2 кОм ± 2% 200 В 0,125 Вт шаг 2,54 мм
2 кОм ± 2% 200 В 0,125 Вт шаг 2,54 мм  3 кОм ± 2% 200 В 0,125 Вт шаг 2,54 мм
3 кОм ± 2% 200 В 0,125 Вт шаг 2,54 мм  125 Вт шаг 2,54 мм
125 Вт шаг 2,54 мм  0641 *
0641 *Значение R для SIP Insulspan | Сравните R-значения изоляции
Энергоэффективность изоляции обычно измеряется ее значением R. R-значение относится к способности данного материала сопротивляться теплопередаче. Чем выше значение R, тем более стойким является материал, тем меньше энергии вы потребляете и тем меньше ваши счета за электроэнергию.
Перейти к рейтингам Insulspan
Стабильно ли значение R со временем?
При выборе изоляционного материала важно понимать, что существует индивидуальная R-ценность материала, такого как рулон стекловолокна или SIP-панель. Затем существует R-значение для всей стены или крыши после фактической установки продукта (эффективное R-значение).
Начальные значения сопротивления изоляции
Большинство производителей указывают индивидуальную R-ценность своих изоляционных материалов. Это значение R представляет собой значение R этого элемента во время установки в идеальных условиях. Например, если вы покупаете рулон изоляционного материала из стекловолокна R-19, это число относится к наилучшему возможному значению R, предлагаемому этим рулоном изоляции. Однако, как только аккумулятор установлен, значение сразу же начинает снижаться.
Это значение R представляет собой значение R этого элемента во время установки в идеальных условиях. Например, если вы покупаете рулон изоляционного материала из стекловолокна R-19, это число относится к наилучшему возможному значению R, предлагаемому этим рулоном изоляции. Однако, как только аккумулятор установлен, значение сразу же начинает снижаться.
Изоляция из стекловолокна
работает вместе с воздухом, поэтому любое сжатие войлока снижает его эффективность. Ширина войлока R-19 указана как 6¼ дюймов.Рекомендуется для использования с каркасной стеной 2 ″ x 6 ″. Однако шпильки размером 2 x 6 дюймов на самом деле имеют номинальный размер около 5½ дюймов. Так как стойки имеют ширину, которая на дюйма меньше, чем ширина войлока, установка гипсокартона сжимает изоляцию войлока в полости стены. Это уменьшает воздух в войлоке и, как следствие, R-значение.
Перейдите ниже, чтобы увидеть начальные значения R для Insulspan SIP. Это послужит отправной точкой для сравнения наших термических сопротивлений.
Установка имеет значение
Изоляция может быть снижена из-за неправильной техники монтажа.Чрезмерное сжатие войлока, плохое смешивание распыляемой пены и зазоры между стойками — распространенные проблемы. Использование панелей Insulspan EPS практически исключает возможность уменьшения значения R во время установки, поскольку изоляция из пенополистирола (EPS) непрерывно ламинируется на оба листа ориентированно-стружечных плит (OSB).
Тепловые мосты влияют на значения R
Когда мы принимаем во внимание конструкцию всей стены, это называется «эффективным R-значением».«Давайте посмотрим на участок стены, который включает изоляцию и окружающие гвоздики размером 2 x 4 или 2 x 6 дюймов традиционного дома, построенного из палки. Тепловые мосты располагаются в любом месте, где происходит разрыв изоляции в системе стен или крыши. Шпильки — это мосты холода, и в доме с рамой из палочек они встречаются через каждые 16 или 24 дюйма по центру.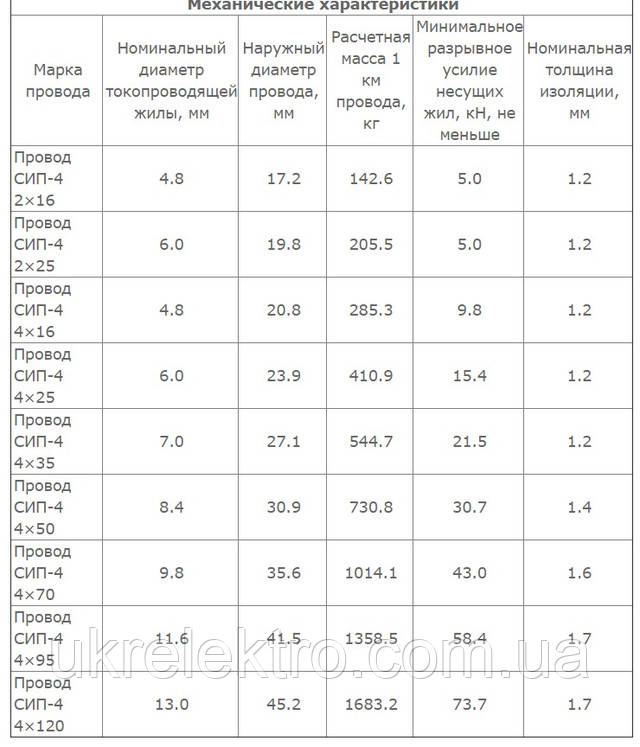 Таким образом, эффективное значение R для стены размером 2 x 6 дюймов со всеми ее тепловыми разрывами составляет примерно 15,62, что меньше, чем автономное значение R для изоляции, помещенной между стойками.
Таким образом, эффективное значение R для стены размером 2 x 6 дюймов со всеми ее тепловыми разрывами составляет примерно 15,62, что меньше, чем автономное значение R для изоляции, помещенной между стойками.
Lumber до сих пор используется для соединения панелей EPS SIP друг с другом. Однако Insulspan производит структурные изолированные панели размером до 8 x 24 футов. В результате в здании Insulspan SIP гораздо меньше тепловых мостов, чем в доме с традиционным каркасом.
Время тоже может повлиять на эффективные значения R
После того, как структура EPS SIP будет завершена, эффективное значение R будет оставаться постоянным с течением времени. Это не относится к другим изоляционным материалам.R-ценность стены уменьшается, когда ватин падает с течением времени из-за силы тяжести. Распылительная пена, в которой в качестве вспенивателя используется термостойкий материал, будет выделять этот агент, теряя большую часть этой дополнительной изоляционной ценности в течение 5 лет. Узнайте больше об эффективном тепловом сопротивлении в PIB 209.
Узнайте больше об эффективном тепловом сопротивлении в PIB 209.
Insulspan Thermal R-значения
Insulspan предлагает две линейки продуктов SIP: SIP с традиционной белой изоляцией EPS и SIP с серым GPS (расширяемый полистирол с графитом), называемый изоляцией R-Plus.В следующих таблицах показано тепловое сопротивление (R-значение) обоих этих продуктов. SIP-панели Insulspan R-Plus предлагают домовладельцам и строителям все преимущества пенополистирола с более высокими значениями R и более тонкими панелями. Нужны подробности? Отъезд: PIB 202 R-Value & U-Factor EPS SIP и PIB 220 R-Value & U-Factor GPS SIP
| Белый EPS | Серый GPS (Neopor) | |
|---|---|---|
| Толщина панели | Тепловое значение R | Тепловое значение R |
| 4 1/2 | 15 | 18.3 |
| 6 1/2 | 22,6 | 27,7 |
| 8 1/4 | 29,2 | 35,9 |
| 10 1/4 | 36,8 | 45,3 |
| 12 1/4 | 44,4 | 54,7 |
Что такое U-фактор?
В то время как R-значение материала измеряет тепловое сопротивление, U-фактор (U-value) измеряет скорость теплопередачи через материал.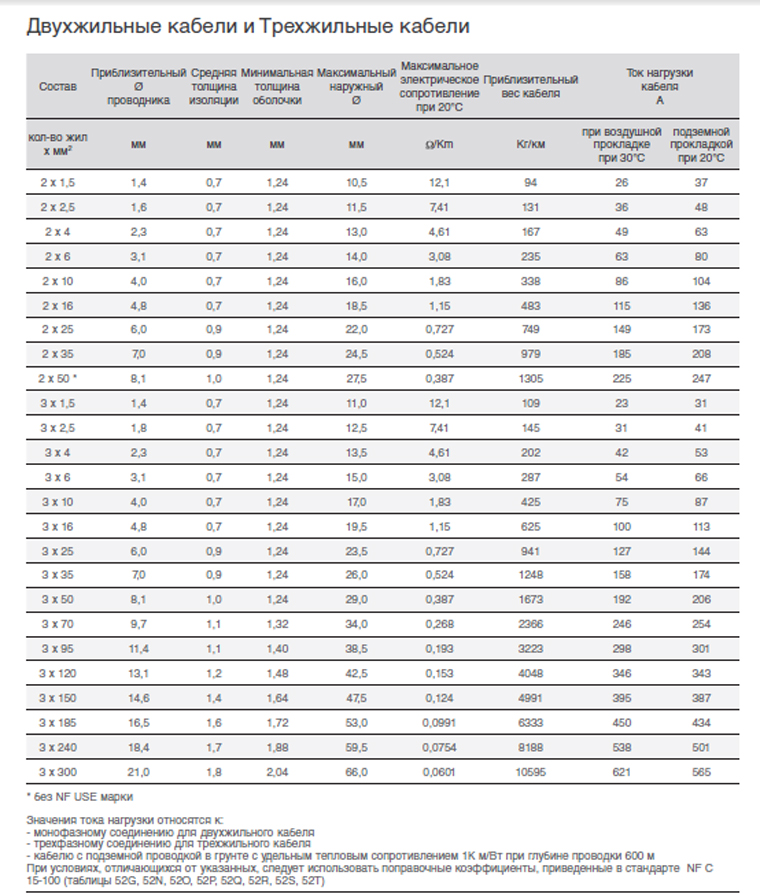 Коэффициент теплопередачи учитывает теплопередачу за счет теплопроводности, конвекции и излучения. Например, энергоэффективность окна обычно указывается по U-значению, потому что конфигурация окна должна учитывать все три метода передачи: проводимость энергии через такие материалы, как стекло или кожух, конвекция через любые незапечатанные части окна. окна, и излучение (от солнца) через остекление, нанесенное на окна.
Коэффициент теплопередачи учитывает теплопередачу за счет теплопроводности, конвекции и излучения. Например, энергоэффективность окна обычно указывается по U-значению, потому что конфигурация окна должна учитывать все три метода передачи: проводимость энергии через такие материалы, как стекло или кожух, конвекция через любые незапечатанные части окна. окна, и излучение (от солнца) через остекление, нанесенное на окна.
Электронный глоссарий; Сеть с двумя оконечными резисторами, схема SIP
Общий технический словарь
«А»
«B»
«C»,
«D»,
«E»,
«F»,
«Г»,
«ЧАС»,
«Я»,
«J»,
«К»,
«L»,
«М»,
«Н»,
«О»,
«П»,
«Q»,
«Р»,
«S»,
«Т»,
«U»,
«V»,
«W»,
«ИКС»,
«Y»,
«Z»
Сети с двойным оконечным резистором
Резисторные сети
Двойные оконечные резисторы [Резисторная сеть]
Резисторная сеть, параллельное соединение [Thevenin]
Параллельная или двойная оконечная сеть Обычное использование; окончание шины.Обычно контакт 1 подключается к земле, а контакт с самым большим номером контакта подключается к цепи питания [Vcc].
Оконечная нагрузка параллельного резистора
Сеть параллельного соединения [Line Terminator] будет одним из немногих сетевых пакетов, которые будут содержать два резистора разного номинала. Как показано на изображении пакета выше; Резистор R1 будет иметь одно значение, а резистор R2 — другое значение. Есть много различных комбинаций значений, которые относятся к стандартным схемам оконечной нагрузки шины.
См. Таблицу справа, чтобы увидеть стандартные значения для R1 и R2. Другие возможные общие значения не показаны. Также обратитесь к определению терминатора резистора, которое является частью раздела терминов резистора.
Внешний вид и размеры пакета резистора SIP
Существует два основных стиля пакетов SIP; формованная упаковка [показанная выше] или упаковка с конформным покрытием. Оба пакета отображаются на странице пакетов SIP.
Связанные компоненты ;
Стиль SIP; [сквозное отверстие]
Схема сети с шинным резистором, SIP, корпус с однопроводным соединением.
Схема сети с изолированным резистором, SIP, корпус с одинарным входом
Схема сети нестандартных резисторов, 6-контактный SIP
Тип DIP; [сквозное отверстие]
Схема резисторной сети, DIP, двухрядный корпус.
RC Networks, DIP, двухрядный корпус.
Стиль LLCC; [поверхностный монтаж]
20-контактная схема сети LLCC, бессвинцовый держатель микросхемы.
Схема 16-контактной сети LLCC, бессвинцовый держатель микросхемы.
6-контактная сетевая схема LLCC, несущий элемент для двух безвыводных микросхем.
Схема 36-контактного массива BGA, шаровая сетка.
Размеры корпуса SIP с 10 контактами
Выше показаны 8-контактный SIP и 10-контактный SIP, но не показан 6-контактный SIP. Однако рисунок 6-контактного резистора SIP доступен на другой странице.
MIL-PRF-83401 ; Резисторные сети, фиксированные, пленочные и конденсаторно-резистивные сети, керамические конденсаторы и фиксированные пленочные резисторы, общие технические условия
In vivo Pyro-SIP Оценка активной кишечной микробиоты хлопкового листового червя Spodoptera littoralis
Abstract
Микробиота кишечника имеет решающее значение для организма-хозяина со значительной метаболической активностью.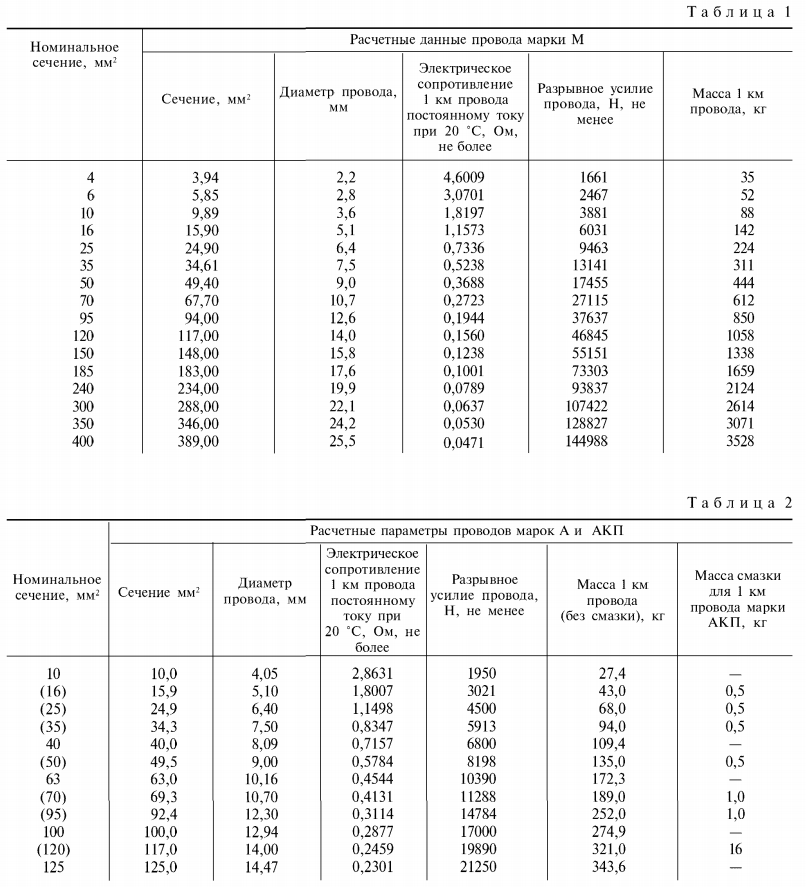 Несмотря на то, что были предприняты большие усилия для характеристики микробного разнообразия, измерение метаболической активности компонентов на удивление не успевает за этим. Здесь мы объединили пиросеквенирование амплифицированных генов 16S рРНК с зондированием in vivo и стабильного изотопа (Pyro-SIP), чтобы выявить метаболически активные бактерии в кишечнике хлопкового листового червя ( Spodoptera littoralis ), многоядного насекомого-травоядного, потребляющего большое количество растений. материал за короткое время, высвобождая обильное количество глюкозы в пищеварительном тракте как наиболее важном источнике углерода и энергии как для хозяйских, так и для активных кишечных бактерий.Используя глюкозу 13 C в качестве трофического звена, Pyro-SIP показал, что относительно простая, но отличительная кишечная микробиота развивалась совместно с хозяином, и метаболическая активность, и состав менялись на протяжении личиночных стадий. Pantoea , Citrobacter и Clostridium были особенно активны в раннем возрасте, вероятно, основные функциональные популяции, связанные с улучшением питания.
Несмотря на то, что были предприняты большие усилия для характеристики микробного разнообразия, измерение метаболической активности компонентов на удивление не успевает за этим. Здесь мы объединили пиросеквенирование амплифицированных генов 16S рРНК с зондированием in vivo и стабильного изотопа (Pyro-SIP), чтобы выявить метаболически активные бактерии в кишечнике хлопкового листового червя ( Spodoptera littoralis ), многоядного насекомого-травоядного, потребляющего большое количество растений. материал за короткое время, высвобождая обильное количество глюкозы в пищеварительном тракте как наиболее важном источнике углерода и энергии как для хозяйских, так и для активных кишечных бактерий.Используя глюкозу 13 C в качестве трофического звена, Pyro-SIP показал, что относительно простая, но отличительная кишечная микробиота развивалась совместно с хозяином, и метаболическая активность, и состав менялись на протяжении личиночных стадий. Pantoea , Citrobacter и Clostridium были особенно активны в раннем возрасте, вероятно, основные функциональные популяции, связанные с улучшением питания. Enterococcus был единственным преобладающим родом в сообществе, и он был по существу стабильным и метаболически активным на протяжении жизни личинок.На основании того, что Enterococci образуют биопленкообразные слои на эпителии кишечника и что изолированные штаммы проявляют антимикробные свойства, Enterococcus может быть в состоянии создать эффект устойчивости к колонизации кишечника против потенциально вредных микробов извне. Это не только представляет собой первую углубленную инвентаризацию кишечной микробиоты модельного организма из в основном фитофагов Lepidoptera, но и это пилотное исследование показывает, что Pyro-SIP может быстро получить представление о метаболической активности кишечной микробиоты с высоким разрешением и высокой точностью. .
Enterococcus был единственным преобладающим родом в сообществе, и он был по существу стабильным и метаболически активным на протяжении жизни личинок.На основании того, что Enterococci образуют биопленкообразные слои на эпителии кишечника и что изолированные штаммы проявляют антимикробные свойства, Enterococcus может быть в состоянии создать эффект устойчивости к колонизации кишечника против потенциально вредных микробов извне. Это не только представляет собой первую углубленную инвентаризацию кишечной микробиоты модельного организма из в основном фитофагов Lepidoptera, но и это пилотное исследование показывает, что Pyro-SIP может быстро получить представление о метаболической активности кишечной микробиоты с высоким разрешением и высокой точностью. .
Образец цитирования: Shao Y, Arias-Cordero E, Guo H, Bartram S, Boland W. (2014) In vivo Pyro-SIP Оценка активной кишечной микробиоты хлопкового листового червя, Spodoptera littoralis . PLoS ONE 9 (1):
PLoS ONE 9 (1):
e85948.
https://doi.org/10.1371/journal.pone.0085948
Редактор: Малкольм Джеймс Хорсбург, Ливерпульский университет, Великобритания
Поступила: 3 октября 2013 г .; Одобрена: 4 декабря 2013 г .; Опубликован: 27 января 2014 г.
Авторские права: © 2014 Shao et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа поддержана Обществом Макса Планка. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Микробиота кишечника, в основном состоящая из бактерий, представляет собой сложную экосистему и образует тесные симбиотические ассоциации с хозяином. Эти внутренние микробные резиденты не только процветают в кишечнике, но и вносят свой вклад в метаболизм хозяина за счет высвобождения питательных веществ, детоксикации ксенобиотиков и иммунной регуляции, что значительно повышает приспособленность хозяина [1], [2]. С быстрым развитием технологий секвенирования нового поколения, все большее количество исследований, от людей до муравьев, продемонстрировало удивительное разнообразие и состав микробов в кишечнике [3], [4].Однако оценка метаболически активных компонентов микробиоты кишечника на сегодняшний день на удивление скудна, особенно с учетом физиологии хозяина. Ясно, что не все микроорганизмы способны колонизировать кишечник, даже несмотря на то, что это среда обитания, богатая питательными веществами, поскольку некоторые пищевые микробы лизируются, а некоторые временные микроорганизмы остаются в спящем состоянии во время прохождения через кишечник [2]. С другой стороны, активные популяции постоянно меняются в ответ на развитие хозяина и воздействие окружающей среды, и, следовательно, метаболические потенциалы, очерченные чистым метагеномным анализом, должны быть проверены in vivo [5].Чтобы получить более полную картину, важно выйти за рамки подхода, в основном основанного на секвенировании, к другим продвинутым инструментам для выявления активных фракций сообщества, которые непосредственно вносят вклад в текущую функцию микробиоты.
С другой стороны, активные популяции постоянно меняются в ответ на развитие хозяина и воздействие окружающей среды, и, следовательно, метаболические потенциалы, очерченные чистым метагеномным анализом, должны быть проверены in vivo [5].Чтобы получить более полную картину, важно выйти за рамки подхода, в основном основанного на секвенировании, к другим продвинутым инструментам для выявления активных фракций сообщества, которые непосредственно вносят вклад в текущую функцию микробиоты.
Зондирование стабильных изотопов (SIP) — многообещающий метод без культивирования, который часто используется в микробиологии окружающей среды для идентификации активных микроорганизмов, участвующих в различных биогеохимических процессах [6], [7]. Эта методология основана на ассимиляции источника углерода, меченного стабильным изотопом ( 13 C), растущими микробами и селективном извлечении обогащенных изотопами клеточных компонентов, таких как наиболее информативные нуклеиновые кислоты, которые могут предоставить конкретную таксономическую информацию. 13 C-обогащенная «тяжелая» ДНК или РНК может быть отделена от немеченых, нормальных «легких» ( 12 C) нуклеиновых кислот ультрацентрифугированием в градиенте плотности и впоследствии извлечена из процедуры градиентного фракционирования для дальнейшего молекулярного анализа [8]. Зондирование нуклеиновых кислот стабильными изотопами обеспечивает прямое доказательство метаболической активности бактерий и, как было показано, более чувствительно, чем подход, основанный на РНК [9]. Недавно Pilloni et al. представил комбинацию ДНК-SIP и пиросеквенирования, а именно Pyro-SIP, в интерпретации градиента SIP [10].Однако в этом исследовании проводилось пиросеквенирование только всей метагеномной ДНК без градиентного разделения и по-прежнему использовался метод снятия отпечатков пальцев на основе гель-электрофореза с низким разрешением (T-RFLP) и трудоемкая конструкция библиотеки клонов для идентификации активных бактерий. Учитывая все эти недостатки, здесь мы непосредственно проводим пиросеквенирование для изучения разделенных фракций градиента для получения большего разрешения, что относительно просто и эффективно объединяет исследование структуры микробиоты с измерением локальной метаболической активности.
13 C-обогащенная «тяжелая» ДНК или РНК может быть отделена от немеченых, нормальных «легких» ( 12 C) нуклеиновых кислот ультрацентрифугированием в градиенте плотности и впоследствии извлечена из процедуры градиентного фракционирования для дальнейшего молекулярного анализа [8]. Зондирование нуклеиновых кислот стабильными изотопами обеспечивает прямое доказательство метаболической активности бактерий и, как было показано, более чувствительно, чем подход, основанный на РНК [9]. Недавно Pilloni et al. представил комбинацию ДНК-SIP и пиросеквенирования, а именно Pyro-SIP, в интерпретации градиента SIP [10].Однако в этом исследовании проводилось пиросеквенирование только всей метагеномной ДНК без градиентного разделения и по-прежнему использовался метод снятия отпечатков пальцев на основе гель-электрофореза с низким разрешением (T-RFLP) и трудоемкая конструкция библиотеки клонов для идентификации активных бактерий. Учитывая все эти недостатки, здесь мы непосредственно проводим пиросеквенирование для изучения разделенных фракций градиента для получения большего разрешения, что относительно просто и эффективно объединяет исследование структуры микробиоты с измерением локальной метаболической активности.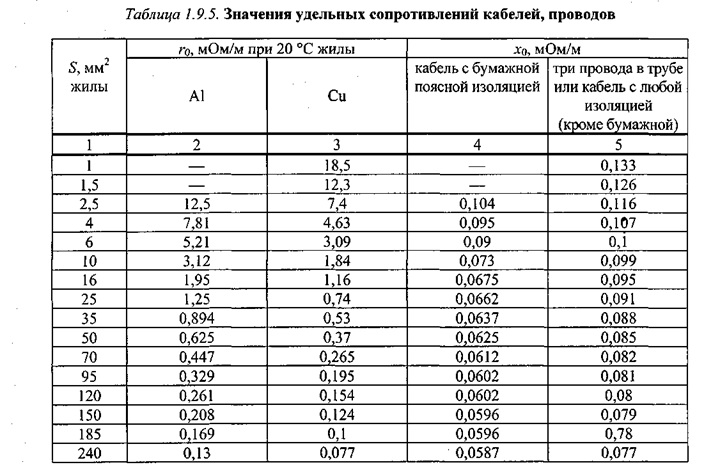
В качестве первой попытки мы успешно провели этот усовершенствованный Pyro-SIP, чтобы раскрыть метаболически активное бактериальное сообщество в кишечнике модельного организма насекомого, Spodoptera littoralis (Lepidoptera, Noctuidae), широко распространенного во всем мире многоядного вредителя, который дает значительный урожай. потери многих экономически важных культур. Опустошительная личиночная стадия S. littoralis (хлопковая листовая червь) потребляет большое количество растительного материала за короткое время, высвобождая в пищеварительном тракте обильные растительные сахариды, в основном глюкозу, которая является наиболее важным источником углерода и энергии как для хозяина, так и для организма. активные кишечные бактерии.Совсем недавно микробиология этого важного насекомого привлекла все большее внимание, в кишечнике личинок обнаружена сложная и изменчивая комменсальная микробиота [11], и предполагается, что кишечные бактерии участвуют в мультитрофических взаимодействиях между растением, травоядными и хищниками, что ранее было занижены [12] — [14].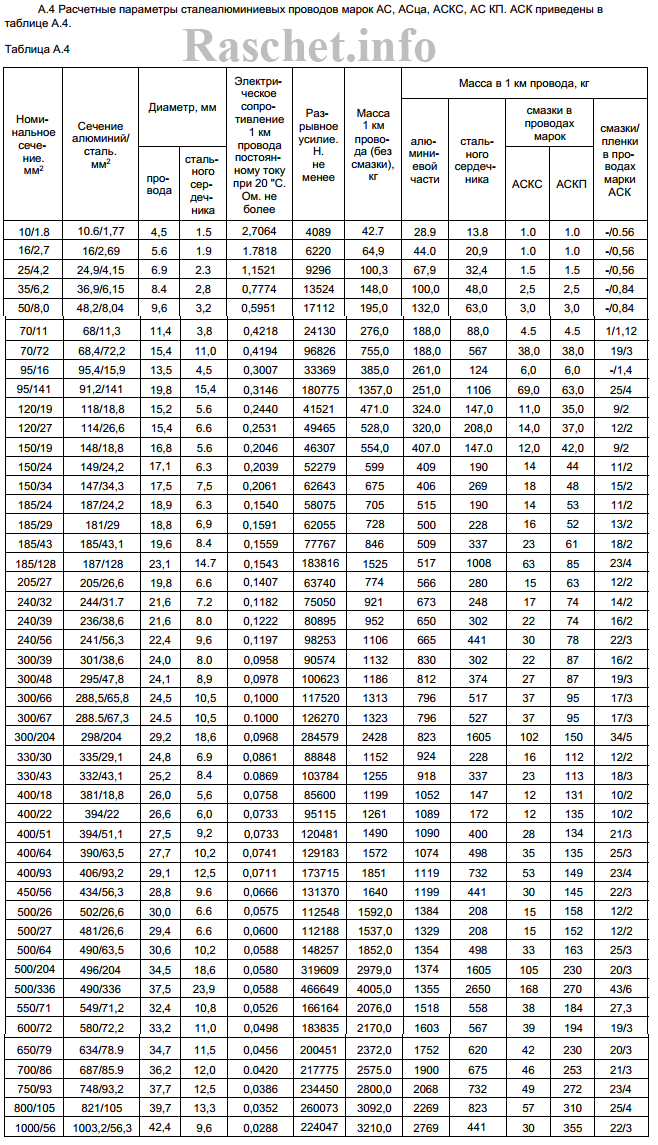
Используя глюкозу 13 C в качестве трофического звена, Pyro-SIP показал, что относительно простая, но отличительная кишечная микробиота развивается совместно с хозяином, причем как метаболическая активность, так и состав меняются на протяжении личиночных стадий.Три семейства, включая Enterococcaceae , Clostridiaceae и Enterobacteriaceae , особенно богаты и активны внутри, которые, вероятно, представляют собой основные функциональные популяции в кишечнике, связанные с улучшением питания и защитой от патогенов для улучшения приспособленности личинок. Это исследование также представляет собой первую углубленную инвентаризацию кишечной микробиоты модельного организма, состоящего в основном из фитофагов чешуекрылых. Знание кишечных бактерий такого крупного насекомого-травоядного может привести к новым целям в борьбе с вредителями.
Результаты
Идентификация и количественное определение сахаридов в кишечнике
Состав сахара был проанализирован в различных областях кишечника, а именно в передней, средней и задней кишках. Содержимое кишечника, собранное у личинок, получавших их естественный рацион (хлопок) или определенный искусственный рацион (без антибиотиков и консервантов), было проанализировано с помощью газовой хроматографии-масс-спектрометрии (ГХ-МС) после водной экстракции и дериватизации альдононитрилацетата, которая является быстрой и чувствительной. метод дериватизации нелетучих сахаридов, и каждый сахар образует один хроматографический пик после разделения с помощью ГХ [15], [16].
Содержимое кишечника, собранное у личинок, получавших их естественный рацион (хлопок) или определенный искусственный рацион (без антибиотиков и консервантов), было проанализировано с помощью газовой хроматографии-масс-спектрометрии (ГХ-МС) после водной экстракции и дериватизации альдононитрилацетата, которая является быстрой и чувствительной. метод дериватизации нелетучих сахаридов, и каждый сахар образует один хроматографический пик после разделения с помощью ГХ [15], [16].
Профиль ГХ-МС показал, что в кишечнике присутствовало несколько пентозных и гексозных сахаров, включая рибозу, арабинозу, маннозу, глюкозу и галактозу (рис. 1A и B). Однако глюкоза преобладала во всех частях кишечника и других мономерах при гораздо более низких концентрациях. Сходный образец состава сахарного мономера в данной личиночной секции был обнаружен в каждой повторности. После кислотного гидролиза количество и концентрация определяемых моносахаридов увеличивались, при этом преобладали глюкоза и арабиноза, отражая различные растворимые полисахариды в кишечнике (рис. S1 и S2).
S1 и S2).
Рис. 1. Состав сахара в кишечнике хлопковой листовой черви.
( A ) Сахар в кишечнике личинок, получавших искусственный корм из хлопка или ( B ), по данным ГХ-МС после получения дериватизации альдононитрилацетата. Личиночный пищеварительный тракт разделен на три области: переднюю, среднюю и заднюю кишки, как показано на диаграмме ( C ). ( D ) Количественное определение доминирующей глюкозы показывает значительное снижение среднего содержания в кишечнике.Однако личинки, питающиеся хлопком, содержат большее количество глюкозы во всех областях кишечника. * и a указывают на значительную разницу: P ( * 1 ) = 0,0020, P ( * 2 ) = 0,0366, P ( a ) = 0,0017. Планки погрешностей указывают на стандартные ошибки. 1, рибоза; 2, арабиноза; 3, манноза; 4, глюкоза; 5, Галактоза.
https://doi.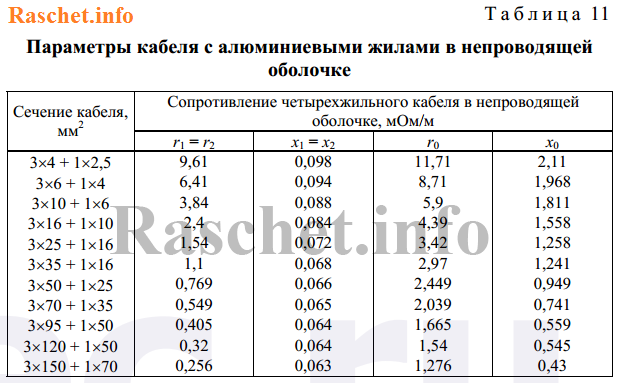 org/10.1371/journal.pone.0085948.g001
org/10.1371/journal.pone.0085948.g001
В качестве основного сахара глюкоза была определена количественно на основе специфического ферментативного анализа, который показал, что максимальные концентрации глюкозы наблюдались в передней кишке, а затем падали в передней части кишечника. к заднему концу кишки.У личинок, питающихся хлопком, средняя концентрация глюкозы в водной фазе передней кишки составляла приблизительно 17,7 мМ, что значительно выше, чем в задней кишке, которая составляла около 9,8 мМ (рис. 1D). Быстрое снижение уровня глюкозы от передней кишки к задней по короткому пищеварительному каналу указывало на то, что глюкоза значительно потреблялась во время прохождения через кишечник. Такая же тенденция наблюдалась и с личинками, получавшими искусственный корм. Однако средние значения концентрации глюкозы во всех отделах кишечника личинок, получавших искусственный корм, были ниже, чем у личинок, питавшихся хлопком (рис.1D).
Стратегия маркировки in vivo с глюкозой 13 C в качестве трофической связи
Как наиболее эффективный из доступных источников энергии и углерода, питательная глюкоза в кишечнике дает хороший шанс выполнить in vivo SIP путем внесения поправки на 13 C-глюкозы.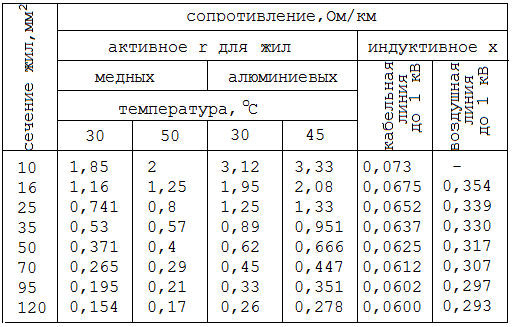 Начальные концентрации глюкозы в передней кишке личинок, питавшихся хлопком, достигли 20 мМ, но только около 10 мМ у личинок, питавшихся искусственным кормом (рис. 1D). Таким образом, в искусственную диету добавляли 10 мМ экзогенной глюкозы для имитации концентрации глюкозы in situ у личинок, питавшихся хлопком.Для отслеживания метаболически активных бактерий в качестве метки добавляли полностью 13 C-меченую глюкозу, тогда как такое же количество нативной глюкозы ( 12 C) использовали в контрольной группе. Мы протестировали процесс маркировки, скармливая личинок раннего возраста (от 1 st до 2 nd ) на искусственных диетах с добавлением глюкозы в течение 24 и 48 часов соответственно. Для выявления различий во время развития личинок тот же метод был использован при изучении личинок поздних возрастов (5 th ).
Начальные концентрации глюкозы в передней кишке личинок, питавшихся хлопком, достигли 20 мМ, но только около 10 мМ у личинок, питавшихся искусственным кормом (рис. 1D). Таким образом, в искусственную диету добавляли 10 мМ экзогенной глюкозы для имитации концентрации глюкозы in situ у личинок, питавшихся хлопком.Для отслеживания метаболически активных бактерий в качестве метки добавляли полностью 13 C-меченую глюкозу, тогда как такое же количество нативной глюкозы ( 12 C) использовали в контрольной группе. Мы протестировали процесс маркировки, скармливая личинок раннего возраста (от 1 st до 2 nd ) на искусственных диетах с добавлением глюкозы в течение 24 и 48 часов соответственно. Для выявления различий во время развития личинок тот же метод был использован при изучении личинок поздних возрастов (5 th ).
Личинок, взятых как из группы, получавшей 13 C, так и из немеченой контрольной группы, вскрывали в каждый момент времени, и общую ДНК экстрагировали из свежих тканей кишечника. Измерения с помощью масс-спектрометрии изотопного соотношения (IRMS) δ 13 C в экстрагированной ДНК показали значительное изотопное обогащение как через 24 часа (δ 13 C = 33,6 ± 5,3 ‰), так и через 48 часов (δ 13 C = 141,2 ± 33,1 ‰) мечения образцов по сравнению с естественным содержанием стабильных изотопов в контроле (δ 13 C = -30.7 ± 1,3 ‰) (рис. 2Б). Кормление в течение 48 часов усиливало процесс мечения с изотопным сдвигом, превышающим 100 ‰ (P = 0,0008), указывая на то, что многочисленные вновь разделенные бактериальные клетки уже заменили старые. Учитывая, что большое количество ДНК насекомых было совместно экстрагировано из ткани кишечника, реальная доля 13 C в чистой бактериальной ДНК будет еще выше, что подходит для центрифугирования в градиенте плотности для разделения меченых нуклеиновых кислот и идентификации активного сообщества. члены.
Измерения с помощью масс-спектрометрии изотопного соотношения (IRMS) δ 13 C в экстрагированной ДНК показали значительное изотопное обогащение как через 24 часа (δ 13 C = 33,6 ± 5,3 ‰), так и через 48 часов (δ 13 C = 141,2 ± 33,1 ‰) мечения образцов по сравнению с естественным содержанием стабильных изотопов в контроле (δ 13 C = -30.7 ± 1,3 ‰) (рис. 2Б). Кормление в течение 48 часов усиливало процесс мечения с изотопным сдвигом, превышающим 100 ‰ (P = 0,0008), указывая на то, что многочисленные вновь разделенные бактериальные клетки уже заменили старые. Учитывая, что большое количество ДНК насекомых было совместно экстрагировано из ткани кишечника, реальная доля 13 C в чистой бактериальной ДНК будет еще выше, что подходит для центрифугирования в градиенте плотности для разделения меченых нуклеиновых кислот и идентификации активного сообщества. члены.
Рисунок 2.Профилирование активных бактерий.
( A ) Для определения активных бактерий, 13 C-меченая ДНК отделяется ультрацентрифугированием в градиенте плотности и затем извлекается из репрезентативных фракций, причем более темный цвет указывает на меченую тяжелую ДНК. Все разделенные образцы ДНК (легкие, средние и тяжелые) были непосредственно подвергнуты количественному пиросеквенированию для выявления родословной и относительной численности видов. Для каждого таксона X метаболическая активность рассчитывается как разница в относительной численности тяжелой фракции меченого образца и контрольной.( B ) Изотопное соотношение (δ 13 C) образцов ДНК. ( C ) Распределение содержания ДНК в градиентных фракциях обработок глюкозой. Символы: O , ДНК, экстрагированная из контроля [ 12 C] -глюкозы; ▪ ДНК, экстрагированная после обработки [ 13 C] -глюкозой в течение 24 часов; ▴ , ДНК, экстрагированная после обработки [ 13 C] -глюкозой в течение 48 часов. Стрелки указывают на то, что значительная 13 C-меченая ДНК сдвинута в сторону сильных градиентов по сравнению с контролем.( D ) Градиентный фракционный анализ по плотности после 40 ч центрифугирования 1,725 г мл -1 исходного раствора CsCl.
Все разделенные образцы ДНК (легкие, средние и тяжелые) были непосредственно подвергнуты количественному пиросеквенированию для выявления родословной и относительной численности видов. Для каждого таксона X метаболическая активность рассчитывается как разница в относительной численности тяжелой фракции меченого образца и контрольной.( B ) Изотопное соотношение (δ 13 C) образцов ДНК. ( C ) Распределение содержания ДНК в градиентных фракциях обработок глюкозой. Символы: O , ДНК, экстрагированная из контроля [ 12 C] -глюкозы; ▪ ДНК, экстрагированная после обработки [ 13 C] -глюкозой в течение 24 часов; ▴ , ДНК, экстрагированная после обработки [ 13 C] -глюкозой в течение 48 часов. Стрелки указывают на то, что значительная 13 C-меченая ДНК сдвинута в сторону сильных градиентов по сравнению с контролем.( D ) Градиентный фракционный анализ по плотности после 40 ч центрифугирования 1,725 г мл -1 исходного раствора CsCl. Планки погрешностей указывают на стандартные ошибки.
Планки погрешностей указывают на стандартные ошибки.
https://doi.org/10.1371/journal.pone.0085948.g002
Ультрацентрифугирование в градиенте плотности и извлечение разделенной ДНК
И меченые, и контрольные образцы были взяты из одной партии изопикнического ультрацентрифугирования, чтобы минимизировать возможные вариации во время этого процесса.После 40 ч ультрацентрифугирования сформированные градиенты были фракционированы на 12 равных аликвот, и количество ДНК, присутствующей в каждой отдельной фракции, было определено количественно, что позволило нам сравнить распределение ДНК-матрицы в сформированных градиентах (рис. 2А). Плотность всех фракций из контрольных и меченых образцов проверяли взвешиванием, охватывая средний градиент от 1,688 г / мл -1 до 1,761 г / мл -1 , что находилось в ожидаемом диапазоне согласно предыдущим отчетам [8] .Определение плотности показало линейный тренд снизу вверх, что указывает на формирование правильного градиента (рис. 2D).
2D).
После 24 часов кормления меченую ДНК из метаболически активных бактерий можно было измерить при обработке 13 C, которая имела новый пик обильной ДНК при плавучей плотности (BD) 1,730 г / мл -1 (фракция 5). Через 48 ч пик сместился в еще более тяжелую фракцию при BD 1,735 мкг / мл -1 (фракция 4), что указывает на большее включение 13 C в ДНК, что подтверждает данные IRMS.Напротив, большая часть ДНК из немеченого контроля все еще распределялась по легким фракциям (BD составляла <1,718 мкг / мл -1 ), и пик не сдвигался в сторону высокого BD (фиг. 2C). Примечательно, что низкий фон неспецифических нуклеиновых кислот был обнаружен во всех градиентных фракциях, что является обычным в эксперименте SIP в окружающей среде [17]. Поскольку образец, полученный в течение 48 часов, генерировал значительно более высокий уровень обогащения, а больший сдвиг BD обеспечивал эффективное отделение меченой изотопом ДНК от немеченой ДНК, образцы этой обработки были отобраны для последующей молекулярной характеристики микробного сообщества.
Ген 16S рРНК амплифицировали с общим бактериальным праймером из каждой градиентной фракции. Амплификация контрольных градиентов дала явные продукты ПЦР из фракций с 8 по 12, что соответствует мазку ДНК, сформированному после ультрацентрифугирования, и некоторые слабые полосы в тяжелых фракциях, которые ожидались от неспецифической фоновой ДНК. Напротив, меченый образец показал повышенную интенсивность полос в соответствующих тяжелых фракциях (3–6), что было вызвано повышенным количеством 13 матрицы С-ДНК.Затем ампликоны 16S рДНК, обнаруженные в градиентных фракциях, анализировали денатурирующим градиентным гель-электрофорезом (DGGE), чтобы «отпечатать пальцем» сообщество. Изменения стали заметны, когда профиль DGGE, полученный в результате обработки 13 C, сравнивался с профилем немеченого контроля. Несколько сильных полос были четко обнаружены в тяжелых фракциях (3–6) меченого образца, тогда как картина практически не изменилась во всех градиентах контроля (рис. S3). Эти уникальные паттерны, связанные с тяжелыми фракциями из образца с измененной стабильной изотопной поправкой, но не с нативным контролем с измененным субстратом, предоставили убедительные доказательства связи определенных организмов с метаболической активностью in situ и кишечника.Взятые вместе, данные подтвердили успешное включение 13 C в ДНК активных кишечных бактерий.
S3). Эти уникальные паттерны, связанные с тяжелыми фракциями из образца с измененной стабильной изотопной поправкой, но не с нативным контролем с измененным субстратом, предоставили убедительные доказательства связи определенных организмов с метаболической активностью in situ и кишечника.Взятые вместе, данные подтвердили успешное включение 13 C в ДНК активных кишечных бактерий.
На основе анализа отпечатков пальцев мы объединили фракции с BD в диапазоне от 1,730 до 1,735 г / мл -1 в скомпилированную «тяжелую» фракцию. BD составляли около 1,718 мкг / мл -1 в «средней» фракции и от 1,688 до 1,705 мкг / мл -1 в «легкой» фракции для последующего пиросеквенирования.
Общая микробная структура в кишечной флоре и филогенетический анализ
Количественное пиросеквенирование проводили непосредственно на представительных градиентах SIP для выявления родословной и относительной численности видов.ПЦР-амплификация с использованием праймеров, специфичных для архей или грибов, не позволила амплифицировать какие-либо последовательности 16S рРНК архей или ITS-последовательности грибов. Попытки извлечь грибы из кишечника с помощью трех обычных чашек с агаром для выращивания грибов также были безуспешными (рис. S4), основанные на культивировании.
Попытки извлечь грибы из кишечника с помощью трех обычных чашек с агаром для выращивания грибов также были безуспешными (рис. S4), основанные на культивировании.
Бактерии-специфический праймер для пиросеквенирования амплифицировал область V1 – V3 гена 16S рРНК для таксономической классификации. После шумоподавления и удаления химерных последовательностей было произведено 120 045 считываний высокого качества со средней длиной 404 нуклеотида.Была низкая частота последовательностей, не классифицированных на уровне филума (максимум 3,82%). Для всех образцов кривая разрежения имела тенденцию к насыщению при одинаковом количестве последовательностей на уровне видов, что указывает на то, что выборка была всеобъемлющей (рис. 3A и 4A). Для точной оценки разнообразия отдельных выборок использовались различные методы (таблица 1). Микробиота кишечника хлопковой листовой черви была таксономически ограничена и содержала 22–42 операционных таксономических единицы (OTU), обнаруженных при 97% сходстве последовательностей, при этом Proteobacteria и Firmicutes были содоминантными типами. Индекс разнообразия Шеннона 1,46–2,48 являлся нижней границей разнообразия почв (2,4–3,6) [18].
Индекс разнообразия Шеннона 1,46–2,48 являлся нижней границей разнообразия почв (2,4–3,6) [18].
Рис. 3. Бактериальное разнообразие и относительное количество в кишечной микробиоте личинок раннего возраста.
( A ) Кривые разрежения последовательностей 16S рДНК получали из репрезентативных фракций SIP контроля ([ 12 C]) и обработки меткой ([ 13 C]). ( B ) Относительное количество бактериальных таксонов в различных фракциях SIP, представленное на графике относительных площадей, выявленное пиросеквенированием.Сокращения: [ 12 C] Легкие, легкие фракции (фракции 9–11, фиг. 2A) поправки на нативную глюкозу; [ 12 C] Средняя, средняя фракция (фракция 7) этого; [ 12 C] Тяжелые тяжелые фракции (фракции 4–5) из них; [ 13 C] Легкие, легкие фракции 13 C-глюкозная поправка; [ 13 C] Средняя, средняя часть этого; [ 13 C] Тяжелые, тяжелые его части.
https://doi.org/10.1371/journal.pone.0085948.g003
Рис. 4. Бактериальное разнообразие и относительная численность кишечной микробиоты личинок позднего возраста.
( A ) Кривые разрежения последовательностей 16S рДНК были получены из репрезентативных фракций SIP личинок раннего и позднего возраста. Сокращения: E, репрезентативные фракции от личинок раннего возраста, питавшихся 13 C-глюкозой; L, фракции от личинок позднего возраста, питавшихся 13 С-глюкозой. ( B ) Относительное количество бактериальных таксонов в различных фракциях SIP, представленное на графике относительных площадей, выявленное пиросеквенированием.Сокращения такие же, как на рис. 3.
https://doi.org/10.1371/journal.pone.0085948.g004
«Легкая» фракция, собранная из немеченых контролей, где все еще распределяется большая часть метагеномной ДНК, служила для профилирования большей части бактериального сообщества. Из-за того, что меньшее количество матриц ДНК из доминантных групп перемещается в тяжелую фракцию контроля, некоторые менее распространенные бактерии, такие как Comamonas и Stella , имели больше шансов на амплификацию и показали повышенную долю в профиле [ 12 C] -Тяжелая фракция (фиг.3Б). Таким образом, всесторонний анализ всех фракций немеченого контроля SIP предоставил подробную информацию об общем разнообразии сообщества, которое во многом схоже с таковым у личинок, получавших диету из белой фасоли, без ультрацентрифугирования ДНК (рис. S6). Явное снижение численности доминирующих групп, таких как Pantoea и Citrobacter , в средней и тяжелой фракциях, вызванное меньшим количеством целевых последовательностей, кроме того, отражает некоторые количественные свойства пиросеквенирования [19].Всего одиннадцать типов были идентифицированы классификатором RDP из набора данных, но только пять показали относительную численность, превышающую 0,1%, по крайней мере, в одном из проанализированных образцов (Таблица 2).
Из-за того, что меньшее количество матриц ДНК из доминантных групп перемещается в тяжелую фракцию контроля, некоторые менее распространенные бактерии, такие как Comamonas и Stella , имели больше шансов на амплификацию и показали повышенную долю в профиле [ 12 C] -Тяжелая фракция (фиг.3Б). Таким образом, всесторонний анализ всех фракций немеченого контроля SIP предоставил подробную информацию об общем разнообразии сообщества, которое во многом схоже с таковым у личинок, получавших диету из белой фасоли, без ультрацентрифугирования ДНК (рис. S6). Явное снижение численности доминирующих групп, таких как Pantoea и Citrobacter , в средней и тяжелой фракциях, вызванное меньшим количеством целевых последовательностей, кроме того, отражает некоторые количественные свойства пиросеквенирования [19].Всего одиннадцать типов были идентифицированы классификатором RDP из набора данных, но только пять показали относительную численность, превышающую 0,1%, по крайней мере, в одном из проанализированных образцов (Таблица 2). Фирмикуты, по-видимому, доминировали на обеих стадиях развития личинок, что составляло 59,2% от числа последовательностей личинок раннего возраста и 97,2% последовательностей личинок позднего возраста.
Фирмикуты, по-видимому, доминировали на обеих стадиях развития личинок, что составляло 59,2% от числа последовательностей личинок раннего возраста и 97,2% последовательностей личинок позднего возраста.
Протеобактерии были еще одним основным филумом личинок раннего возраста, составляя 38,9% от общего числа последовательностей. Бактерии этого типа особенно широко распространены в микробиотах травоядных и часто связаны с симбионтами насекомых.Наиболее доминирующие ОТЕ соответствовали Pantoea citrea и Citrobacter farmeri , что составило 17,2% и 16,4% прочтений, соответственно, и принадлежало к семейству Enterobacteriaceae класса Gammaproteobacteria ([ 12 C] свет, рис. . 3B). Помимо Enterobacteriaceae , были обнаружены другие гаммапротеобактерии из семейства Xanthomonadaceae , Pseudomonadaceae и Moraxellaceaee , включая популяции, связанные с Stenotrophomonas , Pseudomonas и Acinetobacter. 3б и 5б). Также были идентифицированы другие классы Proteobacteria, такие как Paracoccus из Alphaproteobacteria и Geobacter из Deltaproteobacteria. Организмы из филума актинобактерий, включая роды Micrococcus , Solirubrobacter и Propionibacterium , а также новую группу, GP2, из филума Acidobacteria составили небольшую долю ридов. Однако эти редкие филотипы способствовали богатству сообщества.
3б и 5б). Также были идентифицированы другие классы Proteobacteria, такие как Paracoccus из Alphaproteobacteria и Geobacter из Deltaproteobacteria. Организмы из филума актинобактерий, включая роды Micrococcus , Solirubrobacter и Propionibacterium , а также новую группу, GP2, из филума Acidobacteria составили небольшую долю ридов. Однако эти редкие филотипы способствовали богатству сообщества.
Рис. 5. Филогенетический анализ (a) Firmicutes и (b) Proteobacteria, идентифицированных из микробиоты кишечника хлопкового листового червя.
( A ) Дерево максимального правдоподобия было получено из данных частичной последовательности 16S рДНК для членов Firmicutes. ( B ) Дерево присоединения соседей было получено из данных о частичных последовательностях 16S рДНК для членов Proteobacteria. Репрезентативные пиросеквенции из этой работы и почти полноразмерные последовательности 16S рДНК, полученные из предыдущих исследований на основе библиотеки клонов, обозначены черными кружками (•) и синими кружками, соответственно. Помеченные таксоны отмечены треугольниками (▴). Контрольные последовательности загружаются из GenBank (инвентарные номера указаны в скобках). Methanosarcina barkeri (AF028692) используется в качестве внешней группы. Кластеры на уровне семьи обозначены разными цветами. Значения начальной загрузки (в процентах) основаны на 1000 репликаций. Столбик соответствует 2% расхождению последовательности. Правый раздел показывает процент репрезентативных бактериальных последовательностей 16S рРНК в общем наборе данных каждого образца. Сокращения: + 12 C, добавка нативной глюкозы; + 13 C, 13 C-глюкозная поправка; L, легкие фракции; H, тяжелые фракции.
Помеченные таксоны отмечены треугольниками (▴). Контрольные последовательности загружаются из GenBank (инвентарные номера указаны в скобках). Methanosarcina barkeri (AF028692) используется в качестве внешней группы. Кластеры на уровне семьи обозначены разными цветами. Значения начальной загрузки (в процентах) основаны на 1000 репликаций. Столбик соответствует 2% расхождению последовательности. Правый раздел показывает процент репрезентативных бактериальных последовательностей 16S рРНК в общем наборе данных каждого образца. Сокращения: + 12 C, добавка нативной глюкозы; + 13 C, 13 C-глюкозная поправка; L, легкие фракции; H, тяжелые фракции.
https://doi.org/10.1371/journal.pone.0085948.g005
Фирмикуты были представлены семейством Enterococcaceae , составляющим> 50% всех последовательностей. Главный OTU был тесно связан с Enterococcus mundtii , как определено с помощью Mega BLAST с репрезентативной последовательностью.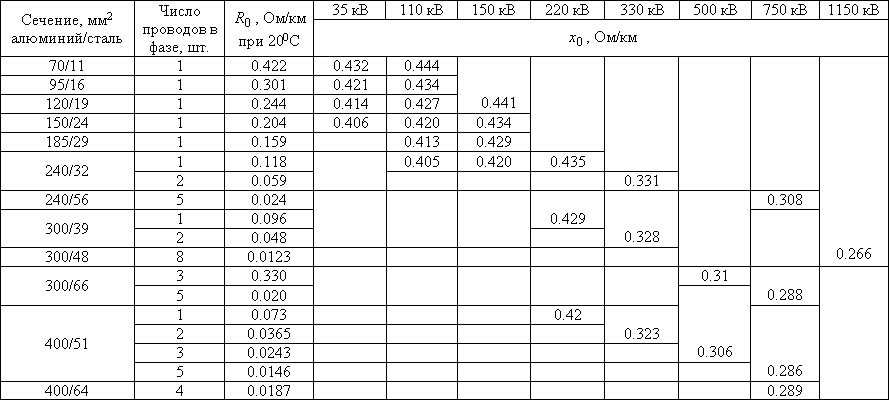 Были обнаружены и другие виды Enterococcus с низкой идентификационной оценкой, такие как E. casseliflavus , но они составили лишь небольшую часть.Небольшое количество Clostridia было также получено от личинок раннего возраста.
Были обнаружены и другие виды Enterococcus с низкой идентификационной оценкой, такие как E. casseliflavus , но они составили лишь небольшую часть.Небольшое количество Clostridia было также получено от личинок раннего возраста.
Микробиота кишечника претерпела резкие изменения в позднем возрасте, в основном характеризуясь уменьшением видового богатства и разнообразия (Таблица 1). Кривая разрежения была значительно ниже, чем у личинок раннего возраста (рис. 4А). Firmicutes полностью доминировали во всем сообществе, составляя> 97% всех прочтений; одновременно уменьшались последовательности из других типов ([ 12 C] -свет, рис.4Б). Чтения пиросеквенирования, принадлежащие к семейству Clostridiaceae , сильно увеличились: 21% от общего числа последовательностей были связаны с видами Clostridium . Enterococcus по-прежнему преобладали, составляя 75% считываний. Были обнаружены и другие бактерии Firmicutes, такие как представители родов Lactobacillus , Lactococcus , Anoxybacillus и Staphylococcus (рис. 5A).
5A).
Однако в контексте Proteobacteria личинки позднего возраста в основном снижали долю Gammaproteobacteria. Pantoea был обнаружен в небольшой пропорции, всего лишь с 1,8% от общего числа последовательностей ([ 12 C] -свет, рис. 4B). Citrobacter быстро уменьшились вместе. Помимо обычного Paracoccus , было обнаружено больше Alphaproteobacteria, например представители родов Sphingomonas , Caulobacter и Rhizobium . Но при этом были удалены Stella и Methylobacterium . Бетапротеобактерии исчезли из личинок позднего возраста, за исключением Vogesella .Другие филогенетические группы, относящиеся к родам Prevotella (Bacteroidetes), Micrococcus и Propionibacterium (Actinobacteria), появились в небольшом количестве последовательностей.
Филогенетический анализ показал, что некоторые риды пиросеквенирования сгруппированы вместе с почти полной длиной последовательностей 16S, извлеченных из других исследований на основе библиотек клонов, что указывает на общие таксоны, присутствующие в S. littoralis (рис. 5). Примечательно, что состав микробиоты кишечника личинки, получавшей эту искусственную диету с добавлением физиологической дозы сахара, был аналогичен составу микробиоты, получавшей ее естественную растительную диету [11].Многие малочисленные таксоны были обнаружены в результате нашего крупномасштабного пиросеквенирования, которое более точно представляло общее микробное сообщество, а также обеспечивало достаточное таксономическое разрешение.
littoralis (рис. 5). Примечательно, что состав микробиоты кишечника личинки, получавшей эту искусственную диету с добавлением физиологической дозы сахара, был аналогичен составу микробиоты, получавшей ее естественную растительную диету [11].Многие малочисленные таксоны были обнаружены в результате нашего крупномасштабного пиросеквенирования, которое более точно представляло общее микробное сообщество, а также обеспечивало достаточное таксономическое разрешение.
Характеристика метаболически активных бактерий в сообществе
«Тяжелые» фракции, собранные из меченого образца, содержащего значительные количества 13 С-обогащенной ДНК, и фракции, собранные из немеченого контроля, служили для профилирования активных популяций в сообществе.На основании предыдущих исследований метаболическая активность бактерий оценивалась путем расчета разницы между относительной численностью отдельных таксонов в тяжелой фракции меченого образца и в контроле (рис. 2А) [20], [21]. Более того, вовлечение надлежащего контроля в SIP-анализ вычитало влияние фонового загрязнения 12 C-ДНК в тяжелой фракции, что гарантировало, что появление или исчезновение бактериального разнообразия и численности не было артефактом самого метода.Кривая разрежения образца «тяжелой» фракции была ниже, чем у «легкой» фракции, потому что группа активных бактерий была подмножеством всего сообщества (рис. 3A).
Более того, вовлечение надлежащего контроля в SIP-анализ вычитало влияние фонового загрязнения 12 C-ДНК в тяжелой фракции, что гарантировало, что появление или исчезновение бактериального разнообразия и численности не было артефактом самого метода.Кривая разрежения образца «тяжелой» фракции была ниже, чем у «легкой» фракции, потому что группа активных бактерий была подмножеством всего сообщества (рис. 3A).
В раннем возрасте профиль пиросеквенирования тяжелой фракции из меченого образца ([ 13 C] -Heavy, рис. 3B) показал значительное увеличение численности определенных видов, включая Pantoea , Citrobacter и Clostridium. с помощью анализа METASTATS (P <0,05), тогда как незначительные части этих последовательностей были обнаружены в тяжелой фракции контроля ([ 12 C] -Heavy, рис.3Б). Поэтому ДНК этих бактерий была значительно помечена, и они считались метаболически активными. Наибольшая активность была оценена для Pantoea и Citrobacter с фактором стимуляции выше 10% (рис. 6). Разнообразный набор других протеобактерий из родов Acinetobacter , Stenotrophomonas , Delftia , Achromobacter и Vogesella также продемонстрировал некоторую активность в соответствии с их слегка повышенной интенсивностью в тяжелой фракции. Clostridium была другой группой, которая показала высокую метаболическую активность, хотя на этой стадии ее было меньше. Близкородственные филотипы этих активных видов обычно идентифицируются по кишечнику травоядных животных и являются хорошо известными деструкторами растительной биомассы. Enterococcus в изобилии присутствовал во всех фракциях SIP независимо от состояния, но количество последовательностей достигло пика в средней фракции меченого образца по сравнению с таковым в контроле, что указывает на то, что он был метаболически менее активен, а ДНК из которого была включена с 13 ° С и не смог полностью перейти в тяжелую фракцию.Представители таких родов, как Paracoccus , Solirubrobacter и Propionibacterium , не были ни обогащены, ни обнаружены во фракции 13 C-ДНК, и, таким образом, считались метаболически неактивными бактериями.
6). Разнообразный набор других протеобактерий из родов Acinetobacter , Stenotrophomonas , Delftia , Achromobacter и Vogesella также продемонстрировал некоторую активность в соответствии с их слегка повышенной интенсивностью в тяжелой фракции. Clostridium была другой группой, которая показала высокую метаболическую активность, хотя на этой стадии ее было меньше. Близкородственные филотипы этих активных видов обычно идентифицируются по кишечнику травоядных животных и являются хорошо известными деструкторами растительной биомассы. Enterococcus в изобилии присутствовал во всех фракциях SIP независимо от состояния, но количество последовательностей достигло пика в средней фракции меченого образца по сравнению с таковым в контроле, что указывает на то, что он был метаболически менее активен, а ДНК из которого была включена с 13 ° С и не смог полностью перейти в тяжелую фракцию.Представители таких родов, как Paracoccus , Solirubrobacter и Propionibacterium , не были ни обогащены, ни обнаружены во фракции 13 C-ДНК, и, таким образом, считались метаболически неактивными бактериями.
Рисунок 6. Частота последовательностей 16S рРНК в микробиоте, полученных из контроля нативной глюкозы (относительное количество бактерий) и обработки [ 13 C] -глюкозы (метаболическая активность бактерий), представленная в виде тепловой карты.
Левая панель отображает динамические изменения таксонов у личинок раннего возраста, правая панель — позднего возраста.Теплые цвета указывают на более высокую, а холодные цвета — на более низкую численность, рассчитанную по формуле на рис. 2А (также см. Рис. 5 для процентного содержания таксонов).
https://doi.org/10.1371/journal.pone.0085948.g006
В позднем возрасте Enterococcus были более многочисленны в тяжелой фракции меченого образца, чем в контрольной, будучи наиболее активным членом внутри сообщества. Флуоресценция in situ при гибридизации (FISH) выявила большое количество Enterococci , плотно прилегающих к слизистому слою эпителия кишечника, где они образовывали толстую биопленочную структуру (рис.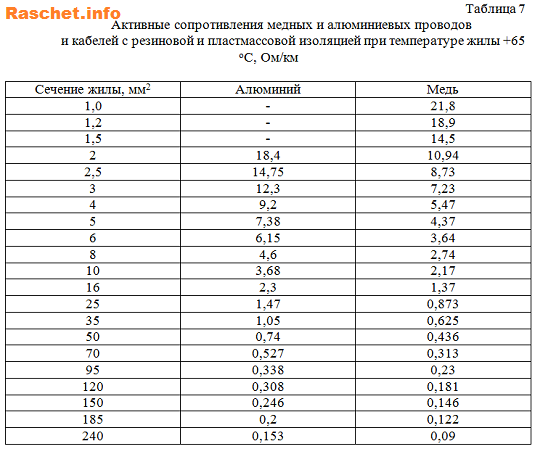 S5a и b). Интересно, что изоляты Enterococcus могут продуцировать новые противомикробные соединения против других бактерий (рис. S5c и d). Clostridium sp. оставалась на высоком уровне популяции, что было следствием высокой активности в раннем возрасте; однако на этом этапе метаболическая активность была незначительной или отсутствовала. Pantoea и другая Proteobacteria, Vogesella , были активно персистентными на обеих стадиях развития. В отличие от Citrobacter не был обнаружен.Но особенно активен был род Legionella . Bacteroidetes, включая Prevotella и Hymenobacter , проявили некоторую метаболическую активность. Было обнаружено, что другие популяции были активными, в том числе Tetrasphaera , Microbacterium и Corynebacterium , связанные с типом Actinobacteria, и Anoxybacillus из Firmicutes.
S5a и b). Интересно, что изоляты Enterococcus могут продуцировать новые противомикробные соединения против других бактерий (рис. S5c и d). Clostridium sp. оставалась на высоком уровне популяции, что было следствием высокой активности в раннем возрасте; однако на этом этапе метаболическая активность была незначительной или отсутствовала. Pantoea и другая Proteobacteria, Vogesella , были активно персистентными на обеих стадиях развития. В отличие от Citrobacter не был обнаружен.Но особенно активен был род Legionella . Bacteroidetes, включая Prevotella и Hymenobacter , проявили некоторую метаболическую активность. Было обнаружено, что другие популяции были активными, в том числе Tetrasphaera , Microbacterium и Corynebacterium , связанные с типом Actinobacteria, и Anoxybacillus из Firmicutes.
В совокупности in situ SIP обозначают различные паттерны метаболической активности, обнаруженные в микробиоте кишечника во время личиночного развития. Консорциум из Enterobacteriaceae (особенно Pantoea , Citrobacter ) и Clostridium , по-видимому, был более активен у личинок раннего возраста, тогда как Enterococcus стал сильно активным у полностью выросших личинок позднего возраста (рис. 6). Эти изменения могут напрямую связываться с их функциональными ролями внутри кишечника.
Консорциум из Enterobacteriaceae (особенно Pantoea , Citrobacter ) и Clostridium , по-видимому, был более активен у личинок раннего возраста, тогда как Enterococcus стал сильно активным у полностью выросших личинок позднего возраста (рис. 6). Эти изменения могут напрямую связываться с их функциональными ролями внутри кишечника.
Обсуждение
Хотя SIP часто используется в микробиологии окружающей среды, очень немногие исследователи применили этот ценный метод к изучению микробиоты кишечника.Эти новаторские исследования почти исключительно использовали экспериментальные системы in vitro и для имитации среды кишечника, которая, очевидно, не могла полностью воспроизвести истинные физиологические условия в неповрежденном кишечнике, особенно факторы хозяина, формирующие микробное сообщество, и, следовательно, могут быть ограниченными или предвзятыми [5 ], [20].
Напротив, мы непосредственно изучали микробиоту кишечника хлопкового листового червя in vivo путем сочетания пиросеквенирования и SIP подходов. Хлопковые листовые черви обладают простым трубчатым пищеварительным трактом, который составляет большую часть всего тела и не имеет каких-либо специализированных субструктур (рис.1С). Несмотря на его простоту, было обнаружено, что большое количество бактерий, превышающее 10 7 мл -1 [22], занимает кишечник, и там выделяется высокая концентрация питательной глюкозы, что делает кишечник идеальной средой для разнообразных микробных микроорганизмов. деятельности, включая брожение. Эти свойства делают этот организм идеальной естественной моделью для изучения микробного симбиоза пищеварительного тракта. В качестве эффективного и повсеместного источника энергии и углерода глюкоза обеспечивает полезную трофическую связь для идентификации метаболически активных кишечных бактерий путем отслеживания их ассимиляции измененной 13 C-глюкозы в искусственной диете.Кишечник содержит другие немеченые источники углерода, такие как аминокислоты, которые могут быть использованы микробами.
Хлопковые листовые черви обладают простым трубчатым пищеварительным трактом, который составляет большую часть всего тела и не имеет каких-либо специализированных субструктур (рис.1С). Несмотря на его простоту, было обнаружено, что большое количество бактерий, превышающее 10 7 мл -1 [22], занимает кишечник, и там выделяется высокая концентрация питательной глюкозы, что делает кишечник идеальной средой для разнообразных микробных микроорганизмов. деятельности, включая брожение. Эти свойства делают этот организм идеальной естественной моделью для изучения микробного симбиоза пищеварительного тракта. В качестве эффективного и повсеместного источника энергии и углерода глюкоза обеспечивает полезную трофическую связь для идентификации метаболически активных кишечных бактерий путем отслеживания их ассимиляции измененной 13 C-глюкозы в искусственной диете.Кишечник содержит другие немеченые источники углерода, такие как аминокислоты, которые могут быть использованы микробами. Однако эти метаболически активные бактерии должны одновременно потреблять и доминирующую общую глюкозу, и поэтому их метят. Почти in situ концентрация глюкозы и относительно короткое время маркировки гарантируют, что исходное кишечное сообщество не будет подвержено влиянию подхода SIP, как подробно объясняется в другом месте [20]. Перекрестное кормление обычно является основным ограничением в интерпретации экспериментов SIP, что не является проблемой в настоящем исследовании, поскольку мы стремились захватить все активное сообщество, и активные бактерии, скорее всего, вовлечены в трофическую сеть in situ [23].ДНК, полученная при обработке 13 C-глюкозой, обогащалась 13 C на основе заметного сдвига ДНК в сторону более высоких значений BD; Напротив, никакого сдвига не наблюдалось для контроля с измененной нативной глюкозой. Существенное мечение ДНК было также подтверждено дополнительными методами, включая масс-спектрометрическое измерение изотопного отношения и анализ отпечатков пальцев.
Однако эти метаболически активные бактерии должны одновременно потреблять и доминирующую общую глюкозу, и поэтому их метят. Почти in situ концентрация глюкозы и относительно короткое время маркировки гарантируют, что исходное кишечное сообщество не будет подвержено влиянию подхода SIP, как подробно объясняется в другом месте [20]. Перекрестное кормление обычно является основным ограничением в интерпретации экспериментов SIP, что не является проблемой в настоящем исследовании, поскольку мы стремились захватить все активное сообщество, и активные бактерии, скорее всего, вовлечены в трофическую сеть in situ [23].ДНК, полученная при обработке 13 C-глюкозой, обогащалась 13 C на основе заметного сдвига ДНК в сторону более высоких значений BD; Напротив, никакого сдвига не наблюдалось для контроля с измененной нативной глюкозой. Существенное мечение ДНК было также подтверждено дополнительными методами, включая масс-спектрометрическое измерение изотопного отношения и анализ отпечатков пальцев.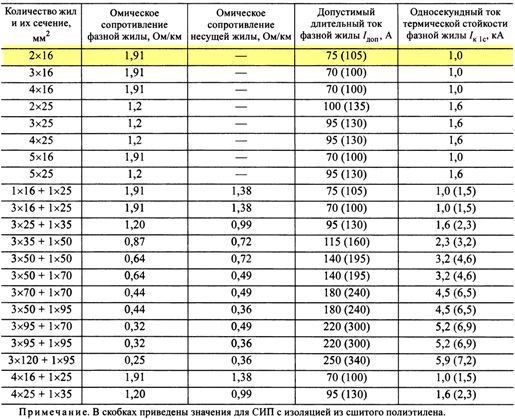 In vivo SIP предоставляет ключевую информацию о том, какие группы в настоящее время участвуют в метаболизме кишечника, и эти активные бактериальные ассоциаты потенциально способствуют приспособленности хозяина.Большая глубина выборки пиросеквенирования гарантирует эффективное обследование разнообразия, а анализ разреженности показал, что наш набор данных обеспечивает достаточную степень охвата для всех образцов.
In vivo SIP предоставляет ключевую информацию о том, какие группы в настоящее время участвуют в метаболизме кишечника, и эти активные бактериальные ассоциаты потенциально способствуют приспособленности хозяина.Большая глубина выборки пиросеквенирования гарантирует эффективное обследование разнообразия, а анализ разреженности показал, что наш набор данных обеспечивает достаточную степень охвата для всех образцов.
Всесторонний анализ восстановленной ДНК выявил относительно простую, но отличительную кишечную микробиоту, которая развивается совместно с хозяином; как состав, так и метаболическая активность устойчиво изменяются на протяжении личиночных стадий. В целом в считываниях пиросеквенирования преобладали таксоны из Proteobacteria и Firmicutes.Подобная картина недавно была описана у личинок плодовых мух [24]. Примечательно, что бактерии этих типов широко распространены в кишечнике муравьев-черепах, пчел, мотыльков и бабочек, что позволяет предположить, что они могут представлять основной тип микробиоты кишечника травоядных насекомых [25] — [27].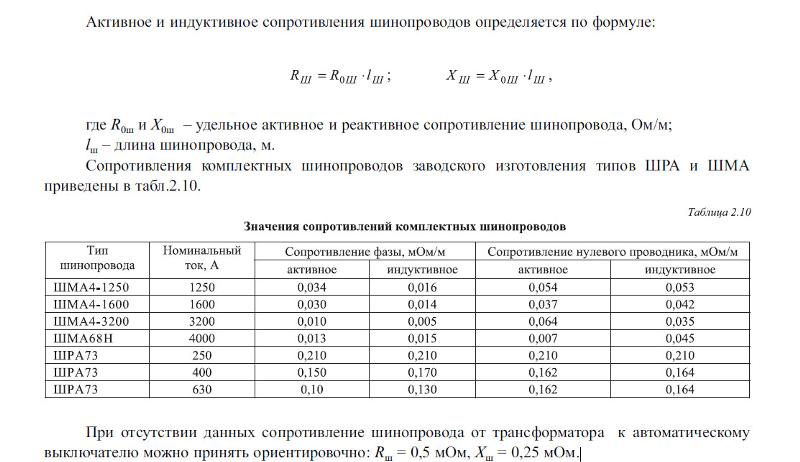 Простота этого сообщества, состоящего из 22–42 OTU, особенно очевидна по сравнению с микробиотой кишечника насекомых из таких отрядов, как термиты Isoptera или позвоночные, которые часто являются носителями сотен филотипов [28], [29].Сильная щелочность в кишечнике, считающаяся важной детерминантой структуры сообщества личинок жуков, также может иметь место у Spodoptera , у которого pH средней кишки> 10 [22], [30]. Другие ключевые факторы, включая производительность фаст-фуда и функцию иммунной системы, также могут составлять таксономически ограниченную кишечную флору [31]. Однако три семейства, включая Enterococcaceae , Clostridiaceae и Enterobacteriaceae , особенно богаты и активны и, вероятно, являются основными функциональными популяциями, живущими внутри кишечника.Кроме того, обнаруживаются явные изменения в развитии (ранний возраст по сравнению с поздним возрастом). Мало того, что они постоянно встречаются на протяжении всей жизни личинок, эти бактерии также постоянно идентифицируются из разных выборочных партий нормальных личинок, что указывает на то, что они являются коренными обитателями кишечника и могут быть настоящими симбионтами S.
Простота этого сообщества, состоящего из 22–42 OTU, особенно очевидна по сравнению с микробиотой кишечника насекомых из таких отрядов, как термиты Isoptera или позвоночные, которые часто являются носителями сотен филотипов [28], [29].Сильная щелочность в кишечнике, считающаяся важной детерминантой структуры сообщества личинок жуков, также может иметь место у Spodoptera , у которого pH средней кишки> 10 [22], [30]. Другие ключевые факторы, включая производительность фаст-фуда и функцию иммунной системы, также могут составлять таксономически ограниченную кишечную флору [31]. Однако три семейства, включая Enterococcaceae , Clostridiaceae и Enterobacteriaceae , особенно богаты и активны и, вероятно, являются основными функциональными популяциями, живущими внутри кишечника.Кроме того, обнаруживаются явные изменения в развитии (ранний возраст по сравнению с поздним возрастом). Мало того, что они постоянно встречаются на протяжении всей жизни личинок, эти бактерии также постоянно идентифицируются из разных выборочных партий нормальных личинок, что указывает на то, что они являются коренными обитателями кишечника и могут быть настоящими симбионтами S. littoralis .
littoralis .
В раннем возрасте наиболее активной группой была разнообразная группа Proteobacteria, особенно Gammaproteobacteria, в которой преобладали Pantoea и Citrobacter из семейства Enterobacteriaceae .Бактерии этого типа тесно связаны с насекомыми-травоядными и обладают широкими способностями к разложению полисахаридов [32] — [34]. Например, исследования, основанные на культивировании, показали, что Enterobacteriaceae , выделенные из кишечника тутового шелкопряда ( Bombyx mori ), обладают способностью секретировать ферменты, важные для переваривания сложных биополимеров пищевых растений, таких как целлюлоза, ксилан и пектин [32]. Геномный анализ штамма Pantoea из древесной оспы ( Sirex noctilio ) обнаружил гены, предположительно кодирующие углеводно-активные ферменты, причем большинство из них, как предполагается, активны в отношении гемицеллюлозы и более простых сахаров [33].Эти анализы роста in vitro, и геномные исследования дают начальную картину потенциальных метаболических возможностей, но не дают информации о бактериальной активности in situ . Наши результаты Pyro-SIP являются прямым доказательством метаболической активности микробов in vivo , подтверждая предыдущую гипотезу о том, что симбионты гаммапротеобактерий участвуют в деградации углеводов. Несмотря на свой низкий титр, Clostridium была другой высокоактивной группой личинок раннего возраста; эти облигатные анаэробы способны образовывать эндоспоры.Колонизация Clostridia обычно связана с его высокоэффективным перевариванием целлюлозы и его способностью сбраживать различные сахара [35]. Можно предположить, что аналогичную роль играют бактерии, связанные с листовой червью хлопчатника. Кроме того, Clostridia может повышать иммунитет хозяина [36]. В совокупности мы предполагаем, что наиболее активные Enterobacteriaceae и Clostridiaceae синергетически участвуют в пищеварительном процессе в кишечнике, способствуя расщеплению органических субстратов в листве и делая их более пригодными для пищеварения, абсорбции и метаболизма хозяина.
Наши результаты Pyro-SIP являются прямым доказательством метаболической активности микробов in vivo , подтверждая предыдущую гипотезу о том, что симбионты гаммапротеобактерий участвуют в деградации углеводов. Несмотря на свой низкий титр, Clostridium была другой высокоактивной группой личинок раннего возраста; эти облигатные анаэробы способны образовывать эндоспоры.Колонизация Clostridia обычно связана с его высокоэффективным перевариванием целлюлозы и его способностью сбраживать различные сахара [35]. Можно предположить, что аналогичную роль играют бактерии, связанные с листовой червью хлопчатника. Кроме того, Clostridia может повышать иммунитет хозяина [36]. В совокупности мы предполагаем, что наиболее активные Enterobacteriaceae и Clostridiaceae синергетически участвуют в пищеварительном процессе в кишечнике, способствуя расщеплению органических субстратов в листве и делая их более пригодными для пищеварения, абсорбции и метаболизма хозяина. Учитывая, что достаточное снабжение питательными веществами важно в раннем периоде жизни, высвобождаемые простые сахара будут непосредственно способствовать раннему развитию личинок, которые в дальнейшем могут быть ферментированы до различных других питательных веществ, таких как жирные кислоты с короткой цепью, что значительно улучшит экологическую среду кишечника. Эти бактерии, широко распространенные в природе, легко могут быть приобретены личинками во время кормления. Большая глубина отбора проб в этом исследовании выявила ряд редких таксонов в кишечнике. Хотя это разнообразие со временем уменьшилось, возможно, что редкие представители выполняют некоторые функции микробиоты или играют роль в особых ситуациях, как предполагали предыдущие исследования [37].Необходима дальнейшая работа, чтобы расшифровать их потенциальные биологические функции в кишечнике.
Учитывая, что достаточное снабжение питательными веществами важно в раннем периоде жизни, высвобождаемые простые сахара будут непосредственно способствовать раннему развитию личинок, которые в дальнейшем могут быть ферментированы до различных других питательных веществ, таких как жирные кислоты с короткой цепью, что значительно улучшит экологическую среду кишечника. Эти бактерии, широко распространенные в природе, легко могут быть приобретены личинками во время кормления. Большая глубина отбора проб в этом исследовании выявила ряд редких таксонов в кишечнике. Хотя это разнообразие со временем уменьшилось, возможно, что редкие представители выполняют некоторые функции микробиоты или играют роль в особых ситуациях, как предполагали предыдущие исследования [37].Необходима дальнейшая работа, чтобы расшифровать их потенциальные биологические функции в кишечнике.
На поздних стадиях микробиота кишечника стала еще проще с большим уменьшением микробного разнообразия и богатства. Более 97% членов сообщества принадлежали к Firmicutes, частично из-за значительного распространения Clostridium . Enterococcus преобладали и были определены как наиболее активная группа. Успешное распространение Firmicutes с течением времени, вероятно, в свою очередь, подавило рост бактерий из других типов в той же среде обитания, особенно Proteobacteria.Связанные с развитием личинок, изменения физиологии насекомых, такие как снижение окислительно-восстановительного потенциала и усиление иммунологического ответа хозяина, могут быть потенциальными движущими факторами наблюдаемых изменений. По мере роста личинок меньше кислорода может проникать в просвет кишечника через более толстую стенку кишечника и удлиненный пищеварительный тракт, что приводит к неизменно низкому напряжению кислорода в отделе кишечника [11]. Это в значительной степени аноксическое состояние, вероятно, способствует развитию облигатно-анаэробных Clostridium и факультативных анаэробных Enterococcus .
Enterococcus преобладали и были определены как наиболее активная группа. Успешное распространение Firmicutes с течением времени, вероятно, в свою очередь, подавило рост бактерий из других типов в той же среде обитания, особенно Proteobacteria.Связанные с развитием личинок, изменения физиологии насекомых, такие как снижение окислительно-восстановительного потенциала и усиление иммунологического ответа хозяина, могут быть потенциальными движущими факторами наблюдаемых изменений. По мере роста личинок меньше кислорода может проникать в просвет кишечника через более толстую стенку кишечника и удлиненный пищеварительный тракт, что приводит к неизменно низкому напряжению кислорода в отделе кишечника [11]. Это в значительной степени аноксическое состояние, вероятно, способствует развитию облигатно-анаэробных Clostridium и факультативных анаэробных Enterococcus .
Enterococcus , грамположительная молочнокислая бактерия (LAB), является единственным преобладающим родом в сообществе. ЛАБ являются общепризнанными полезными организмами микробиоты кишечника многих животных, включая насекомых [38]. Энтерококки по существу стабильны на протяжении всей жизни Spodoptera и вертикально передаются от матери к потомству через яйцеклетку, как показано для Manduca [9]. Следовательно, они первыми получают доступ к кишечнику и могут сразу же доминировать там после вылупления.Как вид-основатель, Enterococcus , вероятно, контролирует всю микробиоту вместе с хозяином, что может иметь решающее значение для создания отличительного и повторяющегося кишечного сообщества. Помимо листовой червей хлопчатника, Enterococcus присутствует у различных чешуекрылых, таких как личинка непарного шелкопряда ( Lymantria dispar ) и хлопковая совка ( Helicoverpa armigera ) как в полевых, так и в лабораторных образцах [39], [40]. ]. Примечательно, что Enterococcus , как было обнаружено, уже активны на яйцах табачного рогатого червя ( Manduca sexta ) на основе обнаружения рРНК [9].
ЛАБ являются общепризнанными полезными организмами микробиоты кишечника многих животных, включая насекомых [38]. Энтерококки по существу стабильны на протяжении всей жизни Spodoptera и вертикально передаются от матери к потомству через яйцеклетку, как показано для Manduca [9]. Следовательно, они первыми получают доступ к кишечнику и могут сразу же доминировать там после вылупления.Как вид-основатель, Enterococcus , вероятно, контролирует всю микробиоту вместе с хозяином, что может иметь решающее значение для создания отличительного и повторяющегося кишечного сообщества. Помимо листовой червей хлопчатника, Enterococcus присутствует у различных чешуекрылых, таких как личинка непарного шелкопряда ( Lymantria dispar ) и хлопковая совка ( Helicoverpa armigera ) как в полевых, так и в лабораторных образцах [39], [40]. ]. Примечательно, что Enterococcus , как было обнаружено, уже активны на яйцах табачного рогатого червя ( Manduca sexta ) на основе обнаружения рРНК [9]. Эти интимные ассоциации показывают, что Enterococcus может иметь важные функциональные последствия для чешуекрылых в целом. FISH показал, что большое количество энтерококков плотно прикрепляются к слизистому слою эпителия кишечника, образуя структуру, подобную биопленке, что может быть причиной его высокой стабильности в кишечнике здоровых личинок и, кроме того, может препятствовать кишечнику от вторжения вредных патогенных микробов [34], [41]. Помимо обеспечения физического барьера, изолированные штаммы показали сильное ингибирование против других бактерий.Новый антимикробный пептид был идентифицирован из бульонной культуры (Шао, неопубликованные данные). В совокупности эти результаты показывают, что Enterococcus , вероятно, является защитным симбионтом для здоровья хозяина. Поскольку хлопковая листовая червь является универсальной кормушкой в этой области, ее пищеварительный тракт постоянно подвергается воздействию потенциально вредных бактерий и грибковых эндофитов, которые он поглощает.
Эти интимные ассоциации показывают, что Enterococcus может иметь важные функциональные последствия для чешуекрылых в целом. FISH показал, что большое количество энтерококков плотно прикрепляются к слизистому слою эпителия кишечника, образуя структуру, подобную биопленке, что может быть причиной его высокой стабильности в кишечнике здоровых личинок и, кроме того, может препятствовать кишечнику от вторжения вредных патогенных микробов [34], [41]. Помимо обеспечения физического барьера, изолированные штаммы показали сильное ингибирование против других бактерий.Новый антимикробный пептид был идентифицирован из бульонной культуры (Шао, неопубликованные данные). В совокупности эти результаты показывают, что Enterococcus , вероятно, является защитным симбионтом для здоровья хозяина. Поскольку хлопковая листовая червь является универсальной кормушкой в этой области, ее пищеварительный тракт постоянно подвергается воздействию потенциально вредных бактерий и грибковых эндофитов, которые он поглощает. Энтерококки , имеющие структуру, подобную биопленке и проявляющие мощные противомикробные свойства, могут быть способны создавать эффект устойчивости к колонизации в кишечнике, который защищает хозяина от патогенов и широкого спектра некомменсальных конкурирующих микробов извне [2], [2] [ 42].Однако хорошо зарекомендовавшим себя энтерококкам не нужно полностью включать свой метаболический аппарат в полупустой лабораторной среде выращивания, что может объяснить их низкую метаболическую активность в раннем возрасте. Они не только связаны с улучшением питания и защитой от патогенов, но и активные бактерии могут также вносить вклад в другой метаболизм кишечника, например, выводить токсины из вредных аллелохимических веществ растительного происхождения и регулировать иммунный гомеостаз хозяина. Метагеномный анализ меченой ДНК из этого исследования расширит наши знания о других функциях, обеспечиваемых активной микробиотой.
Энтерококки , имеющие структуру, подобную биопленке и проявляющие мощные противомикробные свойства, могут быть способны создавать эффект устойчивости к колонизации в кишечнике, который защищает хозяина от патогенов и широкого спектра некомменсальных конкурирующих микробов извне [2], [2] [ 42].Однако хорошо зарекомендовавшим себя энтерококкам не нужно полностью включать свой метаболический аппарат в полупустой лабораторной среде выращивания, что может объяснить их низкую метаболическую активность в раннем возрасте. Они не только связаны с улучшением питания и защитой от патогенов, но и активные бактерии могут также вносить вклад в другой метаболизм кишечника, например, выводить токсины из вредных аллелохимических веществ растительного происхождения и регулировать иммунный гомеостаз хозяина. Метагеномный анализ меченой ДНК из этого исследования расширит наши знания о других функциях, обеспечиваемых активной микробиотой.
Во время совместной эволюции аборигенные кишечные бактерии приспособились работать вместе в этой особой экологической нише и приносить свои метаболические преимущества хозяину. С другой стороны, различные физико-химические условия в отношении развития хозяина, такие как щелочность кишечника, напряжение кислорода, могут влиять на активность сообщества, что, в свою очередь, влияет на его состав и, следовательно, на метаболические функции. Таким образом, и хозяин, и настоящие симбионты тщательно регулируют свою метаболическую активность и эффективность, чтобы сохранить мутуализм между хозяином и микробом. In vivo Измерение активности , предлагаемое SIP, помогает лучше понять изменения и поддержание микробного сообщества и дает более глубокое понимание роли активных членов в приспособленности хозяина. Наши данные также позволили провести первую углубленную инвентаризацию кишечной микробиоты модельного организма, состоящего в основном из фитофагов чешуекрылых. Знание кишечных бактерий такого крупного травоядного насекомого также может стать новыми целями для борьбы с сельскохозяйственными вредителями.
С другой стороны, различные физико-химические условия в отношении развития хозяина, такие как щелочность кишечника, напряжение кислорода, могут влиять на активность сообщества, что, в свою очередь, влияет на его состав и, следовательно, на метаболические функции. Таким образом, и хозяин, и настоящие симбионты тщательно регулируют свою метаболическую активность и эффективность, чтобы сохранить мутуализм между хозяином и микробом. In vivo Измерение активности , предлагаемое SIP, помогает лучше понять изменения и поддержание микробного сообщества и дает более глубокое понимание роли активных членов в приспособленности хозяина. Наши данные также позволили провести первую углубленную инвентаризацию кишечной микробиоты модельного организма, состоящего в основном из фитофагов чешуекрылых. Знание кишечных бактерий такого крупного травоядного насекомого также может стать новыми целями для борьбы с сельскохозяйственными вредителями.
Это пилотное исследование показывает, что Pyro-SIP может быстро получить представление не только о структуре сообщества, но, что более важно, о локальной метаболической активности его компонентов с высоким разрешением и высокой точностью, что обеспечивает отправную точку для исследований более сложных экосистем.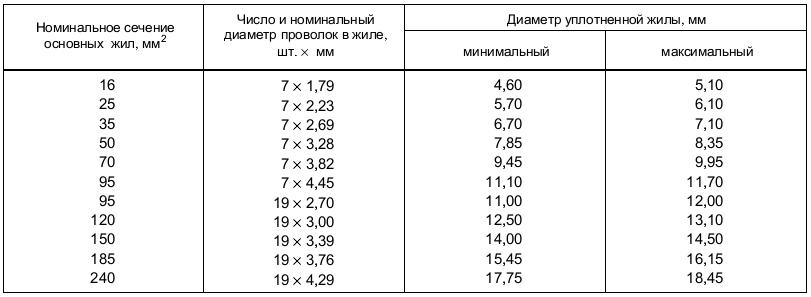 например, термиты или микробиота человека.Хотя наша работа была сосредоточена на распознавании общих активных кишечных бактерий путем отслеживания повсеместной глюкозы, аналогичный анализ может быть проведен на более конкретных источниках углерода, например, при маркировке защитных соединений растений для оценки активных бактерий, участвующих в процессе детоксикации хозяина. Таким образом, Pyro-SIP также предоставляет другой способ понять сложный метаболизм кишечника путем его расщепления. С развитием таких новых подходов скоро может произойти революция в понимании внутреннего мира жизни.
например, термиты или микробиота человека.Хотя наша работа была сосредоточена на распознавании общих активных кишечных бактерий путем отслеживания повсеместной глюкозы, аналогичный анализ может быть проведен на более конкретных источниках углерода, например, при маркировке защитных соединений растений для оценки активных бактерий, участвующих в процессе детоксикации хозяина. Таким образом, Pyro-SIP также предоставляет другой способ понять сложный метаболизм кишечника путем его расщепления. С развитием таких новых подходов скоро может произойти революция в понимании внутреннего мира жизни.
Материалы и методы
Выращивание насекомых и растений и сбор образцов
Spodoptera littoralis (яйца, приобретенные у Bayer Cropscience, Монхайм, Германия) были выведены и выращены на стерильной искусственной диете, состоящей из белой фасоли и основных питательных веществ без добавления антибиотиков или консервантов. Все растения хлопчатника ( Gossypium hirsutum DP90) выращивали в нашей теплице в стандартных условиях (23 ± 2 ° C; влажность 50 ± 5%; 16-часовой световой фотопериод). Для анализа состава сахара в кишечнике личинок выращивали на проростках хлопка в горшках или на свежем искусственном рационе в ящиках соответственно. После 7 дней кормления личинок промывали, успокаивали на льду и препарировали для сбора содержимого кишечника под препарирующим микроскопом. Пищеварительный канал был разделен на три области (передняя кишка, средняя кишка и задняя кишка) стерильными ножницами (рис. 1С), и содержимое кишечника из каждого отдела было выпущено в пробирки Эппендорфа. Материал из одного и того же сечения от пяти до восьми образцов был объединен, чтобы получить приблизительно 0.6 г на образец для экстракции сахара. Для анализа активных кишечных бактерий зондирование стабильных изотопов проводилось на личинках раннего возраста (от 1 st до 2 nd ) или личинках позднего возраста (5 th ) в течение 24 и 48 часов с использованием искусственной диеты с добавлением глюкозы. . Искусственную диету часто меняли, чтобы избежать заражения.
Для анализа состава сахара в кишечнике личинок выращивали на проростках хлопка в горшках или на свежем искусственном рационе в ящиках соответственно. После 7 дней кормления личинок промывали, успокаивали на льду и препарировали для сбора содержимого кишечника под препарирующим микроскопом. Пищеварительный канал был разделен на три области (передняя кишка, средняя кишка и задняя кишка) стерильными ножницами (рис. 1С), и содержимое кишечника из каждого отдела было выпущено в пробирки Эппендорфа. Материал из одного и того же сечения от пяти до восьми образцов был объединен, чтобы получить приблизительно 0.6 г на образец для экстракции сахара. Для анализа активных кишечных бактерий зондирование стабильных изотопов проводилось на личинках раннего возраста (от 1 st до 2 nd ) или личинках позднего возраста (5 th ) в течение 24 и 48 часов с использованием искусственной диеты с добавлением глюкозы. . Искусственную диету часто меняли, чтобы избежать заражения. Целые кишки личинок собирали, как указано выше, для выделения ДНК. Для каждого анализа эксперимент проводили в трех экземплярах.
Целые кишки личинок собирали, как указано выше, для выделения ДНК. Для каждого анализа эксперимент проводили в трех экземплярах.
Анализ состава сахара
Для характеристики растворимых органических соединений в пищеварительном канале, свеже собранное содержимое кишечника было извлечено в соответствии с опубликованной литературой [20], [43].Вкратце, образцы экстрагировали 2 мл бидистиллированной воды в термомиксере (Comfort, Eppendorf, Гамбург, Германия) при 60 ° C, 1400 об / мин в течение примерно 2 минут, а затем охлаждали на льду и гомогенизировали с помощью вращающегося смесителя (RM- Multi 1, STARLAB, Гамбург, Германия) в течение 6 ч при 4 ° C. Жидкости супернатанта (экстракты) отделяли центрифугированием (22000 × g, 5 мин при 4 ° C) и фильтровали (размер пор 0,22 мкм) для анализа растворимых сахарных соединений. Поскольку сахариды нелетучие, перед анализом методом ГХ-МС была проведена стадия дериватизации экстракта, как описано ранее [16].После реакции дериватизации 1 мкл супернатанта подвергали анализу GC-EIMS (Finnigan Trace GC-MS 2000, ThermoQuest, Egelsbach, Германия). Колонка Phenomenex ZB-5 (15 м × 0,25 мм, толщина пленки 0,25 мкм) была оборудована для разделения производных сахаров. Гелий использовался в качестве газа-носителя при расходе 1,5 мл мин. -1 . Температурная программа была установлена следующим образом: 80 ° C (2 мин), затем 15 ° C мин. -1 от до 300 ° C (6 мин). Данные были получены и обработаны с помощью программного обеспечения Xcalibur (Thermo Scientific, Саннивейл, Калифорния, США).Масс-спектры снимали в выбранном режиме ионного мониторинга (SIM) при 70 эВ. Все соединения были идентифицированы путем сравнения их времен удерживания и данных МС с аутентичными ссылками. Аликвоту экстракта также гидролизовали 2 М трифторуксусной кислотой (TFA; Sigma-Aldrich, Сент-Луис, Миссури, США) при 100 ° C в течение 2 часов, солюбилизируя матричные полисахариды в их моносахариды, а затем дериватизировали до соответствующего альдононитрила. ацетаты. Дальнейший анализ ГХ-МС был идентичен анализу негидролизованных экстрактов.
Колонка Phenomenex ZB-5 (15 м × 0,25 мм, толщина пленки 0,25 мкм) была оборудована для разделения производных сахаров. Гелий использовался в качестве газа-носителя при расходе 1,5 мл мин. -1 . Температурная программа была установлена следующим образом: 80 ° C (2 мин), затем 15 ° C мин. -1 от до 300 ° C (6 мин). Данные были получены и обработаны с помощью программного обеспечения Xcalibur (Thermo Scientific, Саннивейл, Калифорния, США).Масс-спектры снимали в выбранном режиме ионного мониторинга (SIM) при 70 эВ. Все соединения были идентифицированы путем сравнения их времен удерживания и данных МС с аутентичными ссылками. Аликвоту экстракта также гидролизовали 2 М трифторуксусной кислотой (TFA; Sigma-Aldrich, Сент-Луис, Миссури, США) при 100 ° C в течение 2 часов, солюбилизируя матричные полисахариды в их моносахариды, а затем дериватизировали до соответствующего альдононитрила. ацетаты. Дальнейший анализ ГХ-МС был идентичен анализу негидролизованных экстрактов. Концентрацию глюкозы проверяли с помощью набора для анализа глюкозы на основе специфического и чувствительного ферментативного метода в соответствии с протоколом, предоставленным производителем (GAHK-20, Sigma-Aldrich).
Концентрацию глюкозы проверяли с помощью набора для анализа глюкозы на основе специфического и чувствительного ферментативного метода в соответствии с протоколом, предоставленным производителем (GAHK-20, Sigma-Aldrich).
Экстракция и амплификация ДНК
Все свежевыобранные образцы кишечника сушили при 45 ° C в Speedvac (Concentrator 5301, Eppendorf) и измельчали в пробирке Eppendorf на 1,5 мл стерильным пластиковым пестиком. Геномную ДНК экстрагировали с использованием набора для выделения ДНК PowerSoil ™ (MO BIO Laboratories, Карлсбад, Калифорния, США) в соответствии с протоколом производителя.Дополнительный этап нагревания при 65 ° C в течение 10 минут был включен непосредственно перед взбиванием шариков. После очистки концентрацию ДНК определяли количественно с помощью спектрофотометра NanoVue (GE Healthcare Europe GmbH, Фрайбург, Германия). Успешная экстракция микробной метагеномной ДНК из кишечника была подтверждена с помощью ПЦР-анализов с общими бактериальными праймерами 16S рРНК (27f и 1492r) [30].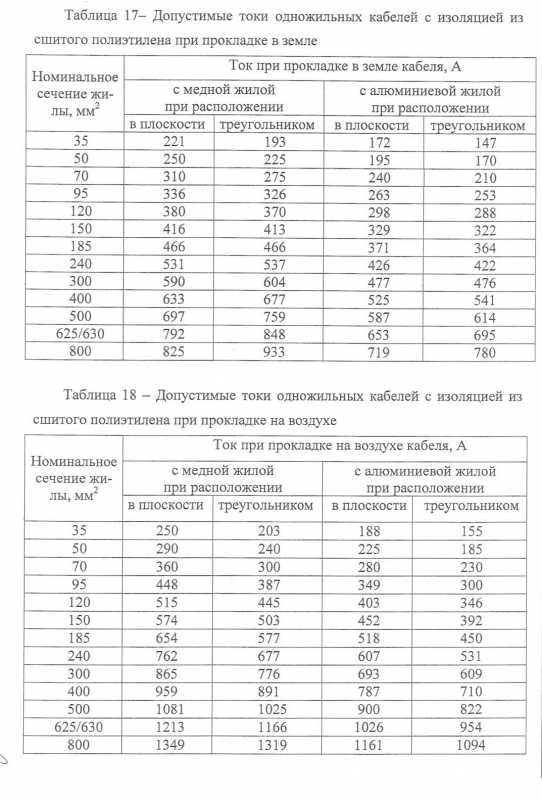 Реакция ПЦР описана ранее [11]. Праймеры, специфичные для архей и грибов, использовали для амплификации генов 16S архей или ITS грибов (Таблица S1) [44].Впоследствии экстрагированную ДНК использовали для измерения IRMS и ультрацентрифугирования в градиенте плотности.
Реакция ПЦР описана ранее [11]. Праймеры, специфичные для архей и грибов, использовали для амплификации генов 16S архей или ITS грибов (Таблица S1) [44].Впоследствии экстрагированную ДНК использовали для измерения IRMS и ультрацентрифугирования в градиенте плотности.
Измерение
13 C обогащения ДНК с помощью IRMS
Отношение изотопов углерода может быть надежным индикатором обогащения 13 C экстрагированными нуклеиновыми кислотами. Состав 13 C определяли с помощью сопряженного элементного анализатора / масс-спектрометрии соотношения изотопов (EA / IRMS). Очищенные образцы ДНК помещали в оловянные капсулы объемом 0,04 мл (3,5 × 5 мм, HEKAtech GmbH, Wegberg, Германия) и сушили в течение ночи.Образцы были полностью преобразованы в CO 2 , N 2 и H 2 O после сжигания (окисление при 1020 ° C, восстановление при 650 ° C) в постоянном потоке гелия (80 мл мин -1 ) путем с помощью элементного анализатора (EuroEA CN2 dual, HEKAtech GmbH). После прохождения водной ловушки (MgClO 4 ) газы разделяли хроматографически при 85 ° C и переносили через разделительный клапан в масс-спектрометр для измерения соотношения изотопов (IRMS) (IsoPrime, Micromass, Манчестер, Великобритания).Стандарт ацетанилида был откалиброван по международному стандарту Vienna Pee Dee Belemnite (VPDB) с использованием NBS 22 (эталонный материал МАГАТЭ) со значением δ 13 C, равным -30,03 ‰ [45]. В качестве заготовок использовались пустые оловянные капсулы. Каждый образец анализировали в трех экземплярах. Изотопные отношения выражаются в дельта-обозначениях следующим образом [46]:
После прохождения водной ловушки (MgClO 4 ) газы разделяли хроматографически при 85 ° C и переносили через разделительный клапан в масс-спектрометр для измерения соотношения изотопов (IRMS) (IsoPrime, Micromass, Манчестер, Великобритания).Стандарт ацетанилида был откалиброван по международному стандарту Vienna Pee Dee Belemnite (VPDB) с использованием NBS 22 (эталонный материал МАГАТЭ) со значением δ 13 C, равным -30,03 ‰ [45]. В качестве заготовок использовались пустые оловянные капсулы. Каждый образец анализировали в трех экземплярах. Изотопные отношения выражаются в дельта-обозначениях следующим образом [46]:
Разделение
13 C-меченой ДНК по плотности
Зависимое от плотности разрешение экстрагированной ДНК проводили в растворе хлорида цезия (CsCl, Sigma-Aldrich) со средней плотностью 1.725 мкг мл -1 с использованием опубликованного протокола [8]. Вкратце, приблизительно 1500 нг очищенной ДНК загружали в среду для центрифугирования CsCl, заполняли пробирку Quick-Seal на 5,1 мл (Beckmann, Fullerton, CA, USA) и закрывали. Все образцы были подготовлены с одной и той же партией среды CsCl и обрабатывались параллельно, чтобы минимизировать возможные вариации. 13 C-меченая ДНК была разделена изопикническим ультрацентрифугированием при 50 000 об / мин. в роторе NVT90 в ультрацентрифуге Optima L-90K (Beckmann) при 20 ° C в течение 40 часов в вакууме, а затем сформированные градиенты плотности фракционировали снизу вверх на 12 равных фракций (по 425 мкл каждая) путем вытеснения сверху водой с помощью насоса ВЭЖХ при скорости потока 850 мкл мин -1 (система Agilent HP1100, Вальдброн, Германия) (рис.2А). Плотность фракции определяли путем взвешивания аликвоты каждой фракции, и последняя фракция, содержащая воду, исключалась из анализа градиентной фракции по плотности. После этого ДНК осаждали из каждой фракции, повторно растворяли в воде, свободной от нуклеаз, и количественно определяли, как указано выше, для следующего анализа микробного сообщества. Распределение ДНК в данном образце рассчитывали как количество соответствующей фракции, деленное на общее количество ДНК всех фракций в образце, чтобы облегчить сравнение между градиентами.
Все образцы были подготовлены с одной и той же партией среды CsCl и обрабатывались параллельно, чтобы минимизировать возможные вариации. 13 C-меченая ДНК была разделена изопикническим ультрацентрифугированием при 50 000 об / мин. в роторе NVT90 в ультрацентрифуге Optima L-90K (Beckmann) при 20 ° C в течение 40 часов в вакууме, а затем сформированные градиенты плотности фракционировали снизу вверх на 12 равных фракций (по 425 мкл каждая) путем вытеснения сверху водой с помощью насоса ВЭЖХ при скорости потока 850 мкл мин -1 (система Agilent HP1100, Вальдброн, Германия) (рис.2А). Плотность фракции определяли путем взвешивания аликвоты каждой фракции, и последняя фракция, содержащая воду, исключалась из анализа градиентной фракции по плотности. После этого ДНК осаждали из каждой фракции, повторно растворяли в воде, свободной от нуклеаз, и количественно определяли, как указано выше, для следующего анализа микробного сообщества. Распределение ДНК в данном образце рассчитывали как количество соответствующей фракции, деленное на общее количество ДНК всех фракций в образце, чтобы облегчить сравнение между градиентами.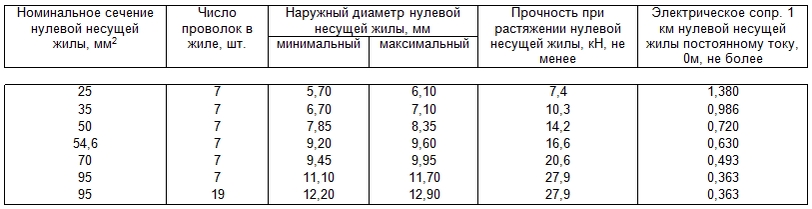
Отпечаток гена 16S рРНК в градиентных фракциях с помощью DGGE
Разделенные фракции градиента были проверены на различия в составе бактериального сообщества с помощью анализа денатурирующего градиентного гель-электрофореза (DGGE). Бактериальный ген 16S рРНК амплифицировали из всех фракций в два этапа в виде вложенной ПЦР с набором праймеров 27f / 1492r и 968F / 1401R (Таблица S1). ДГГЭ выполняли с помощью системы DCode (Bio-Rad, Мюнхен, Германия). Примерно равные количества продуктов ПЦР (300 нг) загружали на 8% полиакриламидный гель с градиентом денатурирующего агента от 20 до 80% (100% денатурант составлял 7 М мочевину и 40% (об. / Об.) Деионизированный формамид).Электрофорез проводили в буфере 1 × TAE при 100 В в течение 16 ч при 60 ° C, и гели окрашивали в течение 30 мин в буфере 0,5 × TAE с окрашиванием геля нуклеиновой кислоты SYBR-Gold (Invitrogen, Карлсруэ, Германия) для фотографирования. Паттерны DGGE сравнивали для оценки изменчивости структуры бактериального сообщества между меченым лечением и контролем.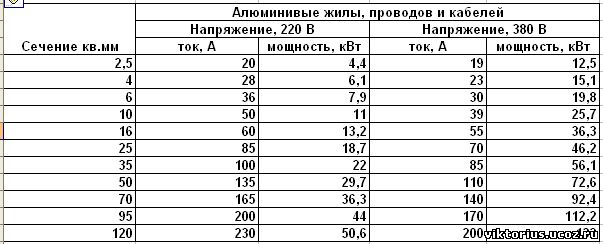 Поскольку профили DGGE показали разницу в градиентах меченой обработки, соответствующие фракции были объединены для получения одной тяжелой (4–5), одной средней (7) и одной легкой (9–11) фракции плотности ДНК из каждого образца.
Поскольку профили DGGE показали разницу в градиентах меченой обработки, соответствующие фракции были объединены для получения одной тяжелой (4–5), одной средней (7) и одной легкой (9–11) фракции плотности ДНК из каждого образца.
Пиросеквенирование ампликона FLX, кодируемое бактериальными метками (bTEFAP), и молекулярный филогенетический анализ
репрезентативных градиентов подвергали пиросеквенированию, как описано ранее [47], [48]. По сути, гипервариабельная часть V1 – V3 в 16S рДНК была амплифицирована с использованием пары праймеров Gray28F / 519r и секвенирована с использованием стратегии Roche 454 FLX Titanium (Таблица S1). Программный пакет Quantitative Insight into Microbial Ecology (QIIME, версия 1.4.0) использовался для обработки данных секвенирования и расчета разнообразия [49], [50].Последовательности сначала пропускались через контроль качества для удаления потенциальных артефактов и ошибок (в нашем анализе использовался скрипт Noise_wrapper.py) и обрезка детали с низким качеством [51].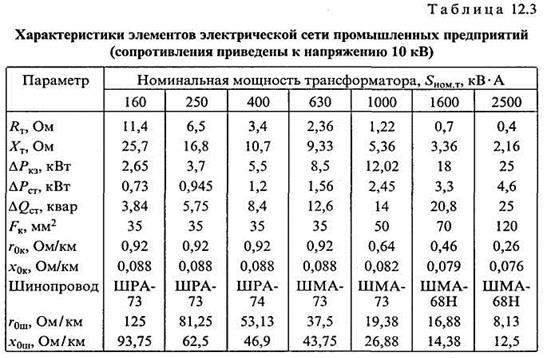 Позднее химеры (метод обнаружения: ChimeraSlayer) и считывания с низким содержанием (<0,1%) были удалены из анализа [52]. Качественные чтения были сгруппированы в операционные таксономические единицы (OTU) с использованием UCLUST с границами сходства 97% [53]. Для каждой OTU была извлечена одна репрезентативная последовательность, и классификатор RDP был использован для определения наивысшего разрешения таксономии на основе проекта базы данных рибосом.Наконец, была создана таблица OTU, описывающая наличие бактериальных филотипов в образце. Репрезентативные последовательности выравнивали с эталонными последовательностями, полученными из базы данных нуклеотидов NCBI, с использованием алгоритма ClustalW. Идентификация OTU, которые значительно различались по численности, проводилась в METASTATS с использованием непараметрического теста t по таксономическим данным, полученным из QIIME. Уровень значимости до порога (значение P) был установлен на уровне 0,05 [54]. Филогенетические деревья были рассчитаны с использованием метода максимального правдоподобия (модель Тамура-Неи) и метода объединения соседей с 1000 повторениями начальной загрузки в MEGA5 [55].
Позднее химеры (метод обнаружения: ChimeraSlayer) и считывания с низким содержанием (<0,1%) были удалены из анализа [52]. Качественные чтения были сгруппированы в операционные таксономические единицы (OTU) с использованием UCLUST с границами сходства 97% [53]. Для каждой OTU была извлечена одна репрезентативная последовательность, и классификатор RDP был использован для определения наивысшего разрешения таксономии на основе проекта базы данных рибосом.Наконец, была создана таблица OTU, описывающая наличие бактериальных филотипов в образце. Репрезентативные последовательности выравнивали с эталонными последовательностями, полученными из базы данных нуклеотидов NCBI, с использованием алгоритма ClustalW. Идентификация OTU, которые значительно различались по численности, проводилась в METASTATS с использованием непараметрического теста t по таксономическим данным, полученным из QIIME. Уровень значимости до порога (значение P) был установлен на уровне 0,05 [54]. Филогенетические деревья были рассчитаны с использованием метода максимального правдоподобия (модель Тамура-Неи) и метода объединения соседей с 1000 повторениями начальной загрузки в MEGA5 [55].
Флуоресценция
in situ Гибридизация (FISH)
Для локализации доминирующих кишечных симбионтов, FISH выполняли на 5 мкм тонких срезах холодной полимеризуемой смолы (Technovit 8100, Heraeus Kulzer GmbH, Wehrheim, Германия), внедренной в ткань кишечника. Специфичность зондов была проверена, и условия гибридизации были достигнуты, как описано [11]. Вскоре предметное стекло гибридизовали с 1,5 мМ каждого зонда (Таблица S1) в буфере для гибридизации, содержащем 900 мМ NaCl, 20 мМ Трис-HCl (pH 8.0), 20% формамида, 1% SDS. Для обнаружения использовали общий зонд эубактерий, меченный FITC, и зонд, специфичный для Cy3 Enterococcus , и получали изображения с помощью микроскопа Axio Imager Z1 (Carl Zeiss, Jena, Germany) [56] — [58].
Биотесты на антибиотики
Энтерококки были выделены из кишечника хлопкового червя с помощью Enterococcus -селективной чашки агара (Fluka, Мюнхен, Германия), и ген 16S рРНК был амплифицирован и секвенирован. Антимикробную активность выделенных штаммов против различных индикаторных бактерий оценивали с помощью диффузионного анализа в агаре.Надосадочную жидкость ночной культуры штамма E. mundtii в бульоне MRS (Roth, Карлсруэ, Германия) при 30 ° C доводили до pH 7,0 и фильтровали через PVDF-мембрану 0,22 мкм. Отверстия (диаметром 6 мм) вырезали из чашки с агаром BHI (Roth), инокулированной индикаторными штаммами ( Micrococcus luteus или Leuconostoc mesenteroides ), и заполняли 60 мкл фильтрата культуры E. mundtii . Чашки с агаром инкубировали при 30 ° C в течение 24 часов, и антимикробную активность определяли по образованию зон зазора вокруг загрузочных отверстий.
Антимикробную активность выделенных штаммов против различных индикаторных бактерий оценивали с помощью диффузионного анализа в агаре.Надосадочную жидкость ночной культуры штамма E. mundtii в бульоне MRS (Roth, Карлсруэ, Германия) при 30 ° C доводили до pH 7,0 и фильтровали через PVDF-мембрану 0,22 мкм. Отверстия (диаметром 6 мм) вырезали из чашки с агаром BHI (Roth), инокулированной индикаторными штаммами ( Micrococcus luteus или Leuconostoc mesenteroides ), и заполняли 60 мкл фильтрата культуры E. mundtii . Чашки с агаром инкубировали при 30 ° C в течение 24 часов, и антимикробную активность определяли по образованию зон зазора вокруг загрузочных отверстий.
Номера последовательностей нуклеотидов
Данные пиросеквенирования были депонированы в NCBI GenBank Short Read Archive под номером доступа. SRA057979.
Дополнительная информация
Рисунок S3.
Дактилоскопия DGGE фракций градиента с обнаруженной плотностью из метки 13 C-глюкозы и нативного контроля глюкозы ( 12 C-глюкоза). (a) Фракционно-зависимый ПЦР-анализ амплификации бактериального гена 16S рРНК.Фракции (1–12) были получены из выявленных градиентов плотности маркировки (+ 13 ° C) или контроля (+ 12 ° C). Повышенная интенсивность полос наблюдалась в тяжелых фракциях мечения. (b) профиль DGGE бактериальных генов 16S рРНК в градиентах обработки меткой и (c) контроль. Стрелки указывают на заметные изменения в составе сообщества маркированной обработки.
(a) Фракционно-зависимый ПЦР-анализ амплификации бактериального гена 16S рРНК.Фракции (1–12) были получены из выявленных градиентов плотности маркировки (+ 13 ° C) или контроля (+ 12 ° C). Повышенная интенсивность полос наблюдалась в тяжелых фракциях мечения. (b) профиль DGGE бактериальных генов 16S рРНК в градиентах обработки меткой и (c) контроль. Стрелки указывают на заметные изменения в составе сообщества маркированной обработки.
https://doi.org/10.1371/journal.pone.0085948.s004
(TIF)
Рисунок S4.
Обнаружение грибов и архей в кишечнике Spodoptera littoralis . Гель-электрофорез показывает отсутствие продуктов амплификации с праймерами, специфичными для грибов и архей. При попытке культивирования грибов использовали три вида обычных чашек с агаром для выращивания грибов (KM, агар Кемплера-МакКея; PDA, агар с картофельной декстрозой; PNM, питательная среда для растений), и грибок не был выделен.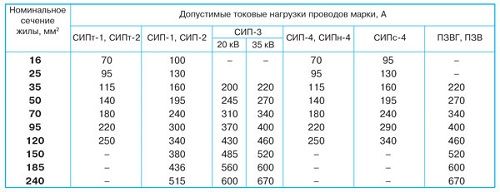
https://doi.org/10.1371/journal.pone.0085948.s005
(TIF)
Рисунок S5.
Изображения Enterococcus sp. из S. littoralis обнаруживают бактериальную локализацию кишечника и антимикробную активность. (a) FISH с Cy3-меченным зондом Enterococcus (желтый) показывает высокую плотность прилипания бактериальных клеток к слизистому слою, выстилающему эпителий кишечника, и это при более высоком увеличении (63 × 10) (b). Белая стрелка указывает на ткань эпителия кишечника.Белая стрелка указывает просвет кишечника. (c) Анализ диффузии в агаре показывает фильтраты культуры Enterococcus против Micrococcus luteus и (d) Leuconostoc mesenteroides . Антимикробная активность определяется по образованию зоны просвета вокруг загрузочного отверстия.
https://doi.org/10.1371/journal.pone.0085948.s006
(TIF)
Благодарности
Мы благодарим Анжелику Берг за помощь в лаборатории, Сайлендхарана Судакарана и Мартина Кальтенпота за помощь в пиросеквенировании. Благодарим Кристиана Коста за подробные комментарии к этой работе. Мы также благодарим Эмили Уиллер за помощь в редактировании.
Благодарим Кристиана Коста за подробные комментарии к этой работе. Мы также благодарим Эмили Уиллер за помощь в редактировании.
Вклад авторов
Задумал и спроектировал эксперименты: YS EAC WB. Проведены эксперименты: YS EAC HG SB. Проанализированы данные: YS EAC WB. Внесенные реактивы / материалы / инструменты анализа: SB. Написал статью: YS EAC WB.
Ссылки
- 1.
Клементе Дж. К., Урселл Л. К., Парфри Л. В., Найт Р. (2012) Влияние микробиоты кишечника на здоровье человека: комплексный взгляд.Cell 148: 1258–1270. - 2.
Диллон Р.Дж., Диллон В.М. (2004) Кишечные бактерии насекомых: непатогенные взаимодействия. Анну Преподобный Энтомол 49: 71–92. - 3.
Структура, функции и разнообразие микробиома здорового человека. Природа 486: 207–214. - 4.
Поульсен М., Сапунцис П. (2012) За каждым великим муравьем стоит большая кишка. Мол Экол 21: 2054–2057. - 5.
Reichardt N, Barclay AR, Weaver LT, Morrison DJ (2011) Использование стабильных изотопов для измерения метаболической активности кишечной микробиоты человека. Прикладная и экологическая микробиология 77: 8009–8014. - 6.
Dumont MG, Murrell JC (2005) Исследование стабильных изотопов — связь микробной идентичности с функцией. Nat Rev Microbiol 3: 499–504. - 7.
Radajewski S, Ineson P, Parekh NR, Murrell JC (2000) Исследование стабильных изотопов как инструмент микробной экологии. Nature 403: 646–649. - 8.
Neufeld JD, Vohra J, Dumont MG, Lueders T, Manefield M и др. (2007) Исследование стабильных изотопов ДНК. Nat Protoc 2: 860–866. - 9.
Бринкманн Н., Мартенс Р., Теббе С.С. (2008) Происхождение и разнообразие метаболически активных кишечных бактерий от лабораторно выведенных личинок Manduca sexta (Sphingidae, Lepidoptera, Insecta). Appl Environ Microbiol 74: 7189–7196. - 10.
Pilloni G, von Netzer F, Engel M, Lueders T (2011) Зависимая от акцептора электронов идентификация основных анаэробных разрушителей толуола в водоносном горизонте, загрязненном смолой и нефтью, с помощью Pyro-SIP. FEMS Microbiol Ecol - 11.
Тан Х, Фрейтак Д., Фогель Х, Пинг Л., Шао И и др.(2012) Сложность и изменчивость кишечной комменсальной микробиоты у многоядных личинок чешуекрылых. PLoS One 7: e36978. - 12.
Пинг Л., Бухлер Р., Митхофер А., Сватос А., Спителлер Д. и др. (2007) Новый белок типа Dps из кишечных бактерий насекомых катализирует гидролиз и синтез N-ациламинокислот. Environ Microbiol 9: 1572–1583. - 13.
Песек Дж., Бюхлер Р., Альбрехт Р., Боланд В., Зет К. (2011) Структура и механизм транслокации железа с помощью белка Dps из Microbacterium arborescens.J Biol Chem 286: 34872–34882. - 14.
Шао Ю., Спителлер Д., Тан Х, Пинг Л., Колси С. и др. (2011) Кристаллизация альфа- и бета-каротина в передней кишке личинок Spodoptera, питающихся токсичным пищевым растением. Биохимия насекомых, биол 41: 273–281. - 15.
Чжан В., Хэ HB, Чжан XD (2007) Определение нейтральных сахаров в почве с помощью капиллярной газовой хроматографии после дериватизации до ацетатов альдононитрила. Биология и биохимия почвы 39: 2665–2669. - 16.Price NP (2004) Производные ацильного сахара для ГХ / МС анализа обогащения 13C во время углеводного метаболизма. Anal Chem 76: 6566–6574.
- 17.
Lueders T, Manefield M, Friedrich MW (2004) Повышенная чувствительность исследования стабильных изотопов на основе ДНК и рРНК путем фракционирования и количественного анализа градиентов изопикнического центрифугирования. Environ Microbiol 6: 73–78. - 18.
Фирер Н., Джексон Р. Б. (2006) Разнообразие и биогеография почвенных бактериальных сообществ.Труды Национальной академии наук Соединенных Штатов Америки 103: 626–631. - 19.
Бенсон А.К., Келли С.А., Легге Р., Ма Ф., Лоу С.Дж. и др. (2010) Индивидуальность в составе микробиоты кишечника — это сложный полигенный признак, сформированный множеством факторов окружающей среды и генетических факторов хозяина. Proc Natl Acad Sci U S A 107: 18933–18938. - 20.
Drake HL, Wust PK, Horn MA (2011) Clostridiaceae и Enterobacteriaceae как активные ферментеры в содержимом кишечника дождевых червей. Isme Journal 5: 92–106. - 21.
Лу И, Конрад Р. (2005) Исследование стабильных изотопов in situ метаногенных архей в ризосфере риса. Наука 309: 1088–1090. - 22.
Funke M, Buchler R, Mahobia V, Schneeberg A, Ramm M и др. (2008) Быстрый гидролиз молекул, чувствительных к кворуму, в кишечнике личинок чешуекрылых. Chembiochem 9: 1953–1959. - 23.
Ковачева-Датчари П., Эгерт М., Маатуис А., Раджилич-Стоянович М., де Грааф А.А. и др. (2009) Связывание филогенетической идентичности бактерий с ферментацией крахмала в модели толстого кишечника in vitro с помощью зондирования стабильных изотопов на основе РНК.Environ Microbiol 11: 914–926. - 24.
Wong CN, Ng P, Douglas AE (2011) Бактериальное сообщество с низким разнообразием в кишечнике плодовой мушки Drosophila melanogaster. Environ Microbiol 13: 1889–1900. - 25.
Мартинсон В.Г., Данфорт Б.Н., Минкли Р.Л., Рюппелл О., Тингек С. и др. (2011) Простая и отличительная микробиота, связанная с медоносными пчелами и шмелями. Мол Экол 20: 619–628. - 26.
Робинсон CJ, Schloss P, Ramos Y, Raffa K, Handelsman J (2010) Устойчивость бактериального сообщества в средней кишке личинок белокочанной бабочки.Microb Ecol 59: 199–211. - 27.
Рассел Дж. А., Моро С. С., Гольдман-Уэртас Б., Фудзивара М., Ломан Д. Д. и др. (2009) Бактериальные симбионты кишечника тесно связаны с эволюцией травоядных муравьев. Proc Natl Acad Sci U S A 106: 21236–21241. - 28.
Варнеке Ф., Лугинбуль П., Иванова Н., Гассемиан М., Ричардсон Т. Х. и др. (2007) Метагеномный и функциональный анализ микробиоты задней кишки высших термитов, питающихся древесиной. Природа 450: 560–565. - 29.Перкинс Г.А., ден Баккер Х.С., Бертон А.Дж., Эрб Х.Н., Макдонау С.П. и др. (2012) В желудках лошадей содержится обильная и разнообразная микробиота слизистой оболочки. Appl Environ Microbiol 78: 2522–2532.
- 30.
Эгерт М., Стингл У., Бруун Л.Д., Поммеренке Б., Брюн А. и др. (2005) Структура и топология микробных сообществ в основных отделах кишечника личинок Melolontha melolontha (Coleoptera: Scarabaeidae). Appl Environ Microbiol 71: 4556–4566. - 31.
Pauchet Y, Wilkinson P, Vogel H, Nelson DR, Reynolds SE и др.(2010) Пиросеквенирование транскриптома средней кишки личинок Manduca sexta: сообщения для пищеварения, детоксикации и защиты. Насекомое Mol Biol 19: 61–75. - 32.
Ананд А.А.П., Веннисон С.Дж., Санкар С.Г., Прабху Д.И.Г., Васан П.Т. и др. (2010) Выделение и характеристика бактерий из кишечника Bombyx mori, разлагающих целлюлозу, ксилан, пектин и крахмал, и их влияние на пищеварение. Журнал насекомых 10. - 33.
Адамс А.С., Джордан М.С., Адамс С.М., Суен Дж., Гудвин Л.А. и др.(2011) Разрушающие целлюлозу бактерии, связанные с инвазивной древесной мятой Sirex noctilio. ISME J 5: 1323–1331. - 34.
Энгель П., Мартинсон В.Г., Моран Н.А. (2012) Функциональное разнообразие в простой кишечной микробиоте медоносной пчелы. Proc Natl Acad Sci U S A 109: 11002–11007. - 35.
Ватанабе Х., Токуда Г. (2010) Целлюлозолитические системы у насекомых. Анну Преподобный Энтомол 55: 609–632. - 36.
Атараси К., Тануэ Т., Шима Т., Имаока А., Кувахара Т. и др.(2011) Индукция регуляторных Т-клеток толстой кишки аборигенными видами Clostridium. Наука 331: 337–341. - 37.
Jones SE, Lennon JT (2010) Покой способствует поддержанию микробного разнообразия. Proc Natl Acad Sci U S A 107: 5881–5886. - 38.
Васкес А., Форсгрен Э., Фрис И., Пакстон Р.Дж., Флаберг Э. и др. (2012) Симбионты как основные модуляторы здоровья насекомых: молочнокислые бактерии и медоносные пчелы. PLoS One 7: e33188. - 39.
Прия Н.Г., Оджа А., Каджла М.К., Радж А., Раджагопал Р. (2012) Изменения кишечных бактерий Helicoverpa armigera, вызванные растением-хозяином.PLoS One 7. - 40.
Бродерик Н.А., Раффа К.Ф., Гудман Р.М., Хандельсман Дж. (2004) Перепись бактериального сообщества средней кишки личинки непарного шелкопряда с использованием культивирования и методов, не зависящих от культуры. Appl Environ Microbiol 70: 293–300. - 41.
Кох Х., Шмид-Хемпель П. (2011) Социально передаваемая кишечная микробиота защищает шмелей от кишечных паразитов. Proc Natl Acad Sci U S A 108: 19288–19292. - 42.
Сильверман Н., Пакетт Н. (2008) Иммунология.Правильные резидентные ошибки. Наука 319: 734–735. - 43.
Хорн М.А., Шрамм А., Дрейк Х.Л. (2003) Кишечник дождевого червя: идеальная среда обитания для проглоченных микроорганизмов, продуцирующих N2O. Appl Environ Microbiol 69: 1662–1669. - 44.
Андерсон И.К., Кэрни JWG (2004) Разнообразие и экология почвенных грибных сообществ: более глубокое понимание за счет применения молекулярных методов. Экологическая микробиология 6: 769–779. - 45.
Coplen TB, Brand WA, Gehre M, Groning M, Meijer HAJ и др.(2006) Новые рекомендации по измерениям дельты C-13. Anal Chem 78: 2439–2441. - 46.
Ленхарт К., Бунге М., Рейтинг С., Ной Т.Р., Шуттманн И. и др. (2012) Доказательства продукции метана сапротрофными грибами. Нац Коммуна 3: 1046. - 47.
Исхак Х.Д., Плаус Р., Сен Р., Келлнер К., Мейер Э. и др. (2011) Бактериальное разнообразие в колониях муравьев Solenopsis invicta и Solenopsis geminata, характеризуемых пиросеквенированием 16S ампликона 454. Microb Ecol 61: 821–831. - 48.Судакаран С., Салем Х, Кост С., Кальтенпот М. (2012) Географическая и экологическая стабильность симбиотической микробиоты среднего кишечника европейских огненных жуков, Pyrrhocoris apterus (Hemiptera, Pyrrhocoridae). Мол Экол
- 49.
Капорасо Дж. Г., Кучински Дж., Стомбо Дж., Биттингер К., Бушман Ф. Д. и др. (2010) QIIME позволяет анализировать высокопроизводительные данные секвенирования сообщества. Nat Методы 7: 335–336. - 50.
Карденас П.А., Купер П.Дж., Кокс М.Дж., Чико М., Ариас С. и др.(2012) Микробиота верхних дыхательных путей у детей с хрипом, ранее не получавших антибиотики, и у здоровых младенцев из тропиков сельских районов Эквадора. PLoS One 7: e46803. - 51.
Reeder J, Knight R (2010) Быстрое устранение шумов при пиросеквенировании считываний ампликонов путем использования распределений ранг-распространенность. Nat методы 7: 668–669. - 52.
Хаас Б.Дж., Геверс Д., Эрл А.М., Фельдгарден М., Уорд Д.В. и др. (2011) Образование и обнаружение последовательности химерной 16S рРНК в ПЦР-ампликонах Сэнгера и 454-пиросеквенировании.Genome Res 21: 494–504. - 53.
Edgar RC (2010) Поиск и кластеризация на порядки быстрее, чем BLAST. Биоинформатика 26: 2460–2461. - 54.
Уайт Дж. Р., Нагараджан Н., Поп М. (2009) Статистические методы обнаружения дифференциально обильных признаков в клинических метагеномных образцах. Плос вычислительная биология 5. - 55.
Тамура К., Петерсон Д., Петерсон Н., Стечер Г., Ней М. и др. (2011) MEGA5: Анализ молекулярной эволюционной генетики с использованием методов максимального правдоподобия, эволюционного расстояния и максимальной экономии. Mol Biol Evol 28: 2731–2739. - 56.
Manero A, Blanch AR (2002) Идентификация Enterococcus spp. основан на специфической гибридизации с зондами 16S рДНК. J Microbiol Methods 50: 115–121. - 57.
Бер Т., Куб С., Щедл М., Мелен А., Мейер Х и др. (2000) Вложенный массив зондов, нацеленных на рРНК, для обнаружения и идентификации энтерококков с помощью обратной гибридизации. Syst Appl Microbiol 23: 563–572. - 58.
Аманн Р.И., Биндер Б.Дж., Олсон Р.Дж., Чисхолм С.В., Деверо Р. и др.(1990) Комбинация олигонуклеотидных зондов, нацеленных на 16S рРНК, с проточной цитометрией для анализа смешанных микробных популяций. Appl Environ Microbiol 56: 1919–1925.
 Прикладная и экологическая микробиология 77: 8009–8014.
Прикладная и экологическая микробиология 77: 8009–8014. FEMS Microbiol Ecol
FEMS Microbiol Ecol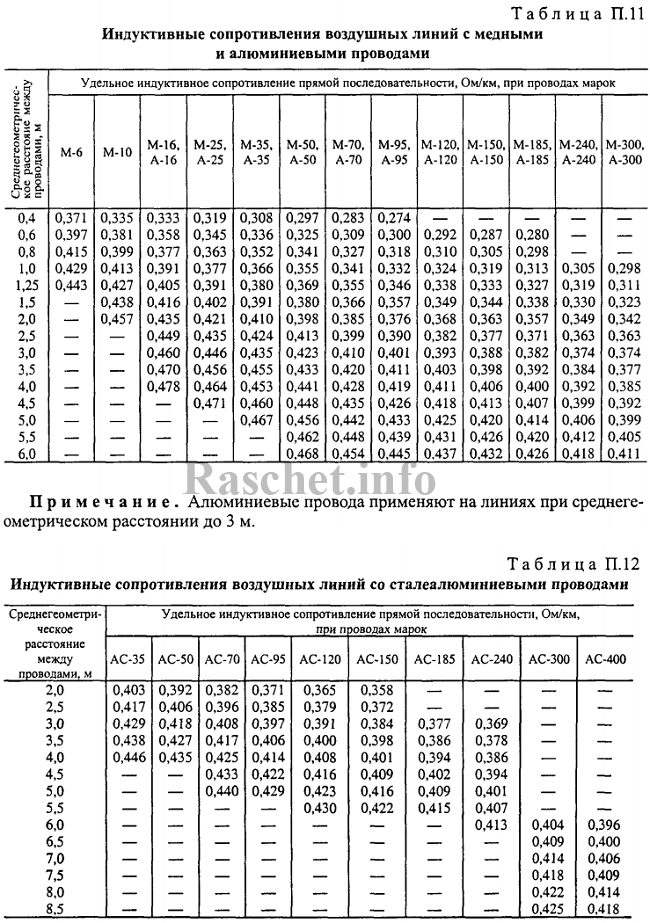 Биология и биохимия почвы 39: 2665–2669.
Биология и биохимия почвы 39: 2665–2669. Isme Journal 5: 92–106.
Isme Journal 5: 92–106.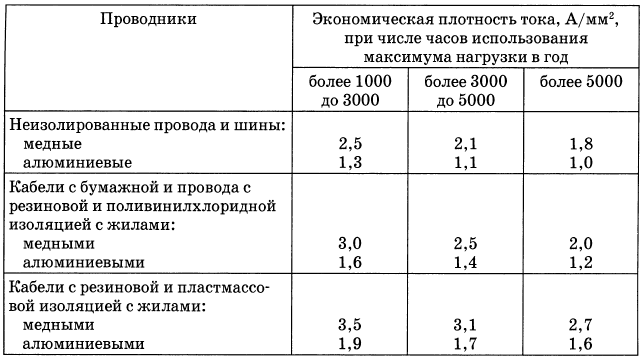 Мол Экол 20: 619–628.
Мол Экол 20: 619–628.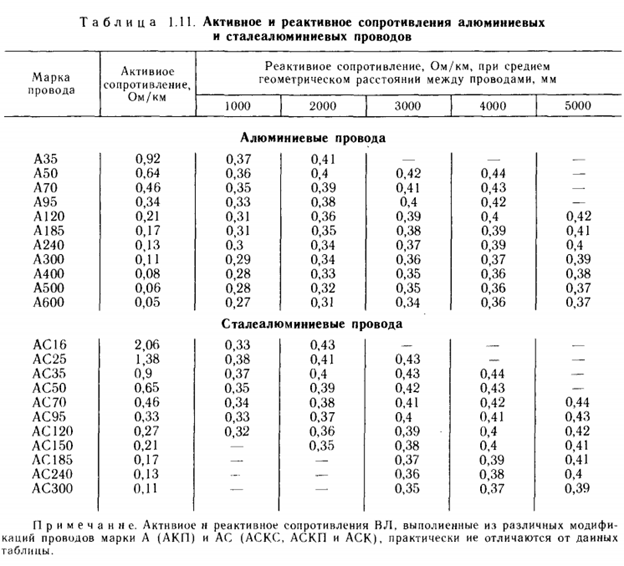 Appl Environ Microbiol 71: 4556–4566.
Appl Environ Microbiol 71: 4556–4566. (2010) Целлюлозолитические системы у насекомых. Анну Преподобный Энтомол 55: 609–632.
(2010) Целлюлозолитические системы у насекомых. Анну Преподобный Энтомол 55: 609–632. Appl Environ Microbiol 70: 293–300.
Appl Environ Microbiol 70: 293–300. и др. (2012) Доказательства продукции метана сапротрофными грибами. Нац Коммуна 3: 1046.
и др. (2012) Доказательства продукции метана сапротрофными грибами. Нац Коммуна 3: 1046. PLoS One 7: e46803.
PLoS One 7: e46803. Mol Biol Evol 28: 2731–2739.
Mol Biol Evol 28: 2731–2739.Прочный корпус SIP Call Box с питанием от PoE JR304-SC-VoIP — J&R Technology Ltd — Каталоги в формате PDF | Техническая документация
Телефон с громкой связью разработан для удовлетворения потребностей клиентов, потерявших в результате вандализма. Идеально подходит для парковок, тюрем, платформ железных дорог / метро, больниц, полицейских участков, банкоматов, стадионов, вне зданий и т. Д.Эти телефоны в корпусе из холоднокатаной стали и лицевой панели из нержавеющей стали, изготовленной в соответствии с высокими стандартами, обладают повышенной устойчивостью к вандализму и обеспечивают постоянное поддержание основной функции связи. Чрезвычайно проста в установке. Кабельный ввод находится на задней панели телефона, чтобы предотвратить искусственное повреждение, а клавиатура устойчива к погодным условиям и вандализму. Стандартный аналоговый телефон Как стандартный аналоговый телефон, это устройство обеспечивает основные телефонные функции и может быть подключено к любой стандартной двухпроводной телефонной линии, включая аналоговые линии PABX и PSTN.JR304-SC-OW Справочная служба • Стандартный аналоговый телефон или SIP-телефон • Прочный корпус из нержавеющей стали • Антивандальная металлическая кнопка • Работа в режиме громкой связи • 1 кнопка для быстрого набора • Для настенного монтажа, простая установка • Защита от атмосферных воздействий: вверх до IP65 • Внешний источник питания или PoE (SIP) • Порт RJ45 для подключения SIP • 4 винта с защитой от несанкционированного доступа для монтажа • Соответствие CE, FCC, RoHS ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ SIP-VoIP Вход адаптера: 100-240 В переменного тока, 50 ~ 60 Гц Выход: 5 В / Порт 1A: WAN: 10 / 100Base-T RJ-45 для LAN, Auto MDIX LAN: 10 / 100Base-T RJ-45 для ПК, Auto MDIX Энергопотребление: в режиме ожидания: 1.
Идеально подходит для парковок, тюрем, платформ железных дорог / метро, больниц, полицейских участков, банкоматов, стадионов, вне зданий и т. Д.Эти телефоны в корпусе из холоднокатаной стали и лицевой панели из нержавеющей стали, изготовленной в соответствии с высокими стандартами, обладают повышенной устойчивостью к вандализму и обеспечивают постоянное поддержание основной функции связи. Чрезвычайно проста в установке. Кабельный ввод находится на задней панели телефона, чтобы предотвратить искусственное повреждение, а клавиатура устойчива к погодным условиям и вандализму. Стандартный аналоговый телефон Как стандартный аналоговый телефон, это устройство обеспечивает основные телефонные функции и может быть подключено к любой стандартной двухпроводной телефонной линии, включая аналоговые линии PABX и PSTN.JR304-SC-OW Справочная служба • Стандартный аналоговый телефон или SIP-телефон • Прочный корпус из нержавеющей стали • Антивандальная металлическая кнопка • Работа в режиме громкой связи • 1 кнопка для быстрого набора • Для настенного монтажа, простая установка • Защита от атмосферных воздействий: вверх до IP65 • Внешний источник питания или PoE (SIP) • Порт RJ45 для подключения SIP • 4 винта с защитой от несанкционированного доступа для монтажа • Соответствие CE, FCC, RoHS ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ SIP-VoIP Вход адаптера: 100-240 В переменного тока, 50 ~ 60 Гц Выход: 5 В / Порт 1A: WAN: 10 / 100Base-T RJ-45 для LAN, Auto MDIX LAN: 10 / 100Base-T RJ-45 для ПК, Auto MDIX Энергопотребление: в режиме ожидания: 1. 5 Вт / активный: 1,8 Вт Основной чип: функции Broadcom Voice • Поддержка 2-х линий SIP, SIP 2.0 (RFC3261) • Кодек: G.711A / u, G.7231 высокий / низкий, G.729, G.722, G.726 • Подавление эха: поддержка G.168 и 96 мс без использования рук. • Поддержка полного дуплекса. • Поддержка SIP домена SIP, аутентификации SIP (нет, базовая, MD5), DNS-имя сервера, одноранговая сеть • Поддержка SIP 2 сервера, пользователь может через каждый сервер вызывать и исходить Сетевые функции • WAN / LAN: поддержка режима моста • Поддержка PPPoE для xDSL. • Поддержка VLAN.com Веб: www.jrtele.com Добавить: Строение № 1, Индустриальный парк Цзюньхэ, Даланг, город Лунхуа, район Баоань
5 Вт / активный: 1,8 Вт Основной чип: функции Broadcom Voice • Поддержка 2-х линий SIP, SIP 2.0 (RFC3261) • Кодек: G.711A / u, G.7231 высокий / низкий, G.729, G.722, G.726 • Подавление эха: поддержка G.168 и 96 мс без использования рук. • Поддержка полного дуплекса. • Поддержка SIP домена SIP, аутентификации SIP (нет, базовая, MD5), DNS-имя сервера, одноранговая сеть • Поддержка SIP 2 сервера, пользователь может через каждый сервер вызывать и исходить Сетевые функции • WAN / LAN: поддержка режима моста • Поддержка PPPoE для xDSL. • Поддержка VLAN.com Веб: www.jrtele.com Добавить: Строение № 1, Индустриальный парк Цзюньхэ, Даланг, город Лунхуа, район Баоань
.