
Натриевый равновесный потенциал — Справочник химика 21
Пропуская элект]жческий ток через введенный в клетку электрод, можно деполяризовать мембрану аксоиа в каком-то одном участке (рис. 18-20). Если сила тока мала,-деполяризация будет подпороговой натриевые каналы останутся закрытыми и потенциал действия не возникнет. Установится равновесное состояние, при котором ток, протекающий через микроэлектрод внутрь клетки, сбалансирован током, текущим наружу через мембрану. Часть тока будет выходить вблизи электрода, а часть, прежде чем выйти из клетки, пройдет некоторое расстояние внутри аксона в том или другом иаправлении. Сила тока, проходящего по какому-либо из этих путей, будет зависеть от их сопротивления. Так как цитоплазма аксоиа оказывает некоторое сопротивление току, сила тока будет наибольшей вблизи микроэлектрода н наименьшей вдали от него. При таком распределении тока сдвиг мембранного потеициала уменьшается экспоненциально с увеличением расстояния от источника возмущения. Такого рода пассивное распространение электрического сигнала вдоль нервного волокна-без какого-либо усиления, обусловленного открытием потенциал-зависимых каналов,-аналогично распространению сигнала по телеграфному кабелю, лежащему на дне моря. По мере прохождения тока по осевому проводнику (цитоплазме) происходит некоторая утечка его через слой изоляции (мембрану) в окружающую среду, так что сигнал постепенно затухает. Поэтому элект жческие свойства, от которых зависит пассивное распространение сигналов, часто называют кабельными свойствами аксона. [c.88]
Такого рода пассивное распространение электрического сигнала вдоль нервного волокна-без какого-либо усиления, обусловленного открытием потенциал-зависимых каналов,-аналогично распространению сигнала по телеграфному кабелю, лежащему на дне моря. По мере прохождения тока по осевому проводнику (цитоплазме) происходит некоторая утечка его через слой изоляции (мембрану) в окружающую среду, так что сигнал постепенно затухает. Поэтому элект жческие свойства, от которых зависит пассивное распространение сигналов, часто называют кабельными свойствами аксона. [c.88]
Пока представление о потенциале действия носило феноменологический характер, в дальнейшем необходимо рассмотреть лежащие в его основе молекулярные процессы. В гл. 6 эти вопросы обсуждаются подробно, здесь же рассмотрим лишь некоторые из них. В начале 50-х гг. английские физиологи Ходжкин и Хаксли исследовали потенциал действия и заложили основы современного понимания данного явления.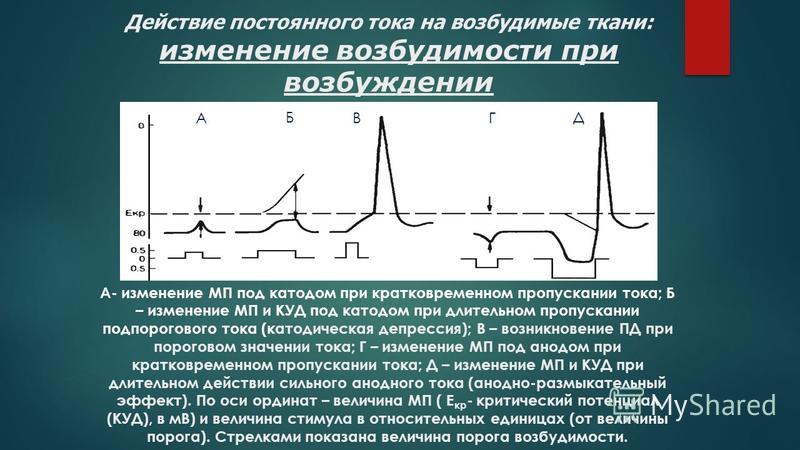 Они показали, что первоначально падение потенциала (деполяризация) обусловлено утечкой ионов натрия (рис. 5.7). По достижении порогового значения ионные каналы в мембране открываются и пропускают ионы натрия. Последующая реполяризация происходит благодаря открытию специальных калиевых каналов и протока ионов калия в обратном направлении, т. е. изнутри наружу, одновременно закрываются натриевые каналы (инактивация). Из рис. 5.7 следует, что первоначально реполяризация превышает значение потенциала покоя, так как при равновесном потенциале для К+ мембрана характеризуется более высоким отрицательным зарядом, чем при потенциале покоя. Это наблюдаемое различие медленно исчезает в результате закрывания калиевого канала и восстановления натриевого потенциала покоя. Инактивация [c.117]
Они показали, что первоначально падение потенциала (деполяризация) обусловлено утечкой ионов натрия (рис. 5.7). По достижении порогового значения ионные каналы в мембране открываются и пропускают ионы натрия. Последующая реполяризация происходит благодаря открытию специальных калиевых каналов и протока ионов калия в обратном направлении, т. е. изнутри наружу, одновременно закрываются натриевые каналы (инактивация). Из рис. 5.7 следует, что первоначально реполяризация превышает значение потенциала покоя, так как при равновесном потенциале для К+ мембрана характеризуется более высоким отрицательным зарядом, чем при потенциале покоя. Это наблюдаемое различие медленно исчезает в результате закрывания калиевого канала и восстановления натриевого потенциала покоя. Инактивация [c.117]
Из рассказа о потенциале покоя и потенциале действия вы знаете, что для каждого иона существует свой равновесный потенциал, при котором число ионов, входящих в клетку и выходящих из клетки, становится одинаковым. В покое для ионов калия равновесный потенциал равен примерно —80 мВ при возбуждении, когда в основном открываются натриевые каналы, равновесный потенциал для натрия равен примерно +40 мВ. У постсинаптической мембраны тоже есть свой равновесный потенциал. Его величина зависит от того, какие ионы пропускает эта мембрана. Например, постсинаптическая мембрана возбуждающего синапса, каналы которой в равной мере пропускают и калий, и натрий, имеет равновесный потенциал, лежащий ровно посередине между таковыми для калия и натрия (—80 + 40)/2 = —20 мВ. А у тормозного синапса, пропускающего ионы хлора, равновесный потенциал равен примерно —80 мВ. [c.173]
В покое для ионов калия равновесный потенциал равен примерно —80 мВ при возбуждении, когда в основном открываются натриевые каналы, равновесный потенциал для натрия равен примерно +40 мВ. У постсинаптической мембраны тоже есть свой равновесный потенциал. Его величина зависит от того, какие ионы пропускает эта мембрана. Например, постсинаптическая мембрана возбуждающего синапса, каналы которой в равной мере пропускают и калий, и натрий, имеет равновесный потенциал, лежащий ровно посередине между таковыми для калия и натрия (—80 + 40)/2 = —20 мВ. А у тормозного синапса, пропускающего ионы хлора, равновесный потенциал равен примерно —80 мВ. [c.173]
Из представленных рисунков следует, что, во-первых чем ближе смещается к значению равновесного потенциала, определяемого по уравнению Нернста (3.2) для ионов Ма+ и К+, тем меньше значение соответствующего тока. Так, при стремлении ф к -I- 60 мВ натриевый ток падает до нуля. То же происходит и с калиевым током при достижении ф величины — 60 мВ. И во-вторых, меняется временной ход IJ и 1 при изменениях. Так, ток натрия при -> ф быстрее уменьшается до нуля (более [c.92]
И во-вторых, меняется временной ход IJ и 1 при изменениях. Так, ток натрия при -> ф быстрее уменьшается до нуля (более [c.92]
Именно так должен был вести себя ток, обусловленный ионами Na+ движущей силой для этих ионов служит натриевый равновесный потенциал, составляющий примерно +55 мВ. [c.156]
Благодаря высокому перенапряжению водорода на ртути на ртутном катоде удается восстановить даже натрий. Натрий растворяется в ртути, и равновесный потенциал амальгамного натриевого электрода значительно менее отрицателен, чем потенциал натриевого электрода. [c.219]
По тем же самым причинам при увеличении проводимости для ионов Na (g Na) мембранный потенциал сместится к новому уровню, близкому к равновесному потенциалу для натрия (рис. 18-8, ). В действительности именно это и происходит на короткий момент во время прохождения импульса (потенциала действия) в результате открывания потенциал-зависимых натриевых каналов. В отличие от каналов утечки этн каналы почти все закрыты, когда нейрон находится в состоянии покоя, и открываются только при изменении мембранного потенциала. Свойства потенциал-зависимых натриевых каналов будут подробно рассмотрены ниже. [c.79]
В отличие от каналов утечки этн каналы почти все закрыты, когда нейрон находится в состоянии покоя, и открываются только при изменении мембранного потенциала. Свойства потенциал-зависимых натриевых каналов будут подробно рассмотрены ниже. [c.79]
Потенциал на мембране в состоянии покоя (ХХШ.1.2) приближается к равно-весному потенциалу для К+, так как проницаемости для К+ в этих условиях намного больше коэффициента проницаемости для Ма (Рк -Рма = 1 0,04). Генерация импульса. Реверсия мембранного потенциала во время развития импульса вызвана тем, что на гребне спайка потенциала действия мембрана избирательно проницаема для Ма , так что отношение ионных проницаемостей становится равным Рк Рма = 1 20. Как видно из формулы (ХХШ.1.2), в этих условиях ( Ма Рк) потенциал на мембране приближается к равновесному натриевому потенциалу (рма- Разность потенциалов на мембране, проницаемой только для Ма , выражается формулой Нернста [c.167]
Ответ. Калиевый и натриевый токи всегда направлены так, чтобы вернуть потенциал к соответствующему равновесному значению калий тянет к —80 мВ, а натрий — к -f 40 мВ.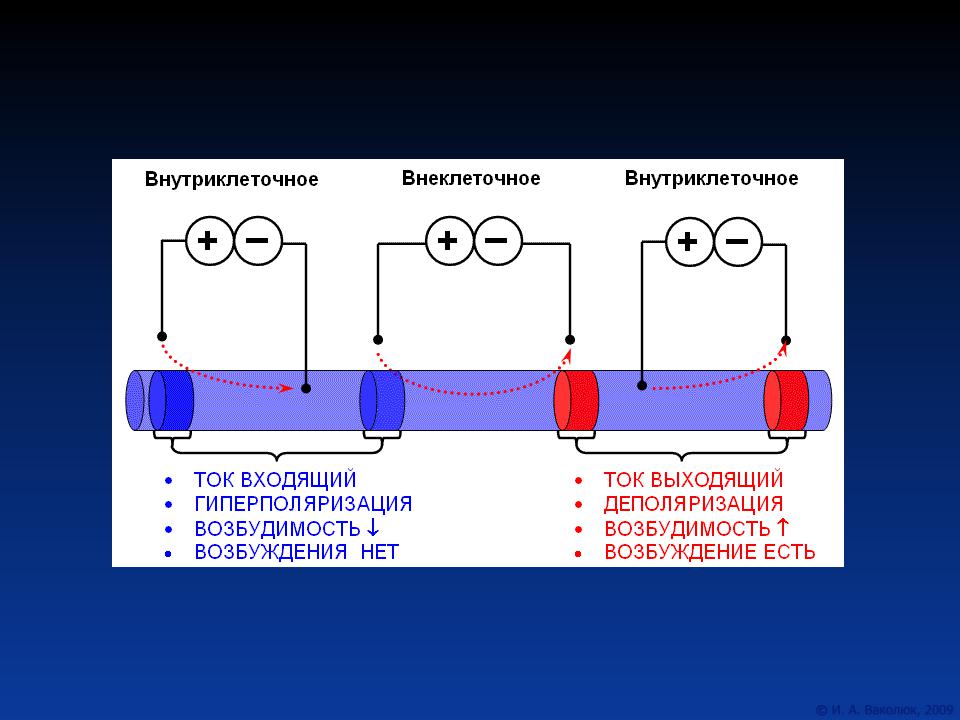 Поэтому для значений МП в интервале от —80 до — -40 мВ эти токи всегда направлены противоположно. Значение МП, при котором калиевый и натриевый токи уравновешивают друг друга,— это и есть ПП. [c.94]
Поэтому для значений МП в интервале от —80 до — -40 мВ эти токи всегда направлены противоположно. Значение МП, при котором калиевый и натриевый токи уравновешивают друг друга,— это и есть ПП. [c.94]
После пика потенциала действия мембранный потенциал возвращается к отрицательной величине, которая зависит от внеклеточной концентрации К и даже ближе к равновесному калиевому потенциалу, чем потенциал покоя мембрана утрачивает проницаемость для натрия, тогда как проницаемость для калия возрастает, т.е. натриевые каналы закрываются, а дополнительные калиевые открываются. [c.299]
Передача электрических сигналов нервной клеткой основана на изменении мембранного потенциала в результате прохождения небольших количеств ионов через управляемые ионные каналы. Эти ионы перемещаются за счет энергии, большой запас которой создается благодаря работе натриево-калиевого насоса, поддерживающего высокие градиенты концентрации На и 1С на мембране нервной клетки. В состоянии покоя мембрана нейрона благодаря каналам утечки более проницаема дм калия, чем для других ионов, и поэтому мембранный потенциал близок к равновесному калиевому потенциалу, составляющему примерно — 70 мВ. Потенциал действия возникает тогда, когда под влиянием короткого деполяризующего стимула открываются потенциал-зависимые натриевые каналы, так что мембрана становится более проницаемой для На а мембранный потенциал еще дальше смещается в сторону равновесного натриевого потенциала Благодаря такой положительной обратной связи открывается еще больше натриевых каналов, что в конечном итоге приводит к возникновению потенциала действия, подчиняющегося закону всё или ничего . На каждом данном участке мембраны потенциал действия быстро исчезает вследствие инактивации натриевых каналов, а во многих нейронах также вследствие открытия потенциал-зависимых калиевых каналов. [c.304]
В состоянии покоя мембрана нейрона благодаря каналам утечки более проницаема дм калия, чем для других ионов, и поэтому мембранный потенциал близок к равновесному калиевому потенциалу, составляющему примерно — 70 мВ. Потенциал действия возникает тогда, когда под влиянием короткого деполяризующего стимула открываются потенциал-зависимые натриевые каналы, так что мембрана становится более проницаемой для На а мембранный потенциал еще дальше смещается в сторону равновесного натриевого потенциала Благодаря такой положительной обратной связи открывается еще больше натриевых каналов, что в конечном итоге приводит к возникновению потенциала действия, подчиняющегося закону всё или ничего . На каждом данном участке мембраны потенциал действия быстро исчезает вследствие инактивации натриевых каналов, а во многих нейронах также вследствие открытия потенциал-зависимых калиевых каналов. [c.304]
Потенциал действия возникает, когда мембрана мгновенно деполяризуется до уровня, превышающего определенный порог. Как уже говорилось в гл. 6, в результате такой деполяризации какого-то участка мембраны здесь откроются потенциал-зависимые натриевые каналы, что вызовет ток ионов Ка вниз по их электрохимическому градиенту следствием будет дальнейшая деполяризация мембраны, в результате чего откроется еще большее число Ка -каналов, и так далее, подобно цепной реакции, до тех пор, пока потенциал в этом участке мембраны не приблизится к натриевому равновесному потенциал (см. схему 19-1). На этом этапе происходят два события, которые возвращают потенциал мембраны к первоначальному отрицательному значению Ка -каналы спонтанно переходят в закрытое, инактивированное состояние, а потенциалзависимые К -каналы открываются. Эти калиевые каналы реагируют на изменение мембранного потенциала почти так же, как и натриевые, но менее быстро, и поэтому их иногда называют меОленными К -каналами Как только К -каналы открываются, выходящий калие- [c.298]
Как уже говорилось в гл. 6, в результате такой деполяризации какого-то участка мембраны здесь откроются потенциал-зависимые натриевые каналы, что вызовет ток ионов Ка вниз по их электрохимическому градиенту следствием будет дальнейшая деполяризация мембраны, в результате чего откроется еще большее число Ка -каналов, и так далее, подобно цепной реакции, до тех пор, пока потенциал в этом участке мембраны не приблизится к натриевому равновесному потенциал (см. схему 19-1). На этом этапе происходят два события, которые возвращают потенциал мембраны к первоначальному отрицательному значению Ка -каналы спонтанно переходят в закрытое, инактивированное состояние, а потенциалзависимые К -каналы открываются. Эти калиевые каналы реагируют на изменение мембранного потенциала почти так же, как и натриевые, но менее быстро, и поэтому их иногда называют меОленными К -каналами Как только К -каналы открываются, выходящий калие- [c.298]
Однако экспериментально было зарегистрировано значение только —70 мВ. Чем вызвано такое расхождение До сих пор мы считали, что мембрана пропускает только К+, но не пропускает Ыа+. Но в действительности мембрана в некоторой степени проницаема и для Ыа+, а так как для концентрации Ыа+ характерен противоположный градиент по сравнению с градиентом концентрации К» «, поток ионов натрия в противоположном потоку ионов калия направлении уменьшает измеряемый потенциал мембраны (такое явление называется потоком покоя для отличия от натриевого потока при возбуждении мембраны) (рис. 5.2). Это достаточно просто доказать экспериментально если Ыа+ заменить на объемный ион, например холин [(СНз)зК-СН2-СН20Н]+, то мембранный потенциал окажется равным —75 мВ, т. е. устанавливается теоретическое значение калиевого равновесного потенциала, так как ион хо-лина не проникает через мембрану из-за своих размеров. [c.113]
Чем вызвано такое расхождение До сих пор мы считали, что мембрана пропускает только К+, но не пропускает Ыа+. Но в действительности мембрана в некоторой степени проницаема и для Ыа+, а так как для концентрации Ыа+ характерен противоположный градиент по сравнению с градиентом концентрации К» «, поток ионов натрия в противоположном потоку ионов калия направлении уменьшает измеряемый потенциал мембраны (такое явление называется потоком покоя для отличия от натриевого потока при возбуждении мембраны) (рис. 5.2). Это достаточно просто доказать экспериментально если Ыа+ заменить на объемный ион, например холин [(СНз)зК-СН2-СН20Н]+, то мембранный потенциал окажется равным —75 мВ, т. е. устанавливается теоретическое значение калиевого равновесного потенциала, так как ион хо-лина не проникает через мембрану из-за своих размеров. [c.113]
Таким образом, суммарный ток ионов Na равен нулю, если мембранный потенциал, называемый в данном случае натриевым равновесным потенцш-лом, N3. равен 581gl(,([Na ](,/[Na ]i) мВ. Точно так же суммарный ток ионов К будет равен нулю при значении мембранного потенциала 581og o([K ]o/[K ]i) мВ, что соответствует калиевому равновесному потенциалу, Кк. Обычно значения лежат в области между -I- 50 и -1-65 мВ, а Кк-между -70 и — 100 мВ. [c.78]
равен 581gl(,([Na ](,/[Na ]i) мВ. Точно так же суммарный ток ионов К будет равен нулю при значении мембранного потенциала 581og o([K ]o/[K ]i) мВ, что соответствует калиевому равновесному потенциалу, Кк. Обычно значения лежат в области между -I- 50 и -1-65 мВ, а Кк-между -70 и — 100 мВ. [c.78]
Когда клеточная мембрана, содержащая много натриевых каналов, частично деполяризуется в ответ на кратковременный стимул, некоторые каналы быстро открываются, пропуская внутрь клетки ионы Ыа . Переход положительных зарядов внутрь ведет к дальнейшей деполяризации мембраны, и в результате открывается еще больше каналов, пропускающих в клетку ионы N8 . Этот самоусиливающийся процесс продолжается до тех пор, пока мембранный потенциал, составлявший вначале -70 мВ (потенциал покоя) не дойдет до + 50 мВ (равновесный потенциал Na ). В этот момент, когда суммарная электрохимическая движущая сила для ионов N3 равна нулю, клетка могла бы перейти в новое состояние покоя, при котором все натриевые каналы постоянно открыты, если бы открытая конформация каналов была стабильной. От такого длительного электрического спазма клетку спасает автоматическая инактивация натриевых каналов, которые постепенно, один за другим, закрываются и остаются закрытыми, пока мшбранный потенциал не вернется к исходной отрицательной величине, т. е. к уровню покоя. Весь цикл от момента воздействия стимула до возвращения к состоянию покоя занимает всего лишь несколько миллисекунд или даже меньше (рис. 18-17). [c.85]
От такого длительного электрического спазма клетку спасает автоматическая инактивация натриевых каналов, которые постепенно, один за другим, закрываются и остаются закрытыми, пока мшбранный потенциал не вернется к исходной отрицательной величине, т. е. к уровню покоя. Весь цикл от момента воздействия стимула до возвращения к состоянию покоя занимает всего лишь несколько миллисекунд или даже меньше (рис. 18-17). [c.85]
Во многих нейронах, хотя и не во всех (важное исключение составляют миелинизированные аксоны млекопитающих), возвращение к состоянию покоя ускоряется благодаря потенциал-зависимым калиевым каналам в плазматической мембране. Эти каналы, подобно натриевым, открываются в ответ на деполяризацию мембраны, но происходит это отноо1тельно медленно. Повышение проницаемости мембраны для ионов К как раз в то время, когда натриевые каналы инактивируются, позволяет быстро сдвинуть мембранный потенциал до равновесного потенциала К и тем самым вернуть мембрану в состояние покоя (рис. 18-18). В результате реполяризации мембраны калиевые каналы вновь закрываются, а натриевые могут теперь выйти из состояния инактивации. Таким образом, клеточная мембрана меньше чем за одну миллисекунду вновь приобретает аюсобность отвечать на деполяризующий стимул [c.85]
18-18). В результате реполяризации мембраны калиевые каналы вновь закрываются, а натриевые могут теперь выйти из состояния инактивации. Таким образом, клеточная мембрана меньше чем за одну миллисекунду вновь приобретает аюсобность отвечать на деполяризующий стимул [c.85]
Одним из наиболее изученных примеров могут служить клетки сетчатки позвоночных. В фоторецепторе в ответ на световое воздействие возникают медленные гиперполяризующие потенциалы, обусловленные снижением натриевой проницаемости и сдвигом мембранного потенциала в сторону равновесного потенциала для К+. В нормальных условиях в фоторецепторах сетчатки генерируются только потенциалы такого рода (рис. 8.11). Эти медленные потенциалы обеспечивают передачу информации о световом воздействии — возможно, путем торможения секреции медиатора из окончаний фоторецепторов на других нейронах сетчатки. Поскольку фоторецепторы представляют собой клетки с очень короткими отростками, потенциалы в них могут достаточно эффективно распространяться электротоническим путем, и импульсов в этих клетках нет. Сигналы с фоторецепторов передаются на биполярные и горизонтальные клетки в этих нейронах тоже возникают исключительно градуальные потенциалы. Роль всех этих клеток в обработке зрительной информации мы рассмотрим в главе 17. [c.195]
Сигналы с фоторецепторов передаются на биполярные и горизонтальные клетки в этих нейронах тоже возникают исключительно градуальные потенциалы. Роль всех этих клеток в обработке зрительной информации мы рассмотрим в главе 17. [c.195]
Ионы, создающие трансмембранный ток, можно идентифицировать, изменяя концентрацию определенных ионов в окружающей клетку среде и наблюдая, к чему это приводит. Например, натриевый ток через мембранные каналы будет зависеть от внеклеточной концентрации На и исчезнет, когда эта концентрация достигнет определенного уровня, при котором равновесный натриевый потенциал, будет равен мембранному потенциалу, V. Токи через каналы с воротами и через каналы утечки тоже будут изменяться при изменении трансмембранных концентрационных градиентов, н поэтому их можно исследовать таким же способом. Потенцнал-зависимые каналы могут быть обнаружены по внезапному изменению проницаемости мембраны для определенного иона при резком изменении V. [c. 80]
80]
Передача электрических сигналов нервной клеткой основана на изменении мембранного потенциала в результате прохождения относительно небольшого числа ионов через мембранные каналы. Эти ионы перемещаются за счет энергии, большой запас которой создаежя благодаря работе Ыа К -АТРазного насоса, поддерживающего более низкую концентрацию N0 и более высокую концентрацию К внутри клетки по сравнению с наружной средой. В покоящемся нейроне каналы избирательной утечки К делают мембрану более проницаемой для калия, чем для других ионов, и поэтому мембранный потенциал покоя близок к равновесному потенциалу К, составляющему примерно — 70 мВ. Внезапная деполяризация мембраны изменяет ее проницаемость, так как при этом открываются потенциал-зависимые натриевые каналы. Но, если деполяризованное состояние поддерживается, эти каналы вскоре инактивируются. Под влиянием мембранного электрического поля отдельные каналы совершают резкий переход от одной из возможных конформаций к другой. Потенциал действия инициируется тогда, когда под влиянием короткого деполяризующего стимула открывается часть потенциал-зависимых натриевых каналов, что делает мембрану более проницаемой для Ыа и еще дальше смещает мембранный потенциал по направлению к равновесному натриевому потенциалу. В результате такой положительной обратной связи открывается еще больше натриевых каналов, и так продолжается до тех пор, пока не возникнет потенциал действия, подчиняющийся закону всё или ничего . Потенциал действия быстро исчезает вследствие инактивации натриевых каналов, а во многих нейронах также и открытия потенциал-зависимых калиевых каналов. Распространение потенциала действия (импульса) по нервному волокну зависит от кабельных свойств этого волокна. Когда при импульсе мембрана на некотором участке деполяризуется, ток, проходящий здесь через натриевые каналы, деполяризует соседние участки мембраны, где в свою очередь возникают потенциалы действия. Во многих аксонах позвоночных высокая скорость и эффективность проведения импульсов достигается благодаря изоляции поверхности аксона миелиновой оболочкой, оставляющей открытыми лишь небольшие участки возбудимой мембраны.
Потенциал действия инициируется тогда, когда под влиянием короткого деполяризующего стимула открывается часть потенциал-зависимых натриевых каналов, что делает мембрану более проницаемой для Ыа и еще дальше смещает мембранный потенциал по направлению к равновесному натриевому потенциалу. В результате такой положительной обратной связи открывается еще больше натриевых каналов, и так продолжается до тех пор, пока не возникнет потенциал действия, подчиняющийся закону всё или ничего . Потенциал действия быстро исчезает вследствие инактивации натриевых каналов, а во многих нейронах также и открытия потенциал-зависимых калиевых каналов. Распространение потенциала действия (импульса) по нервному волокну зависит от кабельных свойств этого волокна. Когда при импульсе мембрана на некотором участке деполяризуется, ток, проходящий здесь через натриевые каналы, деполяризует соседние участки мембраны, где в свою очередь возникают потенциалы действия. Во многих аксонах позвоночных высокая скорость и эффективность проведения импульсов достигается благодаря изоляции поверхности аксона миелиновой оболочкой, оставляющей открытыми лишь небольшие участки возбудимой мембраны. [c.92]
[c.92]
Судя по равновесным потенциалам на катоде должен выделяться только водород. Действительно, в нейтральном насыщенном растворе Na I при комнатной температуре потенциал водородного электрода равен —0,42 В, потенциал амальгамно-натриевого электрода с активностью натрия (равной единице) составляет около —2,7 В. Однако выделение водорода на ртути происходит с очень большим перенапряжением (рис. 149) в то время как натрий выделяется на ртути с большим эффектом деполяризации. Этот эффект обусловлен образованием прочного химического соединения натрия с ртутью типа NaHgm и растворением его в избытке ртути. Активность натрия в ртути благодаря этому резко снижается. Потенциал без тока разбавленной амальгамы натрия в насыщенном растворе хлорида натрия становится равным примерно —1,8 В. Катодное выделение натрия в ртуть при достаточно интенсивном [c.355]
Процесс возбуждения развивается вследствие зависимости проницаемости мембраны для ионов от мембранного потенциала. При достижении критической деполяризации, когда возрастает проницаемость мембраны для Ма , эти ионы устремляются внутрь и вызывают дальнейшую деполяризацию мембраны. Процесс продолжается до тех пор, пока потенциал не сместится до равновесного натриевого потенциала. В этих условиях потоки Ма наружу и внутрь сравниваются. Затем происходит увеличение проницаемости для К+ и ионы К начинают выходить из клетки по градиенту своего электрохимического потенциала. В этом процессе мембрана реполяризуется. Выход К+ прекраш ается, когда потенциал на мембране приблизится к равновесному калиевому потенциалу. [c.169]
При достижении критической деполяризации, когда возрастает проницаемость мембраны для Ма , эти ионы устремляются внутрь и вызывают дальнейшую деполяризацию мембраны. Процесс продолжается до тех пор, пока потенциал не сместится до равновесного натриевого потенциала. В этих условиях потоки Ма наружу и внутрь сравниваются. Затем происходит увеличение проницаемости для К+ и ионы К начинают выходить из клетки по градиенту своего электрохимического потенциала. В этом процессе мембрана реполяризуется. Выход К+ прекраш ается, когда потенциал на мембране приблизится к равновесному калиевому потенциалу. [c.169]
Если мембрана деполяризована так, что разность потенциалов равна напряжению натриевой батареи (срма равновесный Ма -потенциал), то Ма -ток отсутствует и наблюдается только К+-ток, возникаюш ий с обычной задержкой. При смеш ении потенциала до положительных значений, превышаюш их уровень pNa Na -тoк направлен наружу (рис. XXIII.5). [c.172]
Здесь — равновесный натриевый потенциал, т. е. нернстовский потенциал для натрия, который равен примерно -Ь40 мВ (он примерно равен овершуту). [c.91]
е. нернстовский потенциал для натрия, который равен примерно -Ь40 мВ (он примерно равен овершуту). [c.91]
Мы уже знаем о том, что поток ионов через мембрану зависит от двух факторов проницаемости для этих ионов и их электрохимического градиента. Если мы искусственно сдвигаем мембранный потенциал к равновесному потенциалу для данного иона, то электрохимический градиент снижается. Когда мембранный потенциал становится равным равновесному, ток ионов 1прекраш,ается, а при дальнейшем смещении мембранного потенциала направление этого тока меняется на противоположное. Пример подобного опыта приведен на рис. 8.7А. Представленные на данном рисунке кривые соответствуют синаптическим потенциалам, обусловленным увеличением проницаемости для натрия, при различных уровнях фиксированного потенциала. Стрелками указано направление и относительная величина натриевого тока. Поскольку знак синаптического потенциала меняется, когда мембранный потенциал превосходит равновесный, последний называют также потенциалом реверсии.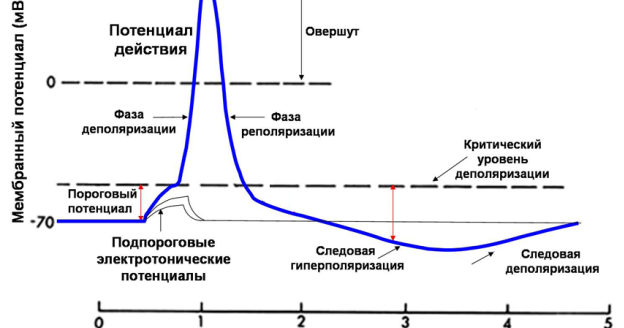 [c.186]
[c.186]
Если обратиться к данным для значений равновесных нернстов-ских потенциалов, созданных различными ионами (табл. 3.1), естественно предположить, что положительный потенциал реверсии имеет натриевую природу, поскольку именно диффузия на- [c.80]
физика электрогенез практическое занятие — Docsity
Электрогенез Мембранный потенциал – электрическая разность потенциалов на мембране живой клетки в состоянии физиологического покоя. Величина МП=-60- 90 мВ. «Наружная» поверхность клетки несет положительный заряд, цитоплазма — отрицательный. Потенциал действия Потенциалом действия (ПД) называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения. Все клетки возбудимых тканей при действии различных раздражителей достаточной силы способны переходить в состояние возбуждения. К возбудимым относятся нервная, мышечная и железистая ткани. Возбудимость – это способность клеток к быстрому ответу на раздражение, проявляющемуся через совокупность физических, физико-химических процессов и функциональных изменений. Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны. Опыт показывает, что возбужденный участок клетки становится электроотрицательным по отношению к невозбужденному, что указывает на перераспределение ионов в возбужденном участке. При возбуждении оно имеет временный характер, и после окончания возбуждения вновь восстанавливается исходный потенциал покоя. Общее изменение разности потенциалов между клеткой и средой, происходящее при пороговом и сверхпороговом возбуждении клеток, называется потенциалом действия. Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток. Современная теория возникновения потенциала действия базируется на данных, полученных методами внутриклеточного отведения потенциалов, фиксации напряжения на мембране, радиоактивных изотопов, перфузии нервных волокон, электропроводности и др. Потенциал действия приблизительно на 98% формируется за счет потоков Na+ , K+ , Cl-.
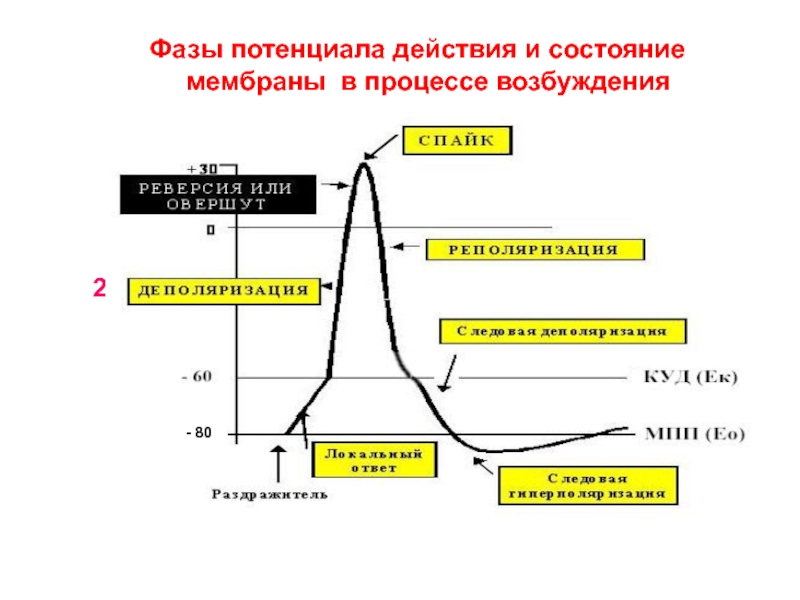
Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны. Опыт показывает, что возбужденный участок клетки становится электроотрицательным по отношению к невозбужденному, что указывает на перераспределение ионов в возбужденном участке. При возбуждении оно имеет временный характер, и после окончания возбуждения вновь восстанавливается исходный потенциал покоя. Общее изменение разности потенциалов между клеткой и средой, происходящее при пороговом и сверхпороговом возбуждении клеток, называется потенциалом действия. Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток. Современная теория возникновения потенциала действия базируется на данных, полученных методами внутриклеточного отведения потенциалов, фиксации напряжения на мембране, радиоактивных изотопов, перфузии нервных волокон, электропроводности и др. Потенциал действия приблизительно на 98% формируется за счет потоков Na+ , K+ , Cl-. ПД имеет три фазы: 1. фаза – деполяризация 2. фаза – реполяризация 3. фаза – сверхполяризация Еще в 1938 г. Коул и Кертис показали, что возбуждение связано с кратковременным увеличением электропроводности клеточной мембраны. Согласно их данным, сопротивление мембраны аксона кальмара изменяется от 1000 Ом/см2 в состоянии покоя до 25 Ом/см2 в момент возбуждения, а клетки водоросли Nitella от 100 000 до 500 Ом/см2. При этом сопротивление цитоплазмы клеток практически не изменялось. Уменьшение электрического сопротивления мембраны при возбуждении может быть объяснено только увеличением ее проницаемости для ионов, поскольку последние являются переносчиками электричества в тканях. Было показано, что возникновение потенциала действия связано с увеличением проницаемости мембраны для ионов натрия и последующим усилением диффузии этих ионов по концентрационному градиенту внутрь клетки, что приводит к изменению (уменьшению) мембранного потенциала. При этом обнаружилось, что если мембранный потенциал уменьшается до некоторой критической величины (на 10 – 30 мВ), то, независимо от того, чем вызвано это уменьшение – наложением внешнего электрического поля или же действием другого раздражителя, между проницаемостью мембраны для натрия и уменьшением ее мембранного потенциала (деполяризацией) возникает регенеративная или положительная обратная связь.
ПД имеет три фазы: 1. фаза – деполяризация 2. фаза – реполяризация 3. фаза – сверхполяризация Еще в 1938 г. Коул и Кертис показали, что возбуждение связано с кратковременным увеличением электропроводности клеточной мембраны. Согласно их данным, сопротивление мембраны аксона кальмара изменяется от 1000 Ом/см2 в состоянии покоя до 25 Ом/см2 в момент возбуждения, а клетки водоросли Nitella от 100 000 до 500 Ом/см2. При этом сопротивление цитоплазмы клеток практически не изменялось. Уменьшение электрического сопротивления мембраны при возбуждении может быть объяснено только увеличением ее проницаемости для ионов, поскольку последние являются переносчиками электричества в тканях. Было показано, что возникновение потенциала действия связано с увеличением проницаемости мембраны для ионов натрия и последующим усилением диффузии этих ионов по концентрационному градиенту внутрь клетки, что приводит к изменению (уменьшению) мембранного потенциала. При этом обнаружилось, что если мембранный потенциал уменьшается до некоторой критической величины (на 10 – 30 мВ), то, независимо от того, чем вызвано это уменьшение – наложением внешнего электрического поля или же действием другого раздражителя, между проницаемостью мембраны для натрия и уменьшением ее мембранного потенциала (деполяризацией) возникает регенеративная или положительная обратная связь. Уменьшение мембранного потенциала ниже критического уровня приводит к увеличению проницаемости мембраны для натрия, а увеличение проницаемости сопровождается усилением диффузии натрия в цитоплазму, что компенсировали друг друга и никакого изменения мембранного потенциала не могло бы происходить В конечном итоге диффузия натрия и калия по градиентам должна бы приводить к выравниванию концентраций этих ионов между наружным раствором и цитоплазмой. В действительности этого не наблюдается. В периоды покоя концентрационные градиенты калия и натрия восстанавливаются в результате работы натрий-калиевого насоса, обусловливающего перенос этих ионов против градиентов. Характерные свойства потенциала действия: 1) наличие порогового значения деполяризующего потенциала; 2) закон «все или ничего», то есть, если деполяризующий потенциал больше порогового, развивается потенциал действия,амплитуда которого не зависит от амплитуды возбуждающего импульса и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой; 3) есть период рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения; 4) в момент возбуждения резко уменьшается сопротивление мембраны (у аксона кальмара от ОД Ом * м2 -в покое до 0,0025Оа * м3 -при возбуждении).
Уменьшение мембранного потенциала ниже критического уровня приводит к увеличению проницаемости мембраны для натрия, а увеличение проницаемости сопровождается усилением диффузии натрия в цитоплазму, что компенсировали друг друга и никакого изменения мембранного потенциала не могло бы происходить В конечном итоге диффузия натрия и калия по градиентам должна бы приводить к выравниванию концентраций этих ионов между наружным раствором и цитоплазмой. В действительности этого не наблюдается. В периоды покоя концентрационные градиенты калия и натрия восстанавливаются в результате работы натрий-калиевого насоса, обусловливающего перенос этих ионов против градиентов. Характерные свойства потенциала действия: 1) наличие порогового значения деполяризующего потенциала; 2) закон «все или ничего», то есть, если деполяризующий потенциал больше порогового, развивается потенциал действия,амплитуда которого не зависит от амплитуды возбуждающего импульса и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой; 3) есть период рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения; 4) в момент возбуждения резко уменьшается сопротивление мембраны (у аксона кальмара от ОД Ом * м2 -в покое до 0,0025Оа * м3 -при возбуждении). При развитии потенциала действия происходит изменение возбудимости ткани. Это изменение протекает по фазам. Состояние исходной поляризации мембраны характерно отражает мембранный потенциал покоя, которому соответствует исходное состояние возбудимости а, следовательно, исходное состояние возбудимой клетки. Период предспайка – это период самого начала потенциала действия. Возбудимость ткани слегка повышена. Данная фаза возбудимости есть первичная экзальтация (первичная супернормальная возбудимость). Во время развития предспайка мембранный потенциал приближается к критическому уровню деполяризации и для достижения этого уровня сила раздражителя может быть меньше пороговой. В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны, и она утрачивает способность отвечать возбуждением на раздражители сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности, т.е. абсолютной невозбудимости, которая длится до конца перезарядки мембраны.
При развитии потенциала действия происходит изменение возбудимости ткани. Это изменение протекает по фазам. Состояние исходной поляризации мембраны характерно отражает мембранный потенциал покоя, которому соответствует исходное состояние возбудимости а, следовательно, исходное состояние возбудимой клетки. Период предспайка – это период самого начала потенциала действия. Возбудимость ткани слегка повышена. Данная фаза возбудимости есть первичная экзальтация (первичная супернормальная возбудимость). Во время развития предспайка мембранный потенциал приближается к критическому уровню деполяризации и для достижения этого уровня сила раздражителя может быть меньше пороговой. В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны, и она утрачивает способность отвечать возбуждением на раздражители сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности, т.е. абсолютной невозбудимости, которая длится до конца перезарядки мембраны. Абсолютная рефрактерность мембраны возникает в связи с тем, что натриевые каналы полностью открываются, а затем инактивируются. После окончания фазы перезарядки возбудимость ее постепенно восстанавливается до исходного уровня – эта фаза есть фаза относительной рефрактерности, т.е. относительной невозбудимости. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Поскольку в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых каналов. Следующему периоду соответствует повышенный уровень возбудимости: фаза вторичной экзальтации или вторичной супернормальной возбудимости. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, по сравнению с состоянием покоя исходной поляризации, то порог раздражения снижен, т.
Абсолютная рефрактерность мембраны возникает в связи с тем, что натриевые каналы полностью открываются, а затем инактивируются. После окончания фазы перезарядки возбудимость ее постепенно восстанавливается до исходного уровня – эта фаза есть фаза относительной рефрактерности, т.е. относительной невозбудимости. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Поскольку в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых каналов. Следующему периоду соответствует повышенный уровень возбудимости: фаза вторичной экзальтации или вторичной супернормальной возбудимости. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, по сравнению с состоянием покоя исходной поляризации, то порог раздражения снижен, т. е. возбудимость клетки повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. Мембранный потенциал увеличивается, то есть возникает состояние гиперполяризации мембраны. Удаляясь от критического уровня деполяризации, порог раздражения слегка повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Распространение нервного импульса вдоль аксона Потенциал действия, возникнув в одном участке нервной клетки, быстро распространяется по всей ее поверхности. Распространение потенциала действия обусловлено возникновением так называемых локальных токов, циркулирующих между возбужденным и невоз-бужденным участками клетки. В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал, а внутренняя—отрицательный. В момент возбуждения полярность мембраны изменяется на обратную: ее внешняя поверхность заряжена отрицательно по отношению к внутренней (рис.
е. возбудимость клетки повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. Мембранный потенциал увеличивается, то есть возникает состояние гиперполяризации мембраны. Удаляясь от критического уровня деполяризации, порог раздражения слегка повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Распространение нервного импульса вдоль аксона Потенциал действия, возникнув в одном участке нервной клетки, быстро распространяется по всей ее поверхности. Распространение потенциала действия обусловлено возникновением так называемых локальных токов, циркулирующих между возбужденным и невоз-бужденным участками клетки. В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал, а внутренняя—отрицательный. В момент возбуждения полярность мембраны изменяется на обратную: ее внешняя поверхность заряжена отрицательно по отношению к внутренней (рис. 2). В результате этого между возбужденным и невозбужденным участками мембраны (В и Н на рис. 2) имеется разность потенциалов. Наличие разности потенциалов приводит к появлению между этими участками электрических токов, называемых локальными токами или токами действия. На поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении. Локальный ток, как и любой электрический ток, оказывает раздражающее действие на соседние невозбужденные участки и вызывает увеличение проницаемости их мембран. Это приводит к снижению в них потенциала покоя. Когда деполяризация достигает критического значения, в этих участках возникают потенциалы действия, а в том участке, который ранее был возбужденным, в это время уже происходят восстановительное процессы реполяризации. Вновь возбужденный участок в свою очередь становится электроотрицательным и возникающий локальный ток раздражает следующий за ним участок. Этот процесс многократно повторяется и обусловливает распространение импульсов возбуждения по всей длине клетки в обоих направлениях.
2). В результате этого между возбужденным и невозбужденным участками мембраны (В и Н на рис. 2) имеется разность потенциалов. Наличие разности потенциалов приводит к появлению между этими участками электрических токов, называемых локальными токами или токами действия. На поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении. Локальный ток, как и любой электрический ток, оказывает раздражающее действие на соседние невозбужденные участки и вызывает увеличение проницаемости их мембран. Это приводит к снижению в них потенциала покоя. Когда деполяризация достигает критического значения, в этих участках возникают потенциалы действия, а в том участке, который ранее был возбужденным, в это время уже происходят восстановительное процессы реполяризации. Вновь возбужденный участок в свою очередь становится электроотрицательным и возникающий локальный ток раздражает следующий за ним участок. Этот процесс многократно повторяется и обусловливает распространение импульсов возбуждения по всей длине клетки в обоих направлениях. В нервной системе прохождение импульсов только в определенном направлении обусловлено наличием синапсов, обладающих односторонней проводимостью.
В нервной системе прохождение импульсов только в определенном направлении обусловлено наличием синапсов, обладающих односторонней проводимостью.
Тема №10600 Ответы к тестам по физиологии 11 тем (Часть 1)
Тема №10600
РАЗДЕЛ 01. ФИЗИОЛОГИЯ НЕРВОВ
Выберите один правильный ответ из предлагаемых.
001. РАЗДРАЖИТЕЛЬ ЛЮБОЙ СИЛЫ НЕ ВЫЗЫВАЕТ ВОЗБУЖДЕНИЕ В ФАЗУ
а) абсолютной рефрактерности
б) относительной рефрактерности
в) супернормальной возбудимости
г) субнормальной возбудимости
002. ЭНЕРГИЯ АТФ ИСПОЛЬЗУЕТСЯ
а) для работы ионоселективных каналов
б) для работы каналов «утечки»
в) для работы каналов «насосов»
003. ФАЗА СУПЕРНОРМАЛЬНОЙ ВОЗБУДИМОСТИ ПРИХОДИТСЯ
а) на подпороговый потенциал
б) на пиковый потенциал
в) на отрицательный следовой потенциал
г) на положительный следовой потенциал
004. ФАЗУ ДЕПОЛЯРИЗАЦИИ ОТРАЖАЕТ СЛЕДУЮЩЕЕ КОЛЕНО ПОТЕНЦИАЛА ДЕЙСТВИЯ
а) нисходящее
б) восходящее
в) оба
г) ничто из перечисленного неверно
005. МЕМБРАНА НЕРВНОЙ КЛЕТКИ В СОСТОЯНИИ ПОКОЯ
МЕМБРАНА НЕРВНОЙ КЛЕТКИ В СОСТОЯНИИ ПОКОЯ
ИМЕЕТ СЛЕДУЮЩИЙ ЗАРЯД
а) отрицательный внутри и положительный снаружи
б) положительный внутри и положительный снаружи
в) положительный внутри и отрицательный снаружи
г) отрицательный внутри и отрицательный снаружи
5
006. ПОДПОРОГОВЫЕ РАЗДРАЖИТЕЛИ ВЫЗЫВАЮТ ВОЗБУЖДЕНИЕ В СЛЕДУЮЩУЮ ФАЗУ ВОЗБУДИМОСТИ
а) абсолютная рефрактерность
б) относительная рефрактерность
в) экзальтация
г) субнормальный период
007. ГИПЕРПОЛЯРИЗАЦИЮ КЛЕТКИ МОЖНО ВЫЗВАТЬ, ЕСЛИ
а) приложить к ее поверхности катод
б) в наружной среде увеличить содержание ионов К
в) приложить к ее поверхности анод
г) возбудить клетку
008. ЗАРЯД ВОЗБУЖДЁННОЙ КЛЕТКИ СНАРУЖИ ПО ОТНОШЕНИЮ ЗАРЯД ВОЗБУЖДЕННОЙ КЛЕТКИ СНАРУЖИ ПО
ОТНОШЕНИЮ К НАРУЖНОЙ ПОВЕРХНОСТИ СОСЕДНИХ
НЕВОЗБУЖДЕННЫХ КЛЕТОК
а) нейтральный
б) положительный
в) отрицательный
009. ВОЗБУДИМОСТЬ — ЭТО
а) способность возбудимых тканей проводить возбуждение вдоль мембраны;
б) способность железистой ткани выделять секрет в ответ на раздражение;
в) способность возбудимых тканей генерировать электрический потенциал в ответ на раздражение;
г) процесс генерации электрического потенциала в ответ на раздражение.
010. ВОЗБУДИМОЙ ТКАНЬЮ ЯВЛЯЕТСЯ
а) нервная ткань
б) костная ткань
в) фиброзная ткань
г) хрящевая ткань
011. КРИТИЧЕСКИЙ УРОВЕНЬ ДЕПОЛЯРИЗАЦИИ — ЭТО
6
а) уровень деполяризации, превышение которого ведет к спаду возбуждения
б) уровень деполяризации, превышение которого ведет к возникновению потенциала покоя
в) уровень деполяризации, превышение которого ведет к возникновению потенциала действия
012. АМПЛИТУДА ПОТЕНЦИАЛА ДЕЙСТВИЯ И КРИТИЧЕСКИЙ
УРОВЕНЬ ДЕПОЛЯРИЗАЦИИ (КУД) ПРИ МЕДЛЕННОМ
НАРАСТАНИИ РАЗДРАЖАЮЩЕГО ТОКА ПО СРАВНЕНИЮ С
АНАЛОГИЧНЫМИ ПОКАЗАТЕЛЯМИ ПРИ БЫСТРОМ ЕГО
УВЕЛИЧЕНИИ
а) не изменяются
б) КУД повышается, увеличивается амплитуда
в) КУД снижается, увеличивается амплитуда
г) КУД повышается, уменьшается амплитуда
013. ВОЗБУЖДЕНИЕ В НЕРВНОЙ КЛЕТКЕ СОПРОВОЖДАЕТСЯ
а) сокращением
б) распространением электрического импульса
в) секрецией
014. БРАНШИ ПИНЦЕТА ГАЛЬВАНИ СДЕЛАНЫ ИЗ СЛЕДУЮЩИХ МЕТАЛЛОВ
а) никель и свинец
б) медь и свинец
в) свинец и железо
г) медь и цинк
015. ПОТЕНЦИАЛ ПОКОЯ — ЭТО
ПОТЕНЦИАЛ ПОКОЯ — ЭТО
а) разность потенциалов между поврежденной и неповрежденной поверхностями мышцы
б) разность потенциалов между наружной и внутренней поверхностями мембраны покоящейся клетки
в) разность потенциалов между наружной и внутренней поверхностями мембраны при возбуждении клетки
г) разность потенциалов между возбужденными и невозбужденными
участками мышцы
7
016. РЕОБАЗА — ЭТО
а) минимальная сила тока, которая должна действовать определенное
время для возникновения возбуждения
б) минимальное время, в течение которого должен действовать ток пороговой силы, чтобы вызвать возбуждение
в) величина тока меньше порога раздражения, не способная вызвать
возбуждение
017. ХРОНАКСИЯ — ЭТО
а) минимальное время действия раздражителя силой в две реобазы,
необходимое для возникновения возбуждения
б) минимальное время действия тока величиной в одну реобазу, необходимое для возбуждения
в) минимальная сила тока, которая должна действовать определенное
время для возникновения возбуждения
018. ПОЛЕЗНОЕ ВРЕМЯ — ЭТО
ПОЛЕЗНОЕ ВРЕМЯ — ЭТО
а) минимальное время действия раздражителя силой в две реобазы, достаточное для возникновения возбуждения
б) минимальное время действия раздражителя пороговой силы, достаточной для возникновения возбуждения
в) это время действия раздражителя сверх того, которое необходимо
для формирования возбуждения
019. ВОЗБУЖДЕНИЕ ПО ТОНКИМ ВЕГЕТАТИВНЫМ ПРЕГАНГЛИОНАРНЫМ ВОЛОКНАМ (ТИП В) ПРОХОДИТ СО
СЛЕДУЮЩЕЙ СКОРОСТЬЮ (М/С)
а) 300 м/c
б) 200 м/c
в) 140-150 м/c
г) 3-18 м/c
020. ВОЗБУЖДЕНИЕ ПО ОЧЕНЬ ТОНКИМ (0,5-1,0 МКМ)
СИМПАТИЧЕСКИМ ПОСТГАНГЛИОНАРНЫМ НЕРВНЫМ
ВОЛОКНАМ (ТИП С) ПРОХОДИТ СО СЛЕДУЮЩЕЙ СКОРОСТЬЮ
(М/С)
а) 300 м/с
б) 100 м/с
в) 0,5-3 м/с
8
г) 0,05 м/с
021. ЛАБИЛЬНОСТЬ ДВИГАТЕЛЬНОГО НЕРВА ТЕПЛОКРОВНОГО
ЖИВОТНОГО СОСТАВЛЯЕТ
а) 10000
б) 0-500
в) 1000
г) 100
022. НАИБОЛЬШУЮ ЛАБИЛЬНОСТЬ ИМЕЕТ СЛЕДУЮЩЕЕ
ВОЛОКНО
а) типа А
б) типа В
в) типа С
г) одинаково
023. ЛАБИЛЬНОСТЬ НЕРВНОГО ВОЛОКНА ТИПА В СОСТАВЛЯЕТ
ЛАБИЛЬНОСТЬ НЕРВНОГО ВОЛОКНА ТИПА В СОСТАВЛЯЕТ
а) 2000
б) 300
в) 4
г) менее 1
024. МИЕЛИНОВАЯ ОБОЛОЧКА ОТСУТСТВУЕТ В СЛЕДУЮЩЕМ
ТИПЕ НЕРВНЫХ ВОЛОКОН
а) С
б) В
в) А
025. В ОРГАНИЗМЕ ЧЕЛОВЕКА
а) одинаковое количество афферентных и эфферентных нервных волокон
б) больше афферентных нервных волокон
в) больше эфферентных нервных волокон
026. БОЛЕЕ КОРОТКИЙ РЕФРАКТЕРНЫЙ ПЕРИОД ИМЕЕТ
СЛЕДУЮЩИЙ ВИД НЕРВНЫХ ВОЛОКОН
а) толстые волокна
б) тонкие волокна
9
в) одинаково
027. К ВОЛОКНАМ ТИПА С ОТНОСЯТСЯ СЛЕДУЮЩИЕ ИЗ
ПЕРЕЧИСЛЕННЫХ НЕРВНЫХ ВОЛОКОН
а) вегетативные постганглионарные
б) вегетативные преганглионарные
в) от мышечных рецепторов, моторные для скелетных мышц
028. К ВОЛОКНАМ ТИПА B ОТНОСЯТСЯ СЛЕДУЮЩИЕ ИЗ
ПЕРЕЧИСЛЕННЫХ НЕРВНЫХ ВОЛОКОН
а) вегетативные постганглионарные
б) вегетативные преганглионарные
в) от мышечных рецепторов, моторные для скелетных мышц
029. К ВОЛОКНАМ ТИПА A ОТНОСЯТСЯ СЛЕДУЮЩИЕ ИЗ
ПЕРЕЧИСЛЕННЫХ НЕРВНЫХ ВОЛОКОН
а) вегетативные постганглионарные
б) вегетативные преганглионарные
в) от мышечных рецепторов, моторные для скелетных мышц
030. ВОЗБУЖДЕНИЕ ПО ТОЛСТЫМ (12-20 МКМ) НЕРВНЫМ
ВОЗБУЖДЕНИЕ ПО ТОЛСТЫМ (12-20 МКМ) НЕРВНЫМ
ВОЛОКНАМ, ИДУЩИМ К СКЕЛЕТНЫМ МЫШЦАМ,
ПРОВОДИТСЯ СО СЛЕДУЮЩЕЙ СКОРОСТЬЮ (М/С)
а) 160 м/с
б) 70-120 м/с
в) 40-70 м/с
г) 0,5 м/с
031. ПРОВОДИМОСТЬ НЕРВНЫХ ИМПУЛЬСОВ В ОБЛАСТИ
АНОДА ПРИ ЗАМЫКАНИИ ЦЕПИ ПОСТОЯННОГО ТОКА
а) не изменяется
б) понижается
в) повышается
032. ВОЗБУДИМОСТЬ НЕРВА В ОБЛАСТИ КАТОДА ПРИ
ЗАМЫКАНИИ ЦЕПИ ПОСТОЯННОГО ТОКА
а) не изменяется
б) понижается
в) повышается
10
033. ПРИ СРЕДНЕЙ СИЛЕ ТОКА, ПРИЛОЖЕННОГО К СЕДАЛИЩНОМУ НЕРВУ ЛЯГУШКИ, НЕЗАВИСИМО ОТ ЕГО
НАПРАВЛЕНИЯ, СОКРАЩЕНИЕ ИКРОНОЖНОЙ МЫШЦЫ БУДЕТ
ПРОИСХОДИТЬ
а) сокращение будет как при замыкании, так и при размыкании цепи
постоянного тока
б) будет только при замыкании цепи постоянного тока
в) будет только при размыкании цепи постоянного тока
034. ОБМЕН ВЕЩЕСТВ В ПАРАБИОТИЧЕСКОМ УЧАСТКЕ НЕРВА
а) повышается
б) понижается
в) не изменяется
035. КАТОДИЧЕСКАЯ ДЕПРЕССИЯ Б. Ф. ВЕРИГО — ЭТО
Ф. ВЕРИГО — ЭТО
а) снижение возбудимости под катодом при длительном прохождении
постоянного тока через нерв
б) повышение возбудимости под катодом при длительном прохождении постоянного тока через нерв
в) гибель нерва под действием постоянного тока
036. ПОД КАТОДОМ ПРИ ЗАМЫКАНИИ ПОСТОЯННОГО ТОКА
ПРОИСХОДЯТ СЛЕДУЮЩИЕ ПРОЦЕССЫ
а) деполяризация и повышение возбудимости
б) деполяризация и снижение возбудимости
в) гиперполяризация и повышение возбудимости
г) гиперполяризация и снижение возбудимости
037. МИНИМАЛЬНЫЙ ГРАДИЕНТ — ЭТО
а) максимальная скорость нарастания раздражителя, при которой ткань
способна отвечать возбуждением
б) минимальная сила раздражителя, способная вызывать возбуждение
в) минимальная скорость нарастания раздражителя, при которой ткань
еще способна отвечать возбуждением
г) минимальное время, в течение которого должен действовать раздражитель силой, равной двум реобазам
038. ФИЗИОЛОГИЧЕСКИЙ ЭЛЕКТРОТОН — ЭТО
11
а) повышение возбудимости под катодом и понижение под анодом при
замыкании цепи постоянного тока
б) повышение возбудимости под катодом и понижение под анодом при
размыкании цепи постоянного тока
в) понижение возбудимости под катодом и повышение под анодом при
замыкании цепи постоянного тока
г) понижение возбудимости под катодом и повышение под анодом при
размыкании цепи постоянного тока
039. ОПРЕДЕЛЕНИЕ И ПРИЧИНА КАТОДИЧЕСКОЙ ДЕПРЕССИИ
ОПРЕДЕЛЕНИЕ И ПРИЧИНА КАТОДИЧЕСКОЙ ДЕПРЕССИИ
а) резкое повышение возбудимости под катодом вследствие активации
натриевых каналов
б) резкое повышение возбудимости под катодом вследствие инактивации натриевых каналов
в) резкое понижение возбудимости под катодом вследствие активации
натриевых каналов
г) резкое понижение возбудимости под катодом вследствие инактивации натриевых каналов
040. НАРУЖНАЯ ПОВЕРХНОСТЬ МЕМБРАНЫ КЛЕТКИ В ПОКОЕ
ЗАРЯЖЕНА
а) электроположительно
б) электроотрицательно
в) не заряжена
041. ВНУТРЕННЯЯ ПОВЕРХНОСТЬ МЕМБРАНЫ КЛЕТКИ В
ПОКОЕ ЗАРЯЖЕНА
а) электроположительно
б) электроотрицательно
в) не заряжена
042. НАРУЖНАЯ ПОВЕРХНОСТЬ МЕМБРАНЫ КЛЕТКИ НА ПИКЕ
ВОЗБУЖДЕНИЯ ЗАРЯЖЕНА
а) электроположительно
б) электроотрицательно
в) не заряжена
043. ВНУТРЕННЯЯ ПОВЕРХНОСТЬ МЕМБРАНЫ КЛЕТКИ НА
ПИКЕ ВОЗБУЖДЕНИЯ ЗАРЯЖЕНА
12
а) электроположительно
б) электроотрицательно
в) не заряжена
044. РОЛЬ НАТРИЙ-КАЛИЕВОГО НАСОСА ЗАКЛЮЧАЕТСЯ В
РОЛЬ НАТРИЙ-КАЛИЕВОГО НАСОСА ЗАКЛЮЧАЕТСЯ В
СЛЕДУЮЩЕМ
а) откачивает ионы Na из клетки, нагнетает ионы K внутрь
б) откачивает ионы K из клетки, нагнетает ионы Na внутрь
в) осуществляет пассивный транспорт Na
г) осуществляет пассивный транспорт K
045. ПОТЕНЦИАЛ ДЕЙСТВИЯ — ЭТО
а) разность потенциалов между поврежденной и неповрежденной поверхностями мышцы
б) разность потенциалов между наружной и внутренней поверхностями мембраны покоящейся клетки
в) быстрые колебания разности потенциалов между наружной и внутренней поверхностями мембраны при возбуждении клетки
г) разность потенциалов между возбужденными и невозбужденными
участками мышцы
046. ИЗМЕНЕНИЕ ЗАРЯДА МЕМБРАНЫ И ВОЗБУДИМОСТЬ ВО
ВРЕМЯ БЫСТРОЙ ДЕПОЛЯРИЗАЦИИ ТАКОВО
а) наружный участок мембраны заряжается отрицательно, возбудимость повышается
б) наружный участок мембраны заряжается отрицательно, возбудимость снижается до нуля
в) наружный участок мембраны заряжается положительно, возбудимость повышается
г) наружный участок мембраны заряжается положительно, возбудимость снижается до нуля
047. ПОД АНОДОМ ПРИ ЗАМЫКАНИИ ПОСТОЯННОГО ТОКА
ПОД АНОДОМ ПРИ ЗАМЫКАНИИ ПОСТОЯННОГО ТОКА
ПРОИСХОДЯТ СЛЕДУЮЩИЕ ПРОЦЕССЫ
а) деполяризация и повышение возбудимости
б) деполяризация и снижение возбудимости
в) гиперполяризация и повышение возбудимости
г) гиперполяризация и снижение возбудимости
13
048. ВОЗБУЖДЕНИЕ В ФАЗУ ОТНОСИТЕЛЬНОЙ РЕФРАКТЕРНОСТИ ВОЗМОЖНО В ОТВЕТ НА СЛЕДУЮЩУЮ СИЛУ
РАЗДРАЖИТЕЛЯ
а) пороговую
б) надпороговую
в) подпороговую
049. ВОЗБУЖДЕНИЕ В ФАЗУ ЭКЗАЛЬТАЦИИ ВОЗМОЖНО В
ОТВЕТ НА СЛЕДУЮЩУЮ СИЛУ РАЗДРАЖИТЕЛЯ
а) пороговую
б) надпороговую
в) подпороговую
050. ЗАКОН «ВСЕ ИЛИ НИЧЕГО» ГЛАСИТ
а) независимость амплитуды потенциала действия от силы раздражителя
б) прямо пропорциональную зависимость амплитуды потенциала действия от силы раздражителя
в) обратно пропорциональную зависимость амплитуды потенциала
действия от силы раздражителя
051. ЗАКОН «СИЛЫ-ВРЕМЕНИ» ГЛАСИТ
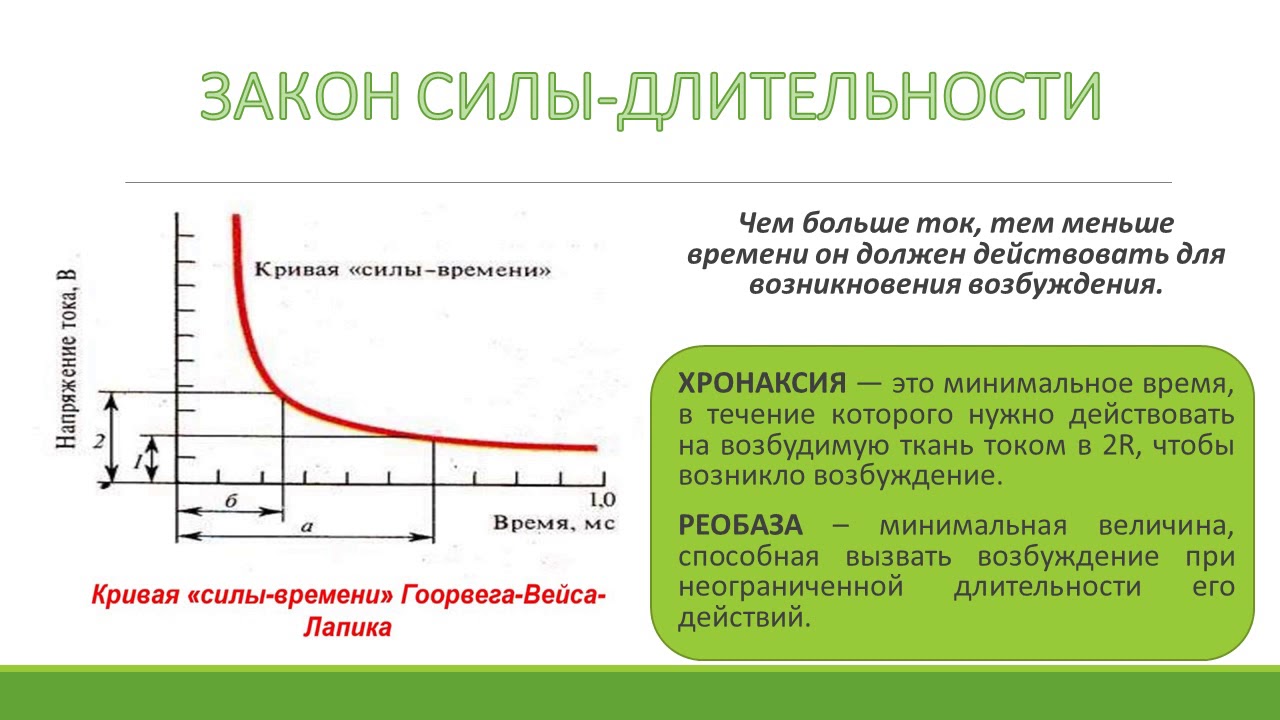
а) чем меньше времени действует на ткань раздражитель, тем больше
должна быть его сила для возбуждения
б) чем меньше времени действует на ткань раздражитель, тем меньше
должна быть его сила для возбуждения
в) пороговая сила раздражителя не зависит от времени его действия
052. ВЛИЯНИЕ КОНЦЕНТРАЦИИ ИОНОВ К ВО ВНЕШНЕЙ СРЕДЕ
ВЛИЯНИЕ КОНЦЕНТРАЦИИ ИОНОВ К ВО ВНЕШНЕЙ СРЕДЕ
НА ВЕЛИЧИНУ МЕМБРАННОГО ПОТЕНЦИАЛА ПОКОЯ (МПП)
СЛЕДУЮЩЕЕ
а) чем выше концентрация ионов K, тем меньше величина МПП
б) чем выше концентрация ионов K, тем больше величина МПП
в) чем ниже концентрация ионов K, тем меньше величина МПП
г) величина МПП не зависит от концентрации ионов K
053. ТОЛЩИНА НЕРВНЫХ ВОЛОКОН НА СКОРОСТЬ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ОКАЗЫВАЕТ СЛЕДУЮЩЕЕ
ВЛИЯНИЕ
14
а) не влияет
б) чем тоньше волокно, тем выше скорость проведения возбуждения
в) чем толще волокно, тем выше скорость проведения возбуждения
054. НА РАЗДРАЖИТЕЛЬ, СИЛА КОТОРОГО БОЛЬШЕ ПОЛОВИНЫ ПОРОГА, НО МЕНЬШЕ ПОРОГА, ВОЗНИКАЕТ
СЛЕДУЮЩИЙ ЭЛЕКТРИЧЕСКИЙ ОТВЕТ
а) электротонический потенциал
б) локальный ответ
в) потенциал действия
г) следовые потенциалы
055. ПОРОГ ДЕПОЛЯРИЗАЦИИ ЗАВИСИТ ОТ СЛЕДУЮЩИХ
ФАКТОРОВ
а) от силы раздражителя
б) от разности значений потенциала покоя и пика деполяризации
в) от разности значений потенциала покоя и критического уровня деполяризации
г) от величины овершута
056. ВОЗБУДИМОСТЬ НАХОДИТСЯ В СЛЕДУЮЩЕЙ ЗАВИСИМОСТИ ОТ ПОРОГА ДЕПОЛЯРИЗАЦИИ
ВОЗБУДИМОСТЬ НАХОДИТСЯ В СЛЕДУЮЩЕЙ ЗАВИСИМОСТИ ОТ ПОРОГА ДЕПОЛЯРИЗАЦИИ
а) не зависит
б) чем больше порог деполяризации, тем выше возбудимость
в) чем больше порог деполяризации, тем ниже возбудимость
057. ОТВЕТ НЕРВА НА РЕДКИЕ И ЧАСТЫЕ РАЗДРАЖЕНИЯ В
УРАВНИТЕЛЬНУЮ ФАЗУ ПАРАБИОЗА ИЗМЕНЯЕТСЯ ПО
СЛЕДУЮЩЕМУ ПРИНЦИПУ
а) на редкие раздражители ответ больше, чем на частые
б) на редкие раздражители ответ меньше, чем на частые
в) на редкие и частые раздражители ответ одинаковый
г) нерв не отвечает на любые частоты раздражителя
058. ОТВЕТ НЕРВА НА РЕДКИЕ И ЧАСТЫЕ РАЗДРАЖЕНИЯ В
ПАРАДОКСАЛЬНУЮ ФАЗУ ПАРАБИОЗА ИЗМЕНЯЕТСЯ ПО
СЛЕДУЮЩЕМУ ПРИНЦИПУ
а) на редкие раздражители ответ больше, чем на частые
б) на редкие раздражители ответ меньше, чем на частые
15
в) на редкие и частые раздражители ответ одинаковый
г) нерв не отвечает на любые частоты раздражителя
059. ОТВЕТ НЕРВА НА РЕДКИЕ И ЧАСТЫЕ РАЗДРАЖЕНИЯ В
ТОРМОЗНУЮ ФАЗУ ПАРАБИОЗА ИЗМЕНИТСЯ ПО
СЛЕДУЮЩЕМУ ПРИНЦИПУ
а) на редкие раздражители ответ больше, чем на частые
б) на редкие раздражители ответ меньше, чем на частые
в) на редкие и частые раздражители ответ одинаковый
г) нерв не отвечает на любые частоты раздражителя
060. ПОВТОРНОЕ ВОЗБУЖДЕНИЕ ПРИ ЕГО ДЕЙСТВИИ В
ПОВТОРНОЕ ВОЗБУЖДЕНИЕ ПРИ ЕГО ДЕЙСТВИИ В
СУБНОРМАЛЬНЫЙ ПЕРИОД ВОЗБУДИМОСТИ ВОЗМОЖНО НА
СЛЕДУЮЩУЮ СИЛУ РАЗДРАЖИТЕЛЯ
а) ответ не возникает на любую силу раздражителя
б) на пороговую силу раздражителя
в) на подпороговую силу раздражителя
г) на надпороговую силу раздражителя
061. ПОВТОРНОЕ ВОЗБУЖДЕНИЕ ПРИ ЕГО ДЕЙСТВИИ В ФАЗУ
АБСОЛЮТНОЙ РЕФРАКТЕРНОСТИ ВОЗМОЖНО НА
СЛЕДУЮЩУЮ СИЛУ РАЗДРАЖИТЕЛЯ
а) ответ не возникает на любую силу раздражителя
б) на пороговую силу раздражителя
в) на подпороговую силу раздражителя
г) на надпороговую силу раздражителя
062. НЕПОСРЕДСТВЕННОЙ ПРИЧИНОЙ СУЩЕСТВОВАНИЯ
ПОТЕНЦИАЛА ПОКОЯ ЯВЛЯЕТСЯ
а) диффузия ионов K в клетку
б) диффузия ионов K из клетки
в) диффузия ионов Na в клетку
г) диффузия ионов Na из клетки
063. ВОСХОДЯЩУЮ ЧАСТЬ ПИКА ПОТЕНЦИАЛА ДЕЙСТВИЯ
ОБУСЛАВЛИВАЮТ СЛЕДУЮЩИЕ ИОНЫ, ДВИЖУЩИЕСЯ В
ОПРЕДЕЛЕННОМ НАПРАВЛЕНИИ
а) диффузия ионов K в клетку
б) диффузия ионов K из клетки
16
в) диффузия ионов Na в клетку
г) диффузия ионов Na из клетки
064. ВЕЛИЧИНА ПОТЕНЦИАЛА ПОКОЯ НЕРВНОЙ КЛЕТКИ
ВЕЛИЧИНА ПОТЕНЦИАЛА ПОКОЯ НЕРВНОЙ КЛЕТКИ
РАВНА
а) —50 мВ
б) —70 мВ
в) —90 мВ
г) +30 мВ
065. ДВИЖУЩЕЙ СИЛОЙ, ОБЕСПЕЧИВАЮЩЕЙ ВЫХОД ИОНОВ
K ИЗ КЛЕТКИ ВО ВРЕМЯ ПОКОЯ ЯВЛЯЕТСЯ
а) диффузия
б) фильтрация
в) осмос
г) активный транспорт
066. ЛАБИЛЬНОСТЬ ТКАНИ ОТ ПРОДОЛЖИТЕЛЬНОСТИ ЕЕ
РЕФРАКТЕРНОЙ ФАЗЫ НАХОДИТСЯ В СЛЕДУЮЩЕЙ
ЗАВИСИМОСТИ
а) не зависит
б) чем длиннее рефрактерный период, тем выше лабильность
в) чем длиннее рефрактерный период, тем ниже лабильность
067. ДЛЯ ВОЗБУДИМОЙ ТКАНИ ПРИ МЕДЛЕННО НАРАСТАЮЩЕЙ СИЛЕ РАЗДРАЖИТЕЛЯ ХАРАКТЕРНО
СЛЕДУЮЩЕЕ ЯВЛЕНИЕ
а) адаптация
б) аккомодация
в) утомление
г) парабиоз
068. В ОСНОВЕ АККОМОДАЦИИ ЛЕЖИТ
а) инактивация Na-каналов
б) активация Na-каналов
в) блокада Na-каналов
17
РАЗДЕЛ 02. ФИЗИОЛОГИЯ МЫШЦ
Выберите один правильный ответ из предлагаемых.
001. ТОЛСТЫЕ ПРОТОФИБРИЛЛЫ СОСТОЯТ ИЗ
а) из актина
б) из миозина
в) из тропонина
г) из тропомиозина
002. СВЕТЛЫЕ ДИСКИ ОБРАЗУЮТ СКОПЛЕНИЯ СЛЕДУЮЩИХ
СВЕТЛЫЕ ДИСКИ ОБРАЗУЮТ СКОПЛЕНИЯ СЛЕДУЮЩИХ
ПРОТОФИБРИЛЛ
а) актиновых
б) миозиновых
в) актиновых и миозиновых
003. ПОПЕРЕЧНЫЕ МОСТИКИ — ЭТО
а) тропонин-тропомиозиновые комплексы
б) выросты белковых молекул актина
в) выросты белковых молекул миозина
г) впячивания поверхностей мембраны
004. УЧАСТИЕ В СОПРЯЖЕНИИ ЭЛЕКТРИЧЕСКИХ И МЕХАНИЧЕСКИХ ПРОЦЕССОВ В МЫШЦЕ ПРИНИМАЕТ
СЛЕДУЮЩИЙ ИЗ ПЕРЕЧИСЛЕННЫХ ИОНОВ
а) ион Са
б) ион Сl
в) ион Na
г) ион K
005. ЗУБЧАТЫЙ ТЕТАНУС МОЖНО ПОЛУЧИТЬ СЛЕДУЮЩИМ
ОБРАЗОМ
а) если наносить раздражения в латентный период сокращения
б) если наносить раздражение в фазу укорочения
в) если наносить раздражение в фазу расслабления
г) если наносить раздражение после окончания полного цикла одиночного сокращения
18
006. ТЕТАНУС — ЭТО
а) сокращение мышцы в ответ на раздражение одиночным импульсом
б) снижение работоспособности после длительной работы
в) ухудшение физиологических свойств под влиянием повреждающего
(альтерирующего) фактора
г) длительное суммированное сокращение мышцы при частом ритмическом раздражении
007. ЧТОБЫ ПОЛУЧИТЬ ОПТИМАЛЬНОЕ СОКРАЩЕНИЕ МЫШЦ,
ЧТОБЫ ПОЛУЧИТЬ ОПТИМАЛЬНОЕ СОКРАЩЕНИЕ МЫШЦ,
НАДО НАНЕСТИ РАЗДРАЖЕНИЕ В СЛЕДУЮЩУЮ ФАЗУ
ВОЗБУЖДЕНИЯ
а) абсолютной рефрактерности
б) относительной рефрактерности
в) экзальтации
г) субнормальной возбудимости
008. УКОРОЧЕНИЕ МЫШЦЫ ПРОИСХОДИТ ЗА СЧЕТ
а) укорочения миозиновых нитей
б) укорочения актиновых нитей
в) ослабления сухожилий
г) скольжения актиновых нитей вдоль миозиновых
009. МЕДИАТОРОМ В НЕРВНО-МЫШЕЧНОМ СИНАПСЕ
ЯВЛЯЕТСЯ
а) норадреналин
б) дофамин
в) ацетилхолин
г) глицин
010. СКЕЛЕТНАЯ МЫШЕЧНАЯ И НЕРВНАЯ ТКАНИ ПО ПОРОГУ
РАЗДРАЖЕНИЯ И ВОЗБУДИМОСТИ ИМЕЮТ СЛЕДУЮЩИЕ
ОТЛИЧИЯ ДРУГ ОТ ДРУГА
а) у скелетной мышечной ткани порог раздражения и возбудимость
выше, чем у нервной ткани
б) у скелетной мышечной ткани порог раздражения выше, а возбудимость ниже, чем у нервной ткани
в) у скелетной мышечной ткани порог раздражения ниже, а возбудимость выше, чем у нервной ткани
19
г) у скелетной мышечной ткани порог раздражения и возбудимость
ниже, чем у нервной ткани
011. АЦЕТИЛХОЛИН ИНАКТИВИРУЕТ СЛЕДУЮЩИЙ ФЕРМЕНТ
АЦЕТИЛХОЛИН ИНАКТИВИРУЕТ СЛЕДУЮЩИЙ ФЕРМЕНТ
а) холинэстераза
б) моноаминоксидаза
в) катехол-о-метилтрансфераза
012. СИНАПС — ЭТО
а) органелла, содержащая медиатор
б) часть нервной клетки, от которой берет начало аксон
в) окончание нервной клетки
г) структура, обеспечивающая передачу возбуждения с нервной клетки
на другую клетку
013. ЗНАЧЕНИЕ ПОТЕНЦИАЛА ПОКОЯ СКЕЛЕТНОЙ МЫШЕЧНОЙ КЛЕТКИ ТАКОВО
а) +30 мВ
б) —70 мВ
в) —50 мВ
г) —90 мВ
014. ИОНЫ Са, УЧАСТВУЮЩИЕ В СОКРАЩЕНИИ СКЕЛЕТНЫХ
МЫШЦ, ПОСТУПАЮТ В САРКОПЛАЗМУ ИЗ
а) из межклеточного пространства
б) из саркоплазматического ретикулума
в) из везикул нервного окончания
г) из синаптической щели
015. МЫШЦЫ ГЛАВНЫМ ОБРАЗОМ ОБЕСПЕЧИВАЮТСЯ
ЭНЕРГИЕЙ ЗА СЧЕТ СЛЕДУЮЩИХ ПРОЦЕССОВ
а) за счет бескислородного окисления глюкозы
б) за счет кислородного окисления глюкозы и жирных кислот
в) за счет кислородного окисления белков
016. В ОСВОБОЖДЕНИИ МЕДИАТОРА ИЗ ПРЕСИНАПТИЧЕСКОГО ОКОНЧАНИЯ УЧАСТВУЕТ СЛЕДУЮЩИЙ ИЗ
ПЕРЕЧИСЛЕННЫХ ИОНОВ
а) ион Са
20
б) ион К
в) ион Na
г) ион Mg
017. ПОТЕНЦИАЛ ДЕЙСТВИЯ В МЫШЕЧНЫХ КЛЕТКАХ ДЛИТСЯ
ПОТЕНЦИАЛ ДЕЙСТВИЯ В МЫШЕЧНЫХ КЛЕТКАХ ДЛИТСЯ
а) 5 с
б) 50-100 мс
в) 1-5 мс
г) до 0,1 мс
018. СКЕЛЕТНЫЕ МЫШЦЫ У ТРЕНИРОВАННОГО ЧЕЛОВЕКА
СОСТАВЛЯЮТ СЛЕДУЮЩИЙ % МАССЫ ТЕЛА
а) 75 %
б) 60 %
в) 35-40 %
г) 50 %
019. ОСНОВНЫЕ ФУНКЦИИ, ВЫПОЛНЯЕМЫЕ СКЕЛЕТНЫМИ
МЫШЦАМИ ЧЕЛОВЕКА
а) опорно-двигательную и сенсорную
б) содействие лимфотоку и доставке венозной крови к сердцу
в) образование тепла
г) депонирующую (гликоген и др.)
д) все перечисленное верно
020. СКЕЛЕТНЫЕ МЫШЦЫ У НЕТРЕНИРОВАННОГО ЧЕЛОВЕКА
СОСТАВЛЯЮТ СЛЕДУЮЩИЙ % МАССЫ ТЕЛА
а) 75 %
б) 60 %
в) 35-40 %
г) 50 %
021. ПРИ ИЗОТОНИЧЕСКОМ СОКРАЩЕНИИ НЕЗНАЧИТЕЛЬНО
ИЗМЕНЯЕТСЯ
а) длина мышцы
б) напряжение мышцы
в) длина и напряжение мышцы
21
022. ПРИ ИЗОМЕТРИЧЕСКОМ СОКРАЩЕНИИ НЕЗНАЧИТЕЛЬНО
ИЗМЕНЯЕТСЯ
а) длина мышцы
б) напряжение мышцы
в) длина и напряжение мышцы
023. НА ОДНО ДВИГАТЕЛЬНОЕ НЕРВНОЕ ВОЛОКНО
НАИБОЛЬШЕЕ ЧИСЛО МЫШЕЧНЫХ ВОЛОКОН ПРИХОДИТСЯ В
СЛЕДУЮЩИХ МЫШЦАХ
а) в глазодвигательных
б) в мышцах, фиксирующих конечности
в) в сгибателях пальцев ноги
г) в разгибателях пальцев руки
024. ПЛАВНОСТЬ ДВИЖЕНИЯ ОБЕСПЕЧИВАЕТСЯ
ПЛАВНОСТЬ ДВИЖЕНИЯ ОБЕСПЕЧИВАЕТСЯ
а) включением мышц антагонистов
б) асинхронным включением нервно-мышечных единиц
в) все перечисленное верно
025. СКЕЛЕТНАЯ МЫШЦА (МАКСИМАЛЬНО) МОЖЕТ УСВОИТЬ
СЛЕДУЮЩЕЕ КОЛИЧЕСТВО ИМПУЛЬСОВ, ЕСЛИ
ДЛИТЕЛЬНОСТЬ КАЖДОГО ПОТЕНЦИАЛА ДЕЙСТВИЯ
МЫШЕЧНОГО ВОЛОКНА СОСТАВЛЯЕТ 3-5 МС
а) 5000
б) 3000
в) 250-300
г) 200
026. ВОЗБУЖДЕНИЕ ПО НЕРВНЫМ ВОЛОКНАМ СКЕЛЕТНОЙ
МУСКУЛАТУРЫ) ПРОВОДИТСЯ СО СЛЕДУЮЩЕЙ СКОРОСТЬЮ
(М/С)
а) 12-14 м/с
б) 70-120 м/с
в) 140 м/с
г) 200 м/с
027. НАИБОЛЬШЕЙ ТОЛЩИНОЙ ОБЛАДАЮТ НИТИ
СЛЕДУЮЩЕГО БЕЛКА — АКТИНА ИЛИ МИОЗИНА
а) одинаковые
22
б) актина
в) миозина
028. ПРИ СОКРАЩЕНИИ МЫШЦ НИТИ АКТИНА
а) вдвигаются в промежутки между нитями миозина
б) укорачиваются
в) утолщаются
г) всё перечисленное верно
029. ПРИ СОКРАЩЕНИИ МЫШЕЧНОГО ВОЛОКНА, ИЗОТРОПНЫЙ ДИСК
а) укорачивается
б) удлиняется
в) не изменяется
030. БОЛЕЕ НИЗКИЙ ПОРОГ РАЗДРАЖЕНИЯ ИМЕЕТ
а) нерв
б) мышца
031. В ОРГАНИЗМЕ УТОМЛЯЕТСЯ БЫСТРЕЕ СЛЕДУЮЩИЙ
В ОРГАНИЗМЕ УТОМЛЯЕТСЯ БЫСТРЕЕ СЛЕДУЮЩИЙ
ЭЛЕМЕНТ НЕЙРОМОТОРНОГО АППАРАТА
а) нерв
б) мионевральный синапс
в) мышца
г) двигательный центр
032. ПРАВИЛО СРЕДНИХ НАГРУЗОК ИМЕЕТ СЛЕДУЮЩУЮ
ФОРМУЛИРОВКУ
а) работа средней величины совершается при средних нагрузках
б) максимальная работа совершается при средних нагрузках
в) минимальная работа совершается при средних нагрузках
033. АВТОР ТЕОРИИ АКТИВНОГО ОТДЫХА
а) И. М. Сеченов
б) И. П. Павлов
в) Н. Е. Введенский
034. ЛАБИЛЬНОСТЬ СКЕЛЕТНОЙ МЫШЦЫ РАВНА
а) 500-1000
23
б) 50-100
в) 300-200
035. ЛАБИЛЬНОСТЬ СИНАПСА РАВНА
а) 500-1000
б) 50-100
в) 300-200
036. ДВИГАТЕЛЬНАЯ ЕДИНИЦА ВКЛЮЧАЕТ В СЕБЯ
СЛЕДУЮЩИЕ СТРУКТУРЫ
а) мотонейрон
б) мотонейрон и иннервируемые им мышечные волокна
в) мышцу
037. ИЗОТОНИЧЕСКИМ НАЗЫВАЕТСЯ СЛЕДУЮЩИЙ РЕЖИМ
СОКРАЩЕНИЯ МЫШЦ
а) сокращение, при котором изменяется длина мышцы при постоянном
ее напряжении
б) сокращение, при котором изменяется напряжение при постоянной
ее длине
в) сокращение, при котором изменяется и напряжение, и длина мышц
038. ИЗОМЕТРИЧЕСКИМ НАЗЫВАЕТСЯ СЛЕДУЮЩИЙ РЕЖИМ
ИЗОМЕТРИЧЕСКИМ НАЗЫВАЕТСЯ СЛЕДУЮЩИЙ РЕЖИМ
СОКРАЩЕНИЯ МЫШЦ
а) сокращение, при котором изменяется длина мышцы при постоянном
ее напряжении
б) сокращение, при котором изменяется напряжение при постоянной
ее длине
в) сокращение, при котором изменяется и напряжение, и длина мышц
039. ПРИ МЕНЬШЕЙ ЧАСТОТЕ СТИМУЛЯЦИИ ВОЗНИКАЕТ
СЛЕДУЮЩИЙ ТИП СОКРАЩЕНИЯ МЫШЦЫ
а) одиночное сокращение
б) зубчатый тетанус
в) гладкий тетанус
040. БОЛЬШЕЙ СИЛОЙ ОБЛАДАЮТ СЛЕДУЮЩИЕ МЫШЦЫ
а) продольные
б) косые
24
в) перистые
г) гладкие
041. ПОД ПЛАСТИЧНОСТЬЮ ГЛАДКОЙ МЫШЦЫ ПОНИМАЮТ
а) способность перистальтически сокращаться
б) способность сохранять длину после растяжения
в) способность усиливать сокращение после растяжения
042. САРКОМЕРОМ НАЗЫВАЕТСЯ СЛЕДУЮЩИЙ УЧАСТОК
МИОФИБРИЛЛЫ
а) ограниченный двумя H-полосками
б) от начала A-диска до начала следующего A-диска
в) от начала I-диска до начала следующего I-диска
г) ограниченный двумя Z-мембранами
043. В СОКРАЩЕНИИ МЫШЦЫ УЧАСТВУЮТ СЛЕДУЮЩИЕ
В СОКРАЩЕНИИ МЫШЦЫ УЧАСТВУЮТ СЛЕДУЮЩИЕ
РЕГУЛИРУЮЩИЕ БЕЛКИ
а) тропонин и тропомиозин
б) тяжелый и легкий меромиозин
в) актин
044. БЛОКАТОРОМ ХОЛИНОРЕЦЕПТОРОВ ЯВЛЯЕТСЯ
а) тетраэтиламмоний
б) тетродотоксин
в) верапамил
г) кураре
045. ХЕМОРЕЦЕПТОРЫ МИОНЕВРАЛЬНОГО СИНАПСА
ОТНОСЯТСЯ К СЛЕДУЮЩЕМУ КЛАССУ РЕЦЕПТОРОВ
а) н-холинорецепторы
б) м-холинорецепторы
в) альфа-адренорецепторы
г) бета-адренорецепторы
046. К КАКОМУ ТИПУ СИНАПСОВ ОТНОСИТСЯ МИОНЕВРАЛЬНЫЙ СИНАПС
а) адренэргический
б) холинэргический
в) гистаминэргический
25
г) глицинэргический
047. БЫСТРЕЕ УТОМЛЯЕТСЯ СЛЕДУЮЩИЙ ЭЛЕМЕНТ НЕРВНОМЫШЕЧНОГО ПРЕПАРАТА
а) нерв
б) синапс
в) мышца
048. ПЕССИМУМ — ЭТО
а) утомление
б) торможение
в) длительное суммирование сокращения
г) остаточное сокращение после выполнения работы
049. КРИВАЯ УТОМЛЕНИЯ — ЭТО
а) кимограмма
б) кардиограмма
в) электромиограмма
г) эргограмма
050. МЕЖДУ АМПЛИТУДОЙ СОКРАЩЕНИЯ ЦЕЛОЙ МЫШЦЫ И
МЕЖДУ АМПЛИТУДОЙ СОКРАЩЕНИЯ ЦЕЛОЙ МЫШЦЫ И
СИЛОЙ РАЗДРАЖИТЕЛЯ СУЩЕСТВУЕТ СЛЕДУЮЩАЯ
ЗАВИСИМОСТЬ
а) по типу «всё или ничего»
б) чем больше сила раздражителя, тем больше амплитуда сокращения
(в определенном диапазоне)
в) чем больше сила раздражителя, тем меньше амплитуда сокращения
(в определенном диапазоне)
051. ЗАВИСИМОСТЬ АМПЛИТУДЫ СОКРАЩЕНИЯ ЦЕЛОЙ
МЫШЦЫ ОТ СИЛЫ РАЗДРАЖИТЕЛЯ ОБОЗНАЧАЕТСЯ
СЛЕДУЮЩИМ ТЕРМИНОМ
а) закон «всё или ничего»
б) лестничная зависимость
в) обратная гиперболическая зависимость
г) линейная зависимость
052. МЕЖДУ РАЗДРАЖЕНИЯМИ ВОЗНИКАЕТ ГЛАДКИЙ
ТЕТАНУС ПРИ СЛЕДУЮЩЕМ ИНТЕРВАЛЕ
26
а) длиннее периода рефрактерности, но короче длительности цикла
одиночного сокращения
б) длиннее периода рефрактерности, но короче длительности фазы
укорочения
в) длиннее цикла одиночного сокращения
г) длиннее фазы укорочения, но короче цикла одиночного сокращения
053. НЕПОСРЕДСТВЕННОЙ ПРИЧИНОЙ СОКРАЩЕНИЯ МЫШЦЫ
ЯВЛЯЕТСЯ
а) потенциал действия
б) локальный ответ
в) растяжение
054. ПОТЕНЦИАЛ ПОКОЯ СКЕЛЕТНОЙ МЫШЕЧНОЙ КЛЕТКИ
ПОТЕНЦИАЛ ПОКОЯ СКЕЛЕТНОЙ МЫШЕЧНОЙ КЛЕТКИ
РАВЕН
а) —50 мВ
б) —70 мВ
в) —90 мВ
г) +30 мВ
055. ЗНАЧЕНИЕ ХОЛИНЭСТЕРАЗЫ В ФУНКЦИОНИРОВАНИИ
НЕРВНО-МЫШЕЧНОГО СИНАПСА ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО
ЭТОТ ФЕРМЕНТ
а) расщепляет ацетилхолин в синаптической щели
б) способствует синтезу ацетилхолина
в) обеспечивает взаимодействие ацетилхолина с хеморецепторами
г) инактивирует хеморецепторы
056. ЧТОБЫ ДОКАЗАТЬ В ЭКСПЕРИМЕНТЕ, ЧТО В
ИЗОЛИРОВАННОЙ МЫШЦЕ УТОМЛЕНИЕ СВЯЗАНО С
НАКОПЛЕНИЕМ ПРОДУКТОВ ОБМЕНА, НУЖНО
а) сменить физиологический раствор
б) увеличить раздражение нерва
в) увеличить нагрузку на мышцу
057. ДЛЯ ТОГО, ЧТОБЫ ДОКАЗАТЬ, ЧТО ПЕССИМУМ НЕ
СВЯЗАН С УТОМЛЕНИЕМ МИОНЕВРАЛЬНОГО СИНАПСА,
НУЖНО ИСПОЛЬЗОВАТЬ СЛЕДУЮЩИЙ ПРИЕМ
а) уменьшить частоту раздражений до оптимальной
27
б) увеличить частоту раздражений выше пессимальной
в) прекратить раздражения
058. УВЕЛИЧЕНИЕ СИЛЫ РАЗДРАЖЕНИЯ МЫШЦЫ УВЕЛИЧИВАЕТ СИЛУ ЕЕ СОКРАЩЕНИЯ, ПОТОМУ ЧТО
а) каждая миофибрилла увеличивает амплитуду сокращения
б) увеличивается количество сокращающихся миофибрилл
в) повышается возбудимость миофибрилл
059. ВМЕСТО ОДИНОЧНЫХ СОКРАЩЕНИЙ ВОЗНИКАЕТ
ВМЕСТО ОДИНОЧНЫХ СОКРАЩЕНИЙ ВОЗНИКАЕТ
ЗУБЧАТЫЙ ТЕТАНУС ПРИ СЛЕДУЮЩЕМ ИНТЕРВАЛЕ МЕЖДУ
РАЗДРАЖЕНИЯМИ
а) длиннее фазы рефрактерности, но короче длительности цикла одиночного сокращения
б) длиннее фазы рефрактерности, но короче длительности фазы укорочения
в) длиннее цикла одиночного сокращения
г) длиннее фазы укорочения, но короче цикла одиночного сокращения
28
РАЗДЕЛ 03. ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ
СИСТЕМЫ
Выберите один или несколько правильных ответов из предлагаемых.
001. РЕФЛЕКСОМ НАЗЫВАЕТСЯ
а) совокупность рецепторов, составляющих рецептивное поле
б) ответная реакция организма на действие адекватных раздражителей
при обязательном участии ЦНС
в) время от начала действия раздражителя до ответной реакции
г) совокупность нервных клеток
002. ЗВЕНЬЯМИ РЕФЛЕКТОРНОЙ ДУГИ ЯВЛЯЮТСЯ
а) рецептор, синапс, эффектор
б) рецептор, афферентный чувствительный нейрон, нервный центр,
эфферентный двигательный нейрон, рабочий орган
в) рецептор, ЦНС, рабочий орган
г) афферентный нейрон, рабочий орган
003. К СВОЙСТВАМ НЕРВНЫХ ЦЕНТРОВ ОТНОСЯТСЯ
К СВОЙСТВАМ НЕРВНЫХ ЦЕНТРОВ ОТНОСЯТСЯ
СЛЕДУЮЩИЕ ИЗ УКАЗАННЫХ ЯВЛЕНИЙ
а) одностороннее проведение возбуждения
б) синаптическая задержка
в) суммация возбуждений
г) иррадиация
004. ТОРМОЖЕНИЕ — ЭТО
а) это пассивный процесс
б) процесс, направленный на полное подавление возбуждения
в) самостоятельный физиологический процесс, который вызывается
возбуждением и направлен на подавление другого возбуждения
г) ответная реакция организма на изменяющиеся условия внешней среды
005. ДОМИНАНТА — ЭТО
а) угнетение процесса высвобождения медиатора
б) деполяризация постсинаптической мембраны
в) временно господствующий очаг возбуждения ЦНС
29
г) постсинаптическая потенциация
006. ОСНОВНЫЕ СВОЙСТВА ДОМИНАНТНОГО ОЧАГА — ЭТО
а) длительность возбуждения
б) стойкость возбуждения
в) инертность
г) окклюзия
007. ЗНАЧЕНИЕ ЦНС ДЛЯ ОРГАНИЗМА СОСТОИТ В ТОМ, ЧТО
а) ЦНС обеспечивает связь различных органов и систем
б) ЦНС осуществляет связь организма с внешней средой
в) ЦНС осуществляет процессы сознания и мышления
г) ЦНС регулирует работу внутренних органов
008. СПИННОЙ МОЗГ ИМЕЕТ СЛЕДУЮЩЕЕ ЧИСЛО СЕГМЕНТОВ
СПИННОЙ МОЗГ ИМЕЕТ СЛЕДУЮЩЕЕ ЧИСЛО СЕГМЕНТОВ
а) 20
б) 21
в) 41
г) 31
009. В ШЕЙНЫХ СЕГМЕНТАХ СПИННОГО МОЗГА НАХОДЯТСЯ
СЛЕДУЮЩИЕ ИЗ ПЕРЕЧИСЛЕННЫХ ЦЕНТРОВ
а) центры межреберных нервов
б) центры диафрагмального нерва
в) центры симпатических нервов
г) центр рвоты
010. РОМБОВИДНАЯ ЯМКА НАХОДИТСЯ
а) в продолговатом мозге
б) в варолиевом мосту
в) в гипоталамусе
г) на дне IV желудочка
011. В ГРУДНЫХ СЕГМЕНТАХ СПИННОГО МОЗГА РАСПОЛАГАЮТСЯ СЛЕДУЮЩИЕ ЦЕНТРЫ
а) центры межреберных нервов
б) центры диафрагмального нерва
в) центры тазовых нервов
г) центр рвоты
30
012. В ПОЯСНИЧНЫХ СЕГМЕНТАХ СПИННОГО МОЗГА
РАСПОЛАГАЮТСЯ СЛЕДУЮЩИЕ ЦЕНТРЫ
а) центры мочеиспускания, половых рефлексов
б) центры, регулирующие сокращение мышц нижних конечностей
в) дыхательный центр
г) центры регуляции вегетативных функций
013. В ВАРОЛИЕВОМ МОСТУ РАСПОЛАГАЮТСЯ ЯДРА
СЛЕДУЮЩИХ ЧЕРЕПНО-МОЗГОВЫХ НЕРВОВ
а) с V по VIII
б) с I по III
в) с III по V
г) с VIII по IX
014. В ПРОДОЛГОВАТОМ МОЗГЕ РАСПОЛОЖЕНЫ ЦЕНТРЫ
В ПРОДОЛГОВАТОМ МОЗГЕ РАСПОЛОЖЕНЫ ЦЕНТРЫ
СЛЕДУЮЩИХ ЗАЩИТНЫХ РЕФЛЕКСОВ
а) мигания
б) чихания, кашля
в) рвоты
г) оборонительный
015. В КРЕСТЦОВЫХ СЕГМЕНТАХ СПИННОГО МОЗГА
НАХОДЯТСЯ СЛЕДУЮЩИЕ ЦЕНТРЫ
а) центры диафрагмального нерва
б) центры межреберных нервов
в) центры мочеиспускания и дефекации
г) центры парасимпатических нервов
016. К СРЕДНЕМУ МОЗГУ ОТНОСЯТСЯ СЛЕДУЮЩИЕ ОБРАЗОВАНИЯ
а) зрительные бугры
б) пластинка четверохолмия
в) ножки мозга
г) оливы
017. ПРИ УДАЛЕНИИ МОЗЖЕЧКА ОТМЕЧАЮТСЯ СЛЕДУЮЩИЕ
ИЗ ПЕРЕЧИСЛЕННЫХ ДВИГАТЕЛЬНЫХ РАССТРОЙСТВ
а) снижение силы мышечных сокращений
31
б) потеря способности к слитным тетаническим сокращениям
в) резкое повышение тонуса сгибателей
г) непроизвольное слюноотделение
018. АСТАЗИЯ — ЭТО
а) снижение силы мышечных сокращений
б) быстрая мышечная утомляемость
в) потеря способности к тетаническому мышечному сокращению
г) нарушение координации движений
019. АТАКСИЯ — ЭТО
АТАКСИЯ — ЭТО
а) нарушение координации движений
б) дрожание
в) быстрая мышечная утомляемость
г) снижение силы мышечных сокращений
020. ВЕРХНИЕ БУГРЫ ЧЕТВЕРОХОЛМИЯ ВЫПОЛНЯЮТ
СЛЕДУЮЩИЕ ФУНКЦИИ
а) ориентировочные реакции на звук
б) ориентировочные реакции на свет
в) распределение мышечного тонуса
г) формирование болевой чувствительности
021. КРАСНЫЕ ЯДРА И ЧЕРНАЯ СУБСТАНЦИЯ ВЫПОЛНЯЮТ
СЛЕДУЮЩИЕ ФУНКЦИИ
а) регулируют и распределяют тонус мышц
б) координируют движения тела
в) контролируют мелкие движения пальцев
г) формируют болевую чувствительность
022. НИЖНИЕ БУГРЫ ЧЕТВЕРОХОЛМИЯ ВЫПОЛНЯЮТ
СЛЕДУЮЩИЕ ФУНКЦИИ
а) ориентировочные реакции на звук
б) ориентировочные реакции на свет
в) распределение мышечного тонуса
г) формирование болевой чувствительности
023. ДЕЦЕРЕБРАЦИОННАЯ РИГИДНОСТЬ — ЭТО
32
а) резкое повышение тонуса разгибателей, возникающее при перерезке
ствола между продолговатым и средним мозгом
б) резкое повышение тонуса сгибателей, возникающее при перерезке
ствола между продолговатым и средним мозгом
в) резкое повышение тонуса разгибателей, возникающее при перерезке
ствола между продолговатым и спинным мозгом
г) резкое повышение тонуса сгибателей, возникающее при перерезке
ствола между продолговатым и спинным мозгом
024. ЗРИТЕЛЬНЫЕ БУГРЫ ВЫПОЛНЯЮТ СЛЕДУЮЩИЕ
ЗРИТЕЛЬНЫЕ БУГРЫ ВЫПОЛНЯЮТ СЛЕДУЮЩИЕ
ФУНКЦИИ
а) первичная обработка афферентной импульсации
б) формирование протопатической болевой чувствительности
в) координация движений
г) поддержание мышечного тонуса
025. ПРИ РАЗДРАЖЕНИИ РЕТИКУЛЯРНОЙ ФОРМАЦИИ ВО
ВРЕМЯ СНА У ЭКСПЕРИМЕНТАЛЬНОГО ЖИВОТНОГО
а) наступит кома
б) наступит пробуждение
в) наступит углубление сна
г) будет наблюдаться уравнительная фаза парабиоза
026. ЕСЛИ РЕТИКУЛЯРНУЮ ФОРМАЦИЮ РАЗРУШИТЬ ИЛИ
ПОДАВИТЬ ЛЕКАРСТВАМИ, ТО У ЭКСПЕРИМЕНТАЛЬНОГО
ЖИВОТНОГО
а) наступает возбуждение
б) возникает тремор
в) будет наблюдаться длительный сон
027. В ЛИМБИЧЕСКОЙ СИСТЕМЕ РАСПОЛОЖЕНЫ ЦЕНТРЫ
СЛЕДУЮЩИХ ФУНКЦИЙ
а) центры вегетативных функций
б) центры положительных и отрицательных эмоций
в) центры памяти
г) центры обоняния и осязания
33
028. В ОКОНЧАНИЯХ ПРЕГАНГЛИОНАРНЫХ НЕЙРОНОВ
ПАРАСИМПАТИЧЕСКОЙ СИСТЕМЫ ВЫРАБАТЫВАЕТСЯ
СЛЕДУЮЩИЙ МЕДИАТОР
а) ГАМК
б) серотонин
в) ацетилхолин
г) норадреналин
029. В ОКОНЧАНИЯХ ПРЕГАНГЛИОНАРНЫХ НЕЙРОНОВ
В ОКОНЧАНИЯХ ПРЕГАНГЛИОНАРНЫХ НЕЙРОНОВ
СИМПАТИЧЕСКОЙ СИСТЕМЫ ВЫРАБАТЫВАЕТСЯ
СЛЕДУЮЩИЙ МЕДИАТОР
а) ГАМК
б) норадреналин
в) серотонин
г) ацетилхолин
030. ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ
СИСТЕМЫ ОКАЗЫВАЕТ СЛЕДУЮЩИЕ ВЛИЯНИЯ
а) замедляет и ослабляет сокращения сердца
б) сужает бронхи
в) сужает зрачок
г) ослабляет моторику и секрецию пищеварительных органов
031. СИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ
СИСТЕМЫ ОКАЗЫВАЕТ СЛЕДУЮЩИЕ ВЛИЯНИЯ
а) расширяет бронхи
б) суживает зрачок
в) усиливает моторику ЖКТ
032. МЕСТО РАСПОЛОЖЕНИЯ ХОЛИНОРЕЦЕПТОРОВ
а) в нервномышечных синапсах
б) в вегетативных ганглиях
в) в спинномозговых ганглиях
г) в синапсах на внутренних органах
033. МЕСТО РАСПОЛОЖЕНИЯ Н-ХОЛИНОРЕЦЕПТОРОВ
а) в ганглиях
б) в мышцах
в) в сердце
34
г) в ЖКТ
034. МЕСТО РАСПОЛОЖЕНИЯ ТЕЛ РЕЦЕПТОРНЫХ НЕЙРОНОВ
а) в спинномозговых ганглиях
б) в черепно-мозговых ганглиях
в) в спинном мозге
г) во внутренних органах
035. РОЛЬ ХОЛИНЭСТЕРАЗЫ В СИНАПСАХ СОСТОИТ В ТОМ,
РОЛЬ ХОЛИНЭСТЕРАЗЫ В СИНАПСАХ СОСТОИТ В ТОМ,
ЧТО ЭТОТ ФЕРМЕНТ
а) передает возбуждение от пресинаптической к постсинаптической
мембране
б) участвует в синтезе ацетилхолина
в) разрушает ацетилхолин
г) вызывает торможение в синапсе
036. УКАЖИТЕ РЕЦЕПТИВНОЕ ПОЛЕ КОЛЕННОГО РЕФЛЕКСА
а) кожа коленной области
б) кожа живота
в) коленная чашечка
г) сухожилие четырехглавой мышцы бедра
037. СУЩЕСТВУЮТ СЛЕДУЮЩИЕ ПРОПРИОЦЕПТИВНЫЕ
РЕФЛЕКСЫ
а) рефлекс чихания, мигания
б) чесательный рефлекс
в) коленный, ахиллов
г) рвотный рефлекс
038. ЦЕНТРАЛЬНОЕ ВРЕМЯ РЕФЛЕКСА — ЭТО
а) время прохождения возбуждения от рецептора до исполнительного
органа
б) время прохождения импульса по афферентному волокну
в) время передачи возбуждения через центральную часть рефлекторной дуги
г) время прохождения импульса по эфферентному волокну
039. САМЫМ КОРОТКИМ ЯВЛЯЕТСЯ ВРЕМЯ СЛЕДУЮЩЕГО РЕФЛЕКСА
35
а) парасимпатического
б) моносинаптического двигательного
в) симпатического
г) полисинаптического двигательного
040. ДЛИТЕЛЬНОЕ РЕФЛЕКТОРНОЕ ПОСЛЕДЕЙСТВИЕ В ЦНС
ДЛИТЕЛЬНОЕ РЕФЛЕКТОРНОЕ ПОСЛЕДЕЙСТВИЕ В ЦНС
ОБЪЯСНЯЕТСЯ
а) следовой деполяризацией
б) циркуляцией нервных импульсов по замкнутым нейронным цепям
в) неодновременным поступлением импульсов по нервным волокнам в
ЦНС
г) следовой гиперполяризацией
041. С ОДНИМ МЕТАМЕРОМ СПИННОГО МОЗГА СВЯЗАНО
СЛЕДУЮЩЕЕ КОЛИЧЕСТВО МЕТАМЕРОВ ТЕЛА
а) три
б) два
в) один
г) четыре
042. ЗАКОН БЕЛЛА-МАЖАНДИ ГЛАСИТ
а) все афферентные волокна входят в спинной мозг через передние корешки, а эфферентные выходят через задние
б) все афферентные и эфферентные волокна входят в спинной мозг через задние корешки, а выходят через передние
в) все афферентные и эфферентные волокна входят в спинной мозг через передние корешки, а выходят через задние
г) все афферентные волокна входят в спинной мозг через задние корешки, а эфферентные выходят через передние
043. ЦЕНТР ДИАФРАГМАЛЬНОГО НЕРВА РАСПОЛАГАЕТСЯ В
СЛЕДУЮЩИХ СЕГМЕНТАХ СПИННОГО МОЗГА
а) в I-II сегментах шейного отдела
б) в V-VI сегментах грудного отдела
в) в III-IV сегментах шейного отдела
г) в I-II сегментах грудного отдела
044. ЦЕНТР СИМПАТИЧЕСКОГО ОТДЕЛА ВЕГЕТАТИВНОЙ
ЦЕНТР СИМПАТИЧЕСКОГО ОТДЕЛА ВЕГЕТАТИВНОЙ
НЕРВНОЙ СИСТЕМЫ РАСПОЛАГАЕТСЯ
36
а) в продолговатом мозге
б) в грудном и поясничном отделах спинного мозга
в) в поясничном и сакральном отделах спинного мозга
г) в среднем мозге
045. ЦЕНТРЫ ДЕФЕКАЦИИ И МОЧЕИСПУСКАНИЯ РАСПОЛАГАЕТСЯ В СЛЕДУЮЩИХ СЕГМЕНТАХ СПИННОГО МОЗГА
а) в грудных сегментах
б) в шейных сегментах
в) в поясничных сегментах
г) в крестцовых сегментах
046. СПИНАЛЬНЫМ НАЗЫВАЕТСЯ СЛЕДУЮЩЕЕ ЖИВОТНОЕ
а) животное, у которого сделан разрез на уровне четверохолмия
б) животное, у которого красное ядро отделено от продолговатого мозга
в) животное, у которого головной мозг перерезкой отделен от спинного
г) животное, у которого удалена кора головного мозга
047. СПИНАЛЬНЫЙ ШОК ВЫЗВАН
а) полной перерезкой спинного мозга
б) удалением коры больших полушарий
в) отделением красного ядра от продолговатого мозга
г) разрезом на уровне четверохолмия
048. ВЛИЯНИЕ СПИНАЛЬНОГО ШОКА ОБЪЯСНЯЕТСЯ
а) устранением супраспинальных влияний
б) отсутствием влияний вегетативной нервной системы
в) травматическим повреждением
г) нарушением стриопаллидарной системы
049. ЦЕНТР КОЛЕННОГО РЕФЛЕКСА НАХОДИТСЯ В СЛЕДУЮЩИХ СЕГМЕНТАХ СПИННОГО МОЗГА
ЦЕНТР КОЛЕННОГО РЕФЛЕКСА НАХОДИТСЯ В СЛЕДУЮЩИХ СЕГМЕНТАХ СПИННОГО МОЗГА
а) в II-III сегментах грудного отдела
б) в II-IV сегментах поясничного отдела
в) в крестцовых сегментах
г) II-IV сегментах шейного отдела
37
050. ЦЕНТР ЛОКТЕВОГО РЕФЛЕКСА НАХОДИТСЯ В СЛЕДУЮЩИХ СЕГМЕНТАХ СПИННОГО МОЗГА
а) в V-VI сегментах шейного отдела
б) в II-III сегментах грудного отдела
в) в II-V сегментах поясничного отдела
г) в крестцовых сегментах
051. ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ В ЦНС ОБЕСПЕЧИВАЮТ СЛЕДУЮЩИЕ СТРУКТУРНЫЕ ЭЛЕМЕНТЫ
а) пирамидные клетки
б) нет специальных элементов обеспечивающих данный вид торможения
в) мотонейроны
г) клетки Реншоу
052. ТОРМОЗНЫМ МЕДИАТОРОМ В СПИННОМ МОЗГЕ, УЧАСТВУЮЩИЙ В ПОСТСИНАПТИЧЕСКОМ ТОРМОЖЕНИИ, ЯВЛЯЕТСЯ
а) серотонин
б) глицин
в) адреналин
г) эндорфин
053. ПЕССИМАЛЬНОЕ РАЗДРАЖЕНИЕ РАЗВИВАЕТСЯ В СЛЕДУЮЩИХ СИНАПСАХ
а) в электрических
б) в тормозных
в) в возбуждающих
г) в смешанных синапсах
054. ПОД КООРДИНАЦИЕЙ В ЦНС ПОНИМАЮТ
ПОД КООРДИНАЦИЕЙ В ЦНС ПОНИМАЮТ
а) взаимодействие нейронов в ЦНС, которое обеспечивает торможение
б) взаимодействие нейронов в ЦНС, которое обеспечивает согласованность рефлекторных актов
в) взаимодействие нейронов в ЦНС, которое обеспечивает тонус нервных центров
г) взаимодействие нейронов в ЦНС, которое обеспечивает синаптическую депрессию
38
055. К ВОЗНИКНОВЕНИЮ ДОМИНАНТНОГО ОЧАГА МОГУТ
ПРИВЕСТИ СЛЕДУЮЩИЕ ФАКТОРЫ
а) сильные и частые импульсы с соответствующих рецепторов, гуморальные агенты
б) слабые импульсы с различных рецептивных полей
в) длительная следовая деполяризация
г) реверберация возбуждения
056. К УСЛОВНЫМ РЕФЛЕКСАМ ОТНОСЯТСЯ
а) миотатические рефлексы спинного мозга
б) вегетативные рефлексы
в) реакции организма приобретенные в процессе индивидуального развития
г) брюшные рефлексы спинного мозга
057. ПРИМЕР ЦЕПНОГО РЕФЛЕКСА
а) глотание
б) чихание
в) шагание
г) слезоотделение
058. ПЛАСТИЧНОСТЬ НЕРВНЫХ ЦЕНТРОВ — ЭТО
а) изменение их тонуса
б) изменение в них ритма импульсаций
в) циркуляция нервных импульсов по замкнутым нейронным цепям
г) способность к замещению утраченной функции
059. БУЛЬБАРНЫМ ЖИВОТНЫМ НАЗЫВАЕТСЯ
БУЛЬБАРНЫМ ЖИВОТНЫМ НАЗЫВАЕТСЯ
а) животное, у которого спинной мозг отделен от продолговатого
б) животное, у которого продолговатый мозг отделен от вышележащих
отделов
в) животное, у которого удален продолговатый мозг
г) животное, у которого удален варолиев мост
060. В РЕГУЛЯЦИИ ТОНУСА МЫШЦ ПРИНИМАЮТ УЧАСТИЕ
СЛЕДУЮЩИЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА
а) дорсальная группа ядер
б) вентральная группа ядер
в) бульбопонтинный центр
39
г) ядро вестибулярного нерва
061. ЦЕНТРЫ ЛАБИРИНТНЫХ И ШЕЙНЫХ ТОНИЧЕСКИХ РЕФЛЕКСОВ НАХОДЯТСЯ В СЛЕДУЮЩЕМ ОТДЕЛЕ ЦНС
а) в таламусе
б) в гипоталамусе
в) в продолговатом мозге
г) в спинном мозге
062. ЦЕНТРЫ ВЫПРЯМИТЕЛЬНЫХ РЕФЛЕКСОВ НАХОДЯТСЯ В
а) в спинном мозге
б) в продолговатом мозге
в) в среднем мозге
г) в таламусе
063. У МЕЗЭНЦЕФАЛЬНОГО ЖИВОТНОГО
а) нарушен тонус мышц-сгибателей
б) нарушен тонус мышц-разгибателей
в) мышечный тонус нормальный
г) нарушен пластический тонус
064. ЧЕРЕЗ ПЕРЕДНИЕ БУГРЫ ЧЕТВЕРОХОЛМИЯ ОСУЩЕСТВЛЯЮТСЯ СЛЕДУЮЩИЕ РЕФЛЕКСЫ
ЧЕРЕЗ ПЕРЕДНИЕ БУГРЫ ЧЕТВЕРОХОЛМИЯ ОСУЩЕСТВЛЯЮТСЯ СЛЕДУЮЩИЕ РЕФЛЕКСЫ
а) поворот головы и глаз в сторону светового раздражителя
б) поворот головы и глаз в сторону звукового раздражителя
в) оборонительный рефлекс
г) двигательный рефлекс
065. ЧЕРЕЗ ЗАДНИЕ БУГРЫ ЧЕТВЕРОХОЛМИЯ ОСУЩЕСТВЛЯЮТСЯ СЛЕДУЮЩИЕ РЕФЛЕКСЫ
а) поворот головы и глаз в сторону светового раздражителя
б) поворот головы и глаз в сторону звукового раздражителя
в) оборонительный рефлекс
г) двигательный рефлекс
066. КРАСНЫЕ ЯДРА СРЕДНЕГО МОЗГА ВЫПОЛНЯЮТ СЛЕДУЮЩИЕ ФУНКЦИИ
а) регуляция мышечного тонуса
б) регуляция дыхания
40
в) регуляция работы сердца
г) регуляция пищевых рефлексов
067. ЦЕНТРЫ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ ИМЕЮТ
СЛЕДУЮЩУЮ ЛОКАЛИЗАЦИЮ
а) крестцовый отдел спинного мозга
б) шейный отдел спинного мозга
в) грудной и поясничный отделы спинного мозга
г) сакральный отдел спинного мозга
068. В ПРЕГАНГЛИОНАРНЫХ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ВОЛОКНАХ ВЫДЕЛЯЕТСЯ СЛЕДУЮЩИЙ МЕДИАТОР
а) ацетилхолин
б) норадреналин
в) серотонин
г) ГАМК
069. ВЫСШИЕ ЦЕНТРЫ РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ РАСПОЛАГАЮТСЯ
ВЫСШИЕ ЦЕНТРЫ РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ РАСПОЛАГАЮТСЯ
а) в среднем мозге
б) в продолговатом мозге
в) в спинном мозге
г) в гипоталамусе
070. ВИСЦЕРАЛЬНЫЙ МОЗГ — ЭТО
а) средний мозг
б) таламус
в) гипоталамус
г) лимбическая система
071. ОСНОВНЫМИ ФУНКЦИЯМИ АКСОНА ЯВЛЯЮТСЯ
а) проведение нервного импульса на большое расстояние
б) синтез макромолекул
в) организация межклеточного взаимодействия
г) синтез ацетилхолина
072. СВОЙСТВО НЕРВНЫХ ЦЕНТРОВ, КОГДА ПОСЛЕ ПРЕКРАЩЕНИЯ ДЕЙСТВИЯ РАЗДРАЖИТЕЛЯ В ЦЕПИ НЕЙРОНОВ ОСТА41
ЕТСЯ СЛЕД ВОЗБУЖДЕНИЯ ЗА СЧЕТ СЛЕДОВОЙ ДЕПОЛЯРИЗАЦИИ, НАЗЫВАЕТСЯ
а) посттетаническая потенциация
б) последействие
в) центральное облегчение
г) инертность
073. К СВОЙСТВАМ ДОМИНАНТНОГО ОЧАГА ВОЗБУЖДЕНИЯ
НЕ ОТНОСИТСЯ СЛЕДУЮЩЕЕ ИЗ ПЕРЕЧИСЛЕННЫХ СВОЙСТВ
а) длительность возбуждения
б) стойкость возбуждения
в) быстрая утомляемость
г) инертность
074. ПОД РЕФЛЕКСОМ ДАНИНИ-АШНЕРА ПОДРАЗУМЕВАЕТСЯ
СЛЕДУЮЩЕЕ
а) урежение сердечных сокращений в конце выдоха перед началом
следующего вдоха
б) кратковременное урежение сердцебиений при надавливании на
глазные яблоки
в) учащение сердечных сокращений и повышении артериального давления при переходе из положения лежа в положение стоя
г) кратковременное учащение сердцебиений при надавливании на
глазные яблоки
075. ТЕЛА ДВИГАТЕЛЬНЫХ НЕЙРОНОВ СПИННОГО МОЗГА
ТЕЛА ДВИГАТЕЛЬНЫХ НЕЙРОНОВ СПИННОГО МОЗГА
РАСПОЛОЖЕНЫ
а) в задних рогах серого вещества спинного мозга;
б) в передних рогах серого вещества спинного мозга
в) в боковых рогах серого вещества спинного мозга
г) в спинномозговых узлов
076. ТЕЛА ВЕГЕТАТИВНЫХ ПРЕГАНГЛИОНАРНЫХ НЕЙРОНОВ
РАСПОЛОЖЕНЫ
а) в передних рогах серого вещества спинного мозга
б) в задних рогах серого вещества спинного мозга
в) в боковых рогах серого вещества спинного мозга
г) в спинномозговых узлах
42
077. ЗАДНИЕ КОРЕШКИ СПИННОГО МОЗГА СЧИТАЮТ
а) двигательными
б) чувствительными
в) ни теми, ни другими
г) смешанными
078. ПРИ ПОЛНОМ ПОВРЕЖДЕНИИ (РАЗРУШЕНИИ) ПРОДОЛГОВАТОГО МОЗГА НАБЛЮДАЕТСЯ СЛЕДУЮЩЕЕ
а) ухудшение функций дыхания и кровообращения
б) нарушение равновесия
в) нарушение пищеварительной функции
г) гибель организма
079. ПРИ РАЗДРАЖЕНИИ ПЕРЕДНЕЙ ГРУППЫ ЯДЕР ГИПОТАЛАМУСА НАБЛЮДАЕТСЯ
а) парасимпатический эффект
б) симпатический эффект
в) комплекс эмоциональных реакций
г) нарушение обмена веществ
080.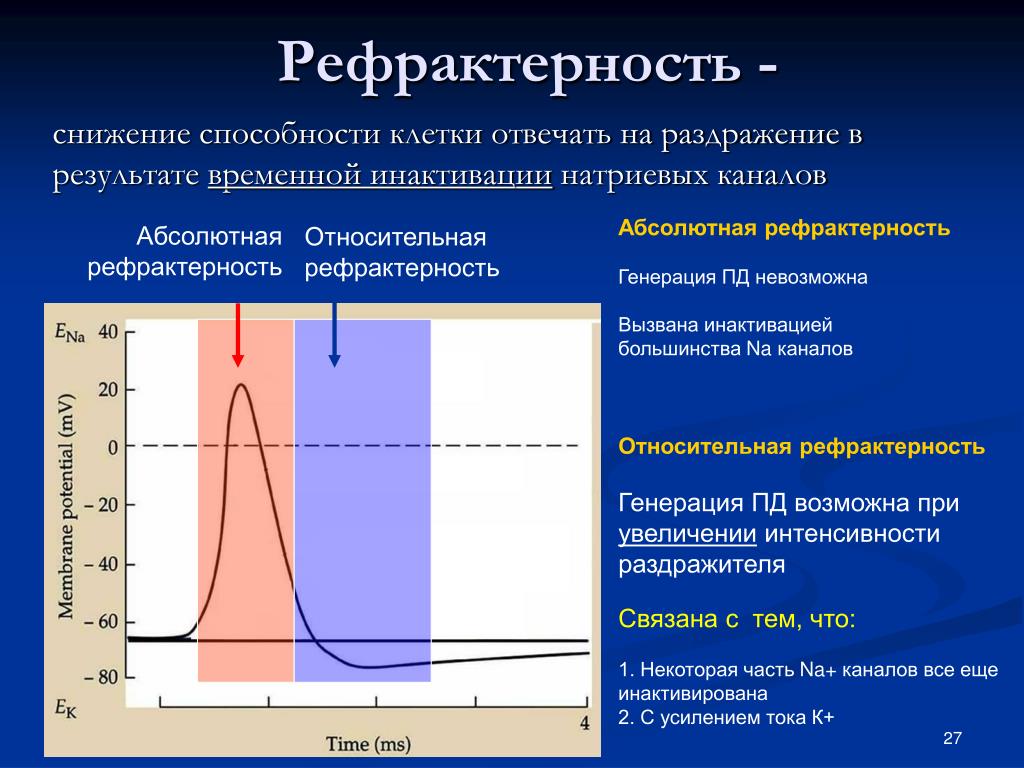 ПРИ РАЗДРАЖЕНИИ ЗАДНЕЙ ГРУППЫ ЯДЕР ГИПОТАЛАМУСА НАБЛЮДАЕТСЯ
ПРИ РАЗДРАЖЕНИИ ЗАДНЕЙ ГРУППЫ ЯДЕР ГИПОТАЛАМУСА НАБЛЮДАЕТСЯ
а) парасимпатический эффект
б) нарушение обмена веществ
в) симпатический эффект
г) болевой синдром
081. ЯВЛЕНИЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ ОТКРЫЛ
а) Декарт
б) Сеченов
в) Павлов
г) Прохазка
082. УЧЕНИЕ О ДОМИНАНТЕ БЫЛО РАЗРАБОТАНО СЛЕДУЮЩИМ УЧЕНЫМ
а) Сеченовым
б) Павловым
в) Введенским
г) Ухтомским
43
083. ГЛАВНЫЙ ПАРАСИМПАТИЧЕСКИЙ НЕРВ ЯВЛЯЕТСЯ
а) IX парой черепно-мозговых нервов
б) X парой черепно-мозговых нервов
в) XI парой черепно-мозговых нервов
г) VII парой черепно-мозговых нервов
084. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА ОБЕСПЕЧИВАЕТ
а) восприятие раздражений
б) сокращение скелетных мышц
в) поведение
г) трофику и адаптацию
085. СОМАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА ОБЕСПЕЧИВАЕТ
а) сокращение гладких мышц
б) адаптацию и трофику
в) сокращение скелетных мышц
г) обмен веществ
086. РЕФЛЕКС СУЖЕНИЯ ЗРАЧКА И БРОНХОВ ОСУЩЕСТВЛЯЕТ
СЛЕДУЮЩАЯ ЧАСТЬ АВТОНОМНОЙ НЕРВНОЙ СИСТЕМЫ
а) соматическая
б) симпатическая
в) метасимпатическая
г) парасимпатическая
087. ОПОРОЖНЕНИЕ ЖЕЛЧНОГО И МОЧЕВОГО ПУЗЫРЯ, ПРЯМОЙ КИШКИ НАБЛЮДАЕТСЯ ПРИ РАЗДРАЖЕНИИ СЛЕДУЮЩИХ НЕРВОВ
ОПОРОЖНЕНИЕ ЖЕЛЧНОГО И МОЧЕВОГО ПУЗЫРЯ, ПРЯМОЙ КИШКИ НАБЛЮДАЕТСЯ ПРИ РАЗДРАЖЕНИИ СЛЕДУЮЩИХ НЕРВОВ
а) соматических
б) симпатических
в) парасимпатических
088. ОККЛЮЗИЯ ХАРАКТЕРИЗУЕТСЯ И ВОЗНИКАЕТ
а) усилением рефлекторного ответа при перекресте центральных зон
нервных центров
б) ослаблением рефлекторного ответа при перекресте центральных зон
нервных центров
44
в) усилением рефлекторного ответа при перекресте периферических
зон нервных центров
г) ослаблением рефлекторного ответа при перекресте периферических
зон нервных центров.
089. ЦЕНТРАЛЬНОЕ ОБЛЕГЧЕНИЕ ХАРАКТЕРИЗУЕТСЯ И ВОЗНИКАЕТ
а) усилением рефлекторного ответа при перекресте периферических
зон нервных центров
б) ослаблением рефлекторного ответа при перекресте периферических
зон нервных центров
в) усилением рефлекторного ответа при перекресте центральных зон
нервных центров
г) ослаблением рефлекторного ответа при перекресте центральных зон
нервных центров
3.
 Подпороговые мембранные явления
Подпороговые мембранные явления
Arvanitaki A (1938): Les Variations Gradues De La Polarization Des Systmes Excitables , Герман, Париж.
Bernstein J (1902): Untersuchungen zur Termodynamik der bioelektrischen Strme. Pflger Arch. ges. Physiol . 9: 521-62.
Bernstein J (1912): Elektrobiologie , 215 стр. Viewag, Брауншвейг.
Дэвис Л.Дж., Лоренте де Н.Р. (1947): Вклад в математическую теорию электротонуса. Шпилька. Rockefeller Inst. Med. Res. 131: 442-96.
Eccles JC (1968): The Physiology of Nerve Cells , 270 pp. Johns Hopkins Press, Балтимор.
Эйнштейн A (1905): ber die von der molkularkinetischen Theorie die wrme Gefordertebewegung von in ruhenden flssigkeiten suspendierten Teilchen. Ann. Physik 17: 549-60.
Фарадей М (1834): Экспериментальные исследования электричества, 7-я серия. Фил. Пер. R. Soc. (Лондон.) 124: 77-122.
Фик М (1855): диффузия. Ann. Physik und Chemie 94: 59-86.
Гольдман Д.Е. (1943): потенциал, сопротивление и выпрямление в мембранах. J. Gen. Physiol. 27: 37-60.
Герман Л (1905): Beitrge zur Physiologie und Physik des Nerven. Pflger Arch. ges. Physiol. 109: 95-144.
Hille B (1992): Ionic Channels of Excitable Membranes , 2nd ed., 607 pp. Sinauer Assoc., Sunderland, Mass.(1-е изд., 1984)
Ходжкин А.Л., Хаксли А.Ф. (1952): количественное описание мембранного тока и его применение к проводимости и возбуждению в нерве. J. Physiol. (Лондон) 117: 500-44.
Ходжкин А.Л., Хаксли А.Ф., Кац Б. (1952): Измерение вольт-амперных отношений в мембране гигантского аксона Loligo. J. Physiol. (Лондон) 116: 424-48.
Ходжкин А.Л., Кац Б. (1949): Влияние ионов натрия на электрическую активность гигантского аксона кальмара. J. Physiol. (Лондон) 108: 37-77.
Ходжкин А.Л., Раштон В.А. (1946): электрические константы нервного волокна ракообразных. Proc. R. Soc. (Биол.) B133: 444-79.
R. Soc. (Биол.) B133: 444-79.
Katchalsky A, Curran PF (1965): Неравновесная термодинамика в биофизике, 248 стр. Harvard University Press, Cambridge, Mass.
Kortum G (1965): Трактат по электрохимии , 2-е изд., 637 стр. Elsevier, New York.
Nernst WH (1888): Zur Kinetik der Lsung befindlichen Krper: Theorie der Diffusion. Z. Phys. Chem. 3: 613-37.
Nernst WH (1889): Die elektromotorische Wirksamkeit der Ionen. Z. Phys. Chem. 4: 129-81.
Planck M (1890a): ber die Erregung von Elektricitt und Wrme in Elektrolyten. Ann. Physik und Chemie, Neue Folge 39: 161-86.
Planck M (1890b): ber die Potentialdifferenz zwischen zwei verdnnten Lsungen binrer Elektrolyte. Ann. Physik und Chemie, Neue Folge 40: 561-76.
Рух Т.С., Паттон HD (ред.) (1982): Physiology and Biophysics , 20-е изд., 1242 стр. У. Б. Сондерс, Филадельфия.
Rysselberghe van P (1963): Термодинамика необратимых процессов , 165 с. Герман, Париж, и Блезедель, Нью-Йорк.
Герман, Париж, и Блезедель, Нью-Йорк.
Эдсалл Дж. Т., Вайман Дж. (1958): Biophysical Chemistry , Vol. 1, 699 стр. Academic Press, New York.
Мур WJ (1972): Physical Chemistry , 4-е изд., 977 стр. Prentice-Hall, Englewood Cliffs, N.J.
Подпороговые колебания потенциала мембраны — обзор
4.37.9.1 Колебания
Колебания и синхронная активность являются характерными чертами спонтанной и обусловленной запахом активности МОБ и других обонятельных структур. Детальное рассмотрение колебаний в MOB выходит за рамки настоящего обзора и недавно было рассмотрено в другом месте (Lledo, PM et al. , 2005; Gelperin, A., 2006; Kepecs, A. et al. ). , 2006). Заметная осцилляторная активность присутствует в потенциалах поля MOB, которые в первую очередь отражают синхронизированные подпороговые мембранные потенциалы, возникающие из больших популяций нейронов, или в мембранном потенциале или спайковой активности отдельных нейронов MOB. Сенсорное воздействие на обонятельную систему происходит в результате дыхания, которое по самой своей природе является циклическим или ритмичным. В тета-диапазоне (2–12 Гц) возникают относительно низкочастотные колебания, синхронизированные с дыханием, при наличии и отсутствии запахов. Колебания более высокой частоты, наложенные на тета, происходят в бета (15–40 Гц) и гамма (30–80 Гц) диапазонах. Осцилляции бета-частоты относительно плохо изучены, но недавно они обсуждались в другом месте (Невилл, К. Р. и Хаберли, Л.Б., 2003; Martin, C. et al. , 2004; Fletcher, M. L. et al. , 2005). Низкочастотная колебательная (0,1–0,03 Гц) и синхронная активность наблюдалась в срезах МОБ при определенных условиях (Пуополо, М. и Беллуцци, О., 2001).
Сенсорное воздействие на обонятельную систему происходит в результате дыхания, которое по самой своей природе является циклическим или ритмичным. В тета-диапазоне (2–12 Гц) возникают относительно низкочастотные колебания, синхронизированные с дыханием, при наличии и отсутствии запахов. Колебания более высокой частоты, наложенные на тета, происходят в бета (15–40 Гц) и гамма (30–80 Гц) диапазонах. Осцилляции бета-частоты относительно плохо изучены, но недавно они обсуждались в другом месте (Невилл, К. Р. и Хаберли, Л.Б., 2003; Martin, C. et al. , 2004; Fletcher, M. L. et al. , 2005). Низкочастотная колебательная (0,1–0,03 Гц) и синхронная активность наблюдалась в срезах МОБ при определенных условиях (Пуополо, М. и Беллуцци, О., 2001).
4.37.9.1.1 Тета-ритм
В MOB колебания от 2 до 12 Гц называются тета главным образом потому, что они занимают сильно перекрывающийся частотный диапазон с тета-колебаниями гиппокампа (Kay, LM and Laurent, G., 1999; Кей, Л. М., 2003; 2005). Как отмечалось выше, колебания тета-частоты управляются дыханием и, таким образом, синхронизируются или синхронизируются по фазе. Следовательно, конкретная тета-частота зависит от частоты обнюхивания (т. Е. От характера воздушного потока через носовой эпителий) и включает компоненты, относящиеся к низкочастотному (1–3 Гц) пассивному обнюхиванию, а также более высокочастотный компонент (5–12). Hz), характерная для активного исследовательского обнюхивания (Adrian, ED, 1950; Welker, WI, 1964; Macrides, F. et al., 1982; Экман, Ф. Х. и Фриман, У. Дж., 1990; Кей, Л. М. и Лоран, Г., 1999; Кей, Л. М., 2003). Как отмечалось ранее, подпороговые колебания мембранного потенциала митральных клеток и / или пиковая активность синхронизируются с определенными фазами дыхательного цикла. Манипуляции, которые нарушают или нарушают управляемый дыханием вход в MOB, нарушают активность митральных клеток и отделяют ее от дыхательного цикла (Philpot, B. D. et al. , 1997). Таким образом, тета-колебания, по-видимому, управляются паттерном респираторного воздействия на МОБ.
М., 2003; 2005). Как отмечалось выше, колебания тета-частоты управляются дыханием и, таким образом, синхронизируются или синхронизируются по фазе. Следовательно, конкретная тета-частота зависит от частоты обнюхивания (т. Е. От характера воздушного потока через носовой эпителий) и включает компоненты, относящиеся к низкочастотному (1–3 Гц) пассивному обнюхиванию, а также более высокочастотный компонент (5–12). Hz), характерная для активного исследовательского обнюхивания (Adrian, ED, 1950; Welker, WI, 1964; Macrides, F. et al., 1982; Экман, Ф. Х. и Фриман, У. Дж., 1990; Кей, Л. М. и Лоран, Г., 1999; Кей, Л. М., 2003). Как отмечалось ранее, подпороговые колебания мембранного потенциала митральных клеток и / или пиковая активность синхронизируются с определенными фазами дыхательного цикла. Манипуляции, которые нарушают или нарушают управляемый дыханием вход в MOB, нарушают активность митральных клеток и отделяют ее от дыхательного цикла (Philpot, B. D. et al. , 1997). Таким образом, тета-колебания, по-видимому, управляются паттерном респираторного воздействия на МОБ. Однако внутренняя мембрана и синаптические свойства увеличивают способность некоторых нейронов MOB увлекаться или синхронизироваться с ритмической респираторной активностью. Например, клетки ET в срезах MOB по своей природе генерируют всплески спайков с частотами в диапазоне от ~ 1 до 8 Гц и легко вовлекаются в структурированный сенсорный ввод в том же диапазоне. Таким образом, свойства этих клеток могут позволить им разряжаться синхронно с дыхательным циклом.
Однако внутренняя мембрана и синаптические свойства увеличивают способность некоторых нейронов MOB увлекаться или синхронизироваться с ритмической респираторной активностью. Например, клетки ET в срезах MOB по своей природе генерируют всплески спайков с частотами в диапазоне от ~ 1 до 8 Гц и легко вовлекаются в структурированный сенсорный ввод в том же диапазоне. Таким образом, свойства этих клеток могут позволить им разряжаться синхронно с дыхательным циклом.
4.37.9.1.2 Гамма-ритм
Гамма-колебания возникают во время одоранта или прямой стимуляции ON и, по-видимому, генерируются внутренне в MOB.Таким образом, они сохраняются, когда центробежный ввод в лампу прерывается или нарушается (Грей, К. М. и Скиннер, Дж. Э., 1988; Невилл, К. Р. и Хаберли, Л. Б., 2003). Как и в случае тета-осцилляций, гамма-осцилляции, вызванные запахом или ON, проявляются в ЭЭГ, потенциалах поля MOB, подпороговом мембранном потенциале и пиковой активности митральных клеток (Adrian, ED, 1950; Eeckman, FH and Freeman, WJ, 1990; Кей, Л. М. и Фриман, У.Дж., 1998; Кашивадани, Х., и др., , 1999; Дебарбье, Ф. et al. , 2003; Фридман Д. и Строубридж Б. В., 2003; Невилл, К. Р. и Хаберли, Л. Б., 2003; Lagier, S. et al. , 2004; Martin, C. et al. , 2004; Fletcher, M. L. et al. , 2005). Анализ плотности источников тока показывает, что гамма-колебания в потенциалах поля МОБ в первую очередь отражают синаптические токи, протекающие в ШС (Невилл, К. Р. и Хаберли, Л. Б., 2003). Записи с нескольких участков показали, что фаза гамма-колебаний может варьироваться в разных частях MOB, особенно при низких концентрациях запаха.Считается, что гамма-колебания возникают в основном в результате ритмических или реверберационных взаимодействий между митральными / пучковыми клетками и тормозящими интернейронами, в основном GC (Friedman, D. and Strowbridge, BW, 2003; Neville, KR and Haberly, LB, 2003; Lagier, S. и др. , 2004). Активность митральных клеток синхронизируется с потенциалами поля MOB с частотой γ, а также с ГАМКергическим синаптическим входом (Lagier, S.
М. и Фриман, У.Дж., 1998; Кашивадани, Х., и др., , 1999; Дебарбье, Ф. et al. , 2003; Фридман Д. и Строубридж Б. В., 2003; Невилл, К. Р. и Хаберли, Л. Б., 2003; Lagier, S. et al. , 2004; Martin, C. et al. , 2004; Fletcher, M. L. et al. , 2005). Анализ плотности источников тока показывает, что гамма-колебания в потенциалах поля МОБ в первую очередь отражают синаптические токи, протекающие в ШС (Невилл, К. Р. и Хаберли, Л. Б., 2003). Записи с нескольких участков показали, что фаза гамма-колебаний может варьироваться в разных частях MOB, особенно при низких концентрациях запаха.Считается, что гамма-колебания возникают в основном в результате ритмических или реверберационных взаимодействий между митральными / пучковыми клетками и тормозящими интернейронами, в основном GC (Friedman, D. and Strowbridge, BW, 2003; Neville, KR and Haberly, LB, 2003; Lagier, S. и др. , 2004). Активность митральных клеток синхронизируется с потенциалами поля MOB с частотой γ, а также с ГАМКергическим синаптическим входом (Lagier, S. et al. , 2004). Продолжительность синаптического входа митральной клетки в GC происходит в течение примерно половины цикла гамма-колебания.Гамма-осцилляторная активность митральных клеток подавляется или ослабляется антагонистами рецептора глутамата (уменьшая возбуждение GC митральными / пучковыми клетками) или антагонистами рецептора GABA A (блокируя GC-опосредованное ингибирование митральных / пучковых клеток) (Friedman, D. and Строубридж, Б.В., 2003; Лагье, С. и др., , 2004). У мышей с дефицитом субъединицы рецептора GABA A β3 функциональная экспрессия рецепторов GC GABA A практически исчезла (Nusser et al., 2001). У этих мышей амплитуды миниатюрных IPSP митральных / пучковых клеток, колебания тета-частоты и колебания γ-частоты были увеличены, а различение близкородственных смесей спиртов после обучения было плохим по сравнению с нормальными мышами. Гамма-колебания, по-видимому, также связаны с электрическими синапсами, поскольку они подавляются ингибиторами щелевых соединений (Friedman, D.
et al. , 2004). Продолжительность синаптического входа митральной клетки в GC происходит в течение примерно половины цикла гамма-колебания.Гамма-осцилляторная активность митральных клеток подавляется или ослабляется антагонистами рецептора глутамата (уменьшая возбуждение GC митральными / пучковыми клетками) или антагонистами рецептора GABA A (блокируя GC-опосредованное ингибирование митральных / пучковых клеток) (Friedman, D. and Строубридж, Б.В., 2003; Лагье, С. и др., , 2004). У мышей с дефицитом субъединицы рецептора GABA A β3 функциональная экспрессия рецепторов GC GABA A практически исчезла (Nusser et al., 2001). У этих мышей амплитуды миниатюрных IPSP митральных / пучковых клеток, колебания тета-частоты и колебания γ-частоты были увеличены, а различение близкородственных смесей спиртов после обучения было плохим по сравнению с нормальными мышами. Гамма-колебания, по-видимому, также связаны с электрическими синапсами, поскольку они подавляются ингибиторами щелевых соединений (Friedman, D.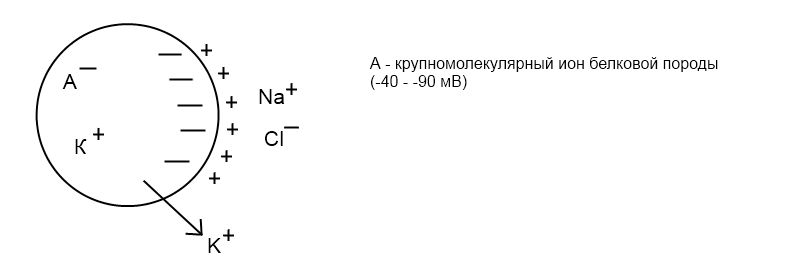 and Strowbridge, B. W., 2003). В соответствии с этим у мышей с нокаутом коннексина 36 нарушаются гамма-колебания (Hormuzdi, S.G. et al. , 2001). Внутренние мембранные свойства митральных клеток могут способствовать гамма-колебательной активности и выбросам. При приближении к порогу спайка митральные клетки демонстрируют внутренне генерируемые подпороговые колебания мембранного потенциала в диапазоне γ-частот (Chen, W. R. и Shepherd, G. M., 1997; Desmaisons, D. et al. , 1999).
and Strowbridge, B. W., 2003). В соответствии с этим у мышей с нокаутом коннексина 36 нарушаются гамма-колебания (Hormuzdi, S.G. et al. , 2001). Внутренние мембранные свойства митральных клеток могут способствовать гамма-колебательной активности и выбросам. При приближении к порогу спайка митральные клетки демонстрируют внутренне генерируемые подпороговые колебания мембранного потенциала в диапазоне γ-частот (Chen, W. R. и Shepherd, G. M., 1997; Desmaisons, D. et al. , 1999).
Калиевые токи A-типа, активные при подпороговых потенциалах в клетках Пуркинье мозжечка мыши
Подпороговая активация
I K (A) в клетках Пуркинье
Основным выводом этого исследования является то, что ПК мозжечка в возрасте 3 лет –9 постнатальных дней, обладают подпорогом активации I K (A) , который достигает пиковых амплитуд, которые при +20 мВ находятся в диапазоне от 3 до 7 нА.Тот факт, что эти токи составляют около одной трети от общего напряжения зависимого, Ca 2+ -независимого тока K + этих клеток, предполагает, что они участвуют в соответствующих физиологических функциях. Этот результат является неожиданным, поскольку ПК также имеют большие токи K + , чувствительные к TEA (Raman & Bean, 1999; Martina et al. 2001) и большое количество Ca 2+ -зависимых каналов K + . (Jacquin & Gruol 1999; Raman & Bean, 1999).Более того, нечувствительные к TEA I K (A) отсутствуют в патчах вне сомы ПК (Southan & Robertson, 2000) и в диссоциированных ПК (Nam & Hockberger, 1997; Raman & Bean, 1999). Отсутствие I K (A) в изолированных ПК может быть связано с обрезкой дендритного дерева или аксона или с другими изменениями, вызванными процедурой диссоциации. Вместе с отсутствием I K (A) в соматических участках (Southan & Robertson, 2000), это позволяет предположить, что каналы K + , ответственные за подпороговое состояние I K (A) , в основном расположены в дендритах или в аксонах ПК.Недавние открытия указывают на присутствие в дендритах ПК релевантного TEA-чувствительного I K (A) , активирующегося при высоком напряжении, что, вероятно, связано с каналами подсемейства Kv3 (Martina et al.
Этот результат является неожиданным, поскольку ПК также имеют большие токи K + , чувствительные к TEA (Raman & Bean, 1999; Martina et al. 2001) и большое количество Ca 2+ -зависимых каналов K + . (Jacquin & Gruol 1999; Raman & Bean, 1999).Более того, нечувствительные к TEA I K (A) отсутствуют в патчах вне сомы ПК (Southan & Robertson, 2000) и в диссоциированных ПК (Nam & Hockberger, 1997; Raman & Bean, 1999). Отсутствие I K (A) в изолированных ПК может быть связано с обрезкой дендритного дерева или аксона или с другими изменениями, вызванными процедурой диссоциации. Вместе с отсутствием I K (A) в соматических участках (Southan & Robertson, 2000), это позволяет предположить, что каналы K + , ответственные за подпороговое состояние I K (A) , в основном расположены в дендритах или в аксонах ПК.Недавние открытия указывают на присутствие в дендритах ПК релевантного TEA-чувствительного I K (A) , активирующегося при высоком напряжении, что, вероятно, связано с каналами подсемейства Kv3 (Martina et al.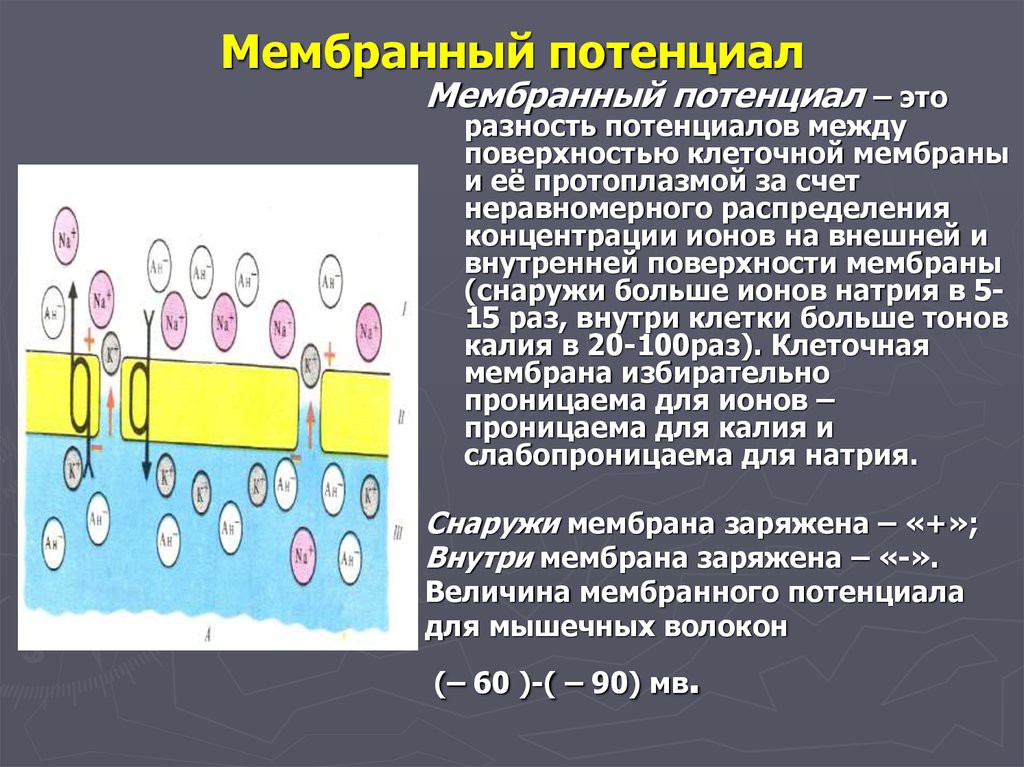 2001). Различные свойства этого тока относительно нечувствительного к ТЭА подпорогового активированного I K (A) , описанного в этой статье, предполагают, что эти токи могут иметь дополнительные функциональные роли. В ПК, токи I K (A) участвовали в ускорении возбуждения, вызванном введением деполяризующего тока (Hounsgaard & Midtgaard, 1988), в дендритных пиках Ca 2+ (Etzion & Grossman, 1998). ) и в контроле повышения [Ca 2+ ] и в дендритах (Midtgaard et al. 1993). Кривые активации и инактивации, описанные в нашем исследовании, показывают, что I K (A) , вероятно, также участвует в контроле подпороговых изменений мембранного потенциала. Это свойство также предполагает, что синаптические сигналы могут фильтроваться и модифицироваться этим током, как было показано в пирамидных нейронах гиппокампа, в которых нечувствительные к дендротоксину токи K + A-типа в большом количестве присутствуют в дендритах, где они могут действовать как фильтр верхних частот входящих синаптических сигналов (Hoffman et al.
2001). Различные свойства этого тока относительно нечувствительного к ТЭА подпорогового активированного I K (A) , описанного в этой статье, предполагают, что эти токи могут иметь дополнительные функциональные роли. В ПК, токи I K (A) участвовали в ускорении возбуждения, вызванном введением деполяризующего тока (Hounsgaard & Midtgaard, 1988), в дендритных пиках Ca 2+ (Etzion & Grossman, 1998). ) и в контроле повышения [Ca 2+ ] и в дендритах (Midtgaard et al. 1993). Кривые активации и инактивации, описанные в нашем исследовании, показывают, что I K (A) , вероятно, также участвует в контроле подпороговых изменений мембранного потенциала. Это свойство также предполагает, что синаптические сигналы могут фильтроваться и модифицироваться этим током, как было показано в пирамидных нейронах гиппокампа, в которых нечувствительные к дендротоксину токи K + A-типа в большом количестве присутствуют в дендритах, где они могут действовать как фильтр верхних частот входящих синаптических сигналов (Hoffman et al. 1997; Johnston et al. 2000).
1997; Johnston et al. 2000).
В нашем исследовании компонент A-типа токов K + был впервые выделен с помощью TEA в концентрации, которая не оказывает существенного влияния на каналы Kv1 или Kv4, за исключением высокочувствительной субъединицы Kv1.1. к этому лекарству с K D 0,3 мм (Coetzee et al. 1999). ПК также экспрессируют некоторые инактивирующие высокопороговые субъединицы канала K + подсемейства Kv3 (Weiser et al. 1994; Goldmann-Wohl et al. 1994), но они очень чувствительны к TEA, так что мы можем исключить вклад каналов Kv3 в наших условиях. PC также экспрессируют субъединицы Kv2.1 и Kv2.2 (Hwang et al. 1993), которые ответственны за медленную активацию и инактивацию запаздывающих выпрямительных токов, которые умеренно чувствительны к TEA (Coetzee et al. 1999). Следовательно, возможно, что медленно отключающиеся токи, зарегистрированные в TEA, как от -80, так и от -40 мВ удерживающих потенциалов, частично связаны с каналами Kv2. Чтобы исключить вклад этого тока в распад I K (A) , мы использовали протокол вычитания (). Следовательно, подпороговое значение I K (A) , описанное в этом исследовании, можно предположительно отнести к каналам подсемейства Kv1, которые не блокируются в значительной степени TEA, и / или к каналам подсемейства Kv4. ПК экспрессируют несколько субъединиц Kv1α (Veh et al. 1995; Verma-Kurvari et al. 1997; Sacco & Tempia, 2000; Chung et al. 2001), которые вместе с субъединицами Kvβ обладают свойствами, совместимыми с зарегистрированным нами I K (A) (Coetzee et al. 1999; Pongs et al. 1999). Фактически, в возрасте клеток, использованных в этом отчете, мышиные ПК экспрессируют Kvβ1 (Butler et al. 1998), тогда как у взрослых крыс они экспрессируют субъединицы Kvβ1 и Kvβ2 (Rhodes et al. 1996). Кроме того, ПК также экспрессируют Kv4.3 (Serodio & Rudy, 1998), который также нечувствителен к концентрации 4 мМ ТЕА, использованной в нашем исследовании (Dixon et al.
Чтобы исключить вклад этого тока в распад I K (A) , мы использовали протокол вычитания (). Следовательно, подпороговое значение I K (A) , описанное в этом исследовании, можно предположительно отнести к каналам подсемейства Kv1, которые не блокируются в значительной степени TEA, и / или к каналам подсемейства Kv4. ПК экспрессируют несколько субъединиц Kv1α (Veh et al. 1995; Verma-Kurvari et al. 1997; Sacco & Tempia, 2000; Chung et al. 2001), которые вместе с субъединицами Kvβ обладают свойствами, совместимыми с зарегистрированным нами I K (A) (Coetzee et al. 1999; Pongs et al. 1999). Фактически, в возрасте клеток, использованных в этом отчете, мышиные ПК экспрессируют Kvβ1 (Butler et al. 1998), тогда как у взрослых крыс они экспрессируют субъединицы Kvβ1 и Kvβ2 (Rhodes et al. 1996). Кроме того, ПК также экспрессируют Kv4.3 (Serodio & Rudy, 1998), который также нечувствителен к концентрации 4 мМ ТЕА, использованной в нашем исследовании (Dixon et al.![]() 1996).
1996).
Подсемейства Kv1 и Kv4 можно различить по некоторым биофизическим свойствам. Каналы, образованные субъединицами Kv1, восстанавливаются после инактивации довольно медленно, с постоянными времени в несколько секунд и инактивируются с постоянными времени, которые становятся быстрее при более деполяризованных напряжениях (Coetzee et al. 1999). Добавление субъединиц Kvβ дополнительно продлевает восстановление после инактивации и во многих случаях вызывает или усиливает процесс инактивации, который поддерживает зависимость от напряжения с более быстрым спадом при более деполяризованных потенциалах (Pongs et al. 1999). Единственным исключением является Kv1.4, постоянная времени инактивации которого лишь незначительно зависит от напряжения, особенно при положительных потенциалах (McIntosh et al. 1997). Однако при отрицательных потенциалах Kv1.4 имеет более сильную зависимость от напряжения, либо экспрессируется отдельно, либо совместно с Kvβ1 или Kvβ2 (McIntosh et al. 1997).
1997).
Напротив, каналы, образованные субъединицами Kv4, быстро восстанавливаются после инактивации (Serodio et al. 1994, 1996), и их скорость инактивации очень слабо зависит от напряжения (Serodio et al. 1994, 1996). Добавление вспомогательных субъединиц Kv4 (KChIP1, KChIP2, KChIP3) дополнительно увеличивает восстановление после инактивации и замедляет инактивацию, но не влияет на отсутствие зависимости постоянных времени инактивации от напряжения (An et al. 2000). Модель I K (A) , описанная в этом исследовании, восстановлена после инактивации с двойным экспоненциальным изменением во времени с постоянными времени около 60 мс и 1 с. Более быстрая постоянная времени соответствует восстановлению каналов Kv4 (Serodio et al. 1994, 1996; An et al. 2000), хотя он по крайней мере на порядок короче, чем в каналах Kv1 (Coetzee et al. 1999; Pongs et al. 1999). Более длинная постоянная времени соответствует восстановлению канала Kv1 (Coetzee et al. 1999; Pongs et al. 1999), но она явно больше, чем в каналах Kv4 (Serodio et al. 1994,1996; An и др., , 2000). Возможное объяснение состоит в том, что более быстрая постоянная времени отражает активность каналов, образованных субъединицами Kv4, в то время как более медленная постоянная времени отражает активность каналов, образованных субъединицами Kv1.В этом отношении 78% вклада более быстрого компонента по сравнению с 22% вклада более медленного предполагает, что каналы Kv4 ответственны за большую часть подпорога I K (A) , чем каналы Kv1. Анализ кинетики инактивации, по-видимому, подтверждает эту интерпретацию, поскольку зависимость обеих постоянных времени затухания от напряжения очень слабая при всех напряжениях, как сообщалось для каналов Kv4, включая Kv4.3, который выражается ПК (Serodio et al. 1994, 1996; An et al. 2000; Beck et al. 2002). Более того, зависимость от напряжения относительных вкладов быстрых и медленных компонентов идентична той, которая описана для субъединицы Kv4.
1999; Pongs et al. 1999), но она явно больше, чем в каналах Kv4 (Serodio et al. 1994,1996; An и др., , 2000). Возможное объяснение состоит в том, что более быстрая постоянная времени отражает активность каналов, образованных субъединицами Kv4, в то время как более медленная постоянная времени отражает активность каналов, образованных субъединицами Kv1.В этом отношении 78% вклада более быстрого компонента по сравнению с 22% вклада более медленного предполагает, что каналы Kv4 ответственны за большую часть подпорога I K (A) , чем каналы Kv1. Анализ кинетики инактивации, по-видимому, подтверждает эту интерпретацию, поскольку зависимость обеих постоянных времени затухания от напряжения очень слабая при всех напряжениях, как сообщалось для каналов Kv4, включая Kv4.3, который выражается ПК (Serodio et al. 1994, 1996; An et al. 2000; Beck et al. 2002). Более того, зависимость от напряжения относительных вкладов быстрых и медленных компонентов идентична той, которая описана для субъединицы Kv4. 3, коэкспрессируемой с вспомогательной субъединицей KChIP1 (Beck et al. 2002). Кинетика активации как Kv1.4, так и Kv4.3 показывает зависимость напряжения, аналогичную подпороговой I K (A) ПК. Kv1.4, особенно когда он коэкспрессируется с субъединицей Kvβ, имеет быструю постоянную времени активации в диапазоне подпороговых значений I K (A) ПК (McIntosh et al. 1997). Напротив, Kv4.3, даже когда присутствует субъединица KChIP1, имеет большее время до пика (Beck et al. 2002). Взятые вместе, биофизические свойства подпорога I K (A) ПК предполагают, что основным детерминантом этого тока являются каналы Kv4, наиболее вероятно Kv4.3, с незначительным, но значительным вкладом каналов Kv1.
3, коэкспрессируемой с вспомогательной субъединицей KChIP1 (Beck et al. 2002). Кинетика активации как Kv1.4, так и Kv4.3 показывает зависимость напряжения, аналогичную подпороговой I K (A) ПК. Kv1.4, особенно когда он коэкспрессируется с субъединицей Kvβ, имеет быструю постоянную времени активации в диапазоне подпороговых значений I K (A) ПК (McIntosh et al. 1997). Напротив, Kv4.3, даже когда присутствует субъединица KChIP1, имеет большее время до пика (Beck et al. 2002). Взятые вместе, биофизические свойства подпорога I K (A) ПК предполагают, что основным детерминантом этого тока являются каналы Kv4, наиболее вероятно Kv4.3, с незначительным, но значительным вкладом каналов Kv1.
Однако есть несколько известных условий, которые могут изменять биофизические свойства токов в нативном нейроне, таком как ПК в срезах ткани.Например, могут присутствовать новые или непроверенные каналообразующие или вспомогательные субъединицы, или канал может находиться в различных фосфорилированных или окислительно-восстановительных состояниях с разными свойствами, или на него могут влиять взаимодействующие белки (см. Hille, 2001). По этим причинам также важно использовать другой независимый подход, такой как использование селективных блокаторов подсемейства, чтобы оценить, являются ли каналы Kv1 и / или Kv4 ответственными за подпороговое значение I K (A) .
Было показано, что агитоксин-2 блокирует гомомерные каналы, образованные Kv1.1, 1,3 или 1,6 экспрессируется в ооцитах Xenopus со значениями K d , равными 44, 4 и 37 пм соответственно (Garcia et al. 1994). Эффективность этого блокатора подтверждена также на гладкомышечных клетках мышей, которые экспрессируют субъединицу Kv1.6, но не имеют Kv1.5 (Cheong et al. 2001 b ). Интересно, что недавние данные показывают, что в клетках, которые экспрессируют Kv1.6, а также Kv1.5, токи K + не чувствительны к AgTX-2 (Cheong et al. 2001 а ). Эффективность AgTX-2 в ПК мышей согласуется с отсутствием экспрессии Kv1.5 в мозжечке крыс (Koch et al. 1997). Оба данных противоречат результатам иммуногистохимического исследования мозжечка крыс, где ПК были помечены как анти-Kv1.5, так и анти-Kv1.6 антителами (Chung et al. 2001). Однако следует проявлять осторожность при сравнении наших результатов, полученных на ПК с неполовозрелыми мышами, с исследованиями, проведенными на взрослых крысах. Фактически, предварительные результаты предполагают, что неполовозрелые ПК мышей (постнатальный возраст: P7) экспрессируют более расширенный набор субъединиц Kv1, чем взрослые мыши или взрослые крысы (Sacco & Tempia, 2000), открывая возможность образования гетеромерных каналов Kv1, которые могли бы при хотя бы частично, объяснить инактивационные свойства AgTX-2-чувствительного компонента I K (A) .
Флекаинид не является селективным для субъединиц Kv4, но имеет более высокое сродство к этому подсемейству (Dixon et al. 1996; Yeola & Snyders, 1997), чем к каналам Kv1 (Grissmer et al. 1994; Yamagishi et al. 1995; Yeola & Snyders, 1997). Тот факт, что блок флекаинида с более высоким сродством составляет около половины I K (A) , в то время как блок AgTX-2 составляет всего около 20%, предполагает, что вклад каналов Kv4 больше, чем вклад каналов Kv4. Каналы Kv1.Взятые вместе, эти данные предполагают, что клетки Пуркинье имеют большие I K (A) , активные при подпороговых потенциалах, которые обусловлены исключительно каналами, чувствительными как к 4-AP, так и к флекаиниду, и что основной компонент подпорогового I K (A) связан с каналами подсемейства Kv4 с меньшим вкладом Kv1.
Физиологическое значение
A-тип K + токи, активные при подпороговых потенциалах в нейронах, участвуют в нескольких физиологических ролях, включая затухание синаптических возбуждающих сигналов в дендритах пирамидных нейронов (Hoffman et al. 1997) и модуляцию дендритных транзиентов [Ca 2+ ] – в ПК мозжечка (Midtgaard et al. 1993). Описываемый нами образец I K (A) обладает биофизическими свойствами, необходимыми для контроля подпороговой дендритной возбудимости ПК. Было показано, что нанесение 4-AP на ПК в срезах значительно сокращает латентность спайков Ca 2+ и в то же время повышает уровень внутридендритных [Ca 2+ ] и (Midtgaard ). и другие. 1993). Эти эффекты имеют важные последствия для процессов, запускаемых Ca 2+ , таких как индукция параллельных волокон (Ito, 1991; Linden & Connor, 1995; Daniel et al. 1998) и длительная депрессия лазания по волокнам (Hansel & Linden , 2000) и «потенцирования отскока» ГАМКергических синапсов (Kano et al. 1992). Однако недавнее открытие высокого порога I K (A) в дендритах ПК из-за каналов подсемейства Kv3 (Martina et al. 2001), которые блокируются низкими концентрациями 4-АР (Coetzee et al. 1999), могут объяснить эффекты на скачки Ca 2+ и кратковременные повышения [Ca 2+ ] и (Midtgaard и др., , 1993). В этом случае роль I K (A) , описанного в этой статье, если показано, что он локализован на дендритах ПК, будет заключаться в специфическом подавлении подпороговых возбуждающих входных сигналов.
Обнаружение того, что большой компонент подпорога I K (A) высокочувствителен к флекаиниду, предполагая, что это связано с каналами Kv4, согласуется с результатами, полученными для других типов клеток.Например, каналы K + подсемейства Kv4 являются основным фактором, определяющим кратковременный исходящий ток сердечных миоцитов (Dixon et al. 1996; Johns et al. 1997; Barry et al. 1998). , в то время как оставшийся компонент почти полностью связан с каналом Kv1.4 (Guo et al. 2000). Также было показано, что инактивирующий дендритный ток K + пирамидных нейронов определяется каналами, нечувствительными к блокатору подсемейства Kv1, поэтому он предположительно был отнесен к подсемейству Kv4 (Hoffman et al. 1997). Более того, в симпатических нейронах верхнего шейного ганглия каналы Kv4 являются основным компонентом I K (A) и играют важную роль в контроле возбудимости мембран и паттерна возбуждения, но не в реполяризации потенциала действия (Малин И Нербон, 2000). Также у ПК мозжечка мы не обнаружили влияния на физиологические потенциалы действия, если только они не были пролонгированы применением ТЭА.
Влияние блокаторов подпороговых значений I K (A) на гиперполяризацию, которая развивается сразу после этапа деполяризации, указывает на то, что каналы, ответственные за этот ток, значительно открываются во время активации вызванных клеток.Деактивация при потенциале приблизительно -70 мВ, при котором ячейка поддерживалась, имела постоянную времени 3,4 мс, что позволяло напряжению примерно 10 мс превысить такой потенциал и вызвать гиперполяризацию. В некоторых условиях ПК демонстрируют длительную деполяризацию, называемую потенциалами плато, как in vitro, (Llinas & Sugimori, 1980, , , b, ), так и in vivo, (Ekerot & Oscarsson, 1981), которые имеют некоторое сходство с ступенчатые деполяризации, использованные в этом исследовании.Потенциалы плато часто следуют за восходящей активностью волокон (Ekerot & Oscarsson, 1981) и могут длиться более 1 секунды. Следовательно, возможно, что физиологическая роль подпорога I K (A) заключается в том, чтобы участвовать в прекращении потенциалов плато.
Анализ спонтанного или вызванного возбуждения у молодых животных очень затруднен, поскольку характер незрелых выделений сильно различается между PC (Woodward et al. 1969; Crepel, 1972).Оценку роли подпорога I K (A) следует проводить на более зрелых или взрослых ПК, свойства разряда которых изучены подробно (см. Обзор Ito, 1984). Фактически, для ПК в срезах мозжечка взрослой черепахи было высказано предположение, что I K (A) отвечает за ускорение спайков (Hounsgaard & Midtgaard, 1988). Это открытие подразумевает роль I K (A) в формировании паттерна возбуждения ПК с последствиями для сигналов, которые покидают кору мозжечка и вносят вклад в моторный контроль (Ito, 1984).Чтобы воздействовать на схему возбуждения, необязательно, чтобы каналы, отвечающие за подпороговое значение I K (A) , располагались в дендритах. Расположение в соме или на бугре аксона было бы более благоприятным для такого рода роли. Следовательно, наши настоящие данные оставляют открытым вопрос о локализации предполагаемых каналов Kv1 и Kv4, которые лежат в основе подпорога I K (A) ПК мозжечка. В нейронах верхнего шейного ганглия Kv4.3, как было показано, локализуется в дистальных дендритных ветвях (Malin & Nerbonne, 2000), а другие члены подсемейства Kv4, как было показано, экспрессируются в дендритном компартменте (Song et al. 1998). Напротив, в большинстве типов нейронов Kv1.4 экспрессируется в аксоне (Sheng et al. 1992; Song et al. 1998; Arnold & Clapham, 1999). Заметным исключением являются нейроны дорсального ядра улитки, где Kv1.4 локализован в дендритах и дендритных шипах (Juiz et al. 2000). Следовательно, возможно, что в ПК мозжечка большая часть подпорога I K (A) , из-за каналов Kv4, расположена в дендритах, тогда как меньший компонент Kv1 расположен в аксоне.
Подпорогово-активный ток KV7 регулирует нейротрансмиссию, ограничивая спайк-индуцированный приток Ca2 + в синаптические терминалы гиппокампа, покрытые мхом.
Schneggenburger, R. & Rosenmund, C. выпуск. Nat. Neurosci. 18 , 935–941 (2015).
CAS
Статья
Google Scholar
Kaeser, P. S. & Regehr, W. G. Молекулярные механизмы синхронного, асинхронного и спонтанного высвобождения нейротрансмиттеров. Annu. Rev. Physiol. 76 , 333–363 (2014).
CAS
Статья
Google Scholar
Кавалали, Э.Т. Механизмы и функции спонтанного выброса нейромедиаторов. Nat. Rev. Neurosci. 16 , 5–16 (2015).
CAS
Статья
Google Scholar
Триммер, Дж. С. Субклеточная локализация каналов K + в нейронах мозга млекопитающих: исключительная точность среди необычайной сложности. Нейрон 85 , 238–256 (2015).
CAS
Статья
Google Scholar
Хоппа, М. Б., Гузер, Г., Армбрустер, М. и Райан, Т. А. Контроль и пластичность формы волны пресинаптического потенциала действия на малых нервных окончаниях ЦНС. Нейрон 84 , 778–789 (2014).
CAS
Статья
Google Scholar
Foust, A. J., Yu, Y., Popovic, M., Zecevic, D. & McCormick, D. A. Соматический мембранный потенциал и каналы Kv1 контролируют реполяризацию спайков в коллатералях кортикальных аксонов и пресинаптических бутонах. J. Neurosci. 31 , 15490–15498 (2011).
CAS
Статья
Google Scholar
Бегум Р., Бакири Ю., Волынски К. Э. и Куллманн Д. М. Расширение потенциала действия при пресинаптической каннелопатии. Nat. Commun. 7 , 12102 (2016).
CAS
Статья
Google Scholar
Накамура Ю. и Такахаши Т.Изменения в развитии калиевых токов в чашечке пресинаптического терминала Held крысы. J. Physiol. 581 , 1101–1112 (2007).
CAS
Статья
Google Scholar
Ishikawa, T. et al. Различная роль калиевых каналов Kv1 и Kv3 в чашечке пресинаптического терминала Held. Дж. Neurosci . 23 , 10445–10453 (2003).
CAS
Статья
Google Scholar
Бишофбергер, Дж., Энгель, Д., Фротшер, М. и Йонас, П. Время и эффективность высвобождения передатчика в синапсах мшистых волокон в сети гиппокампа. Pflugers Arch. 453 , 361–372 (2006).
CAS
Статья
Google Scholar
Dodson, P. D. et al. Пресинаптические каналы Kv1.2 крысы подавляют синаптическую терминальную гипервозбудимость после инвазии потенциала действия. J. Physiol. 550 , 27–33 (2003).
CAS
Статья
Google Scholar
Саутан, А. П. и Робертсон, Б. Электрофизиологическая характеристика потенциал-зависимых K (+) токов в корзине мозжечка и клетках Пуркинье: подсемейства каналов Kv1 и Kv3 присутствуют в нервных окончаниях корзинчатых клеток. J. Neurosci. 20 , 114–122 (2000).
CAS
Статья
Google Scholar
Алле, Х., Кубота, Х. и Гейгер, Дж. Р. Редкие, но высокоэффективные Kv3 опережают каналы BKCa в реполяризации потенциала действия в бутонах из мшистых волокон гиппокампа. J. Neurosci. 31 , 8001–8012 (2011).
CAS
Статья
Google Scholar
Гейгер, Дж. Р. и Джонас, П. Динамический контроль пресинаптического притока Са 2+ путем быстрой инактивации каналов K + в бутонах из мшистых волокон гиппокампа. Нейрон 28 , 927–939 (2000).
CAS
Статья
Google Scholar
Кавагути, С. Ю. и Сакаба, Т. Контроль тормозных синаптических выходов за счет низкой возбудимости окончаний аксонов, выявленной при прямой записи. Нейрон 85 , 1273–1288 (2015).
CAS
Статья
Google Scholar
Ян, Ю.М.и другие. Повышение точности нейротрансмиссии за счет зависимого от активности облегчения пресинаптических калиевых потоков. Nat. Commun. 5 , 4564 (2014).
CAS
Статья
Google Scholar
Лай, Х. К. и Ян, Л. Ю. Распределение и нацеливание нейронных потенциалзависимых ионных каналов. Nat. Ред. 7 , 548–562 (2006).
CAS
Статья
Google Scholar
Браун, Д. А. и Пассмор, Г. М. Нейронные каналы KCNQ (Kv7). руб. J. Pharmacol. 156 , 1185–1195 (2009).
CAS
Статья
Google Scholar
Jentsch, T. J. Нейрональные калиевые каналы KCNQ: физиология и роль в заболевании. Nat. Rev. Neuerosci 1 , 21–30 (2000).
CAS
Статья
Google Scholar
Debanne, D., Campanac, E., Bialowas, A., Carlier, E., Alcaraz, G., физиология Axon. Physiol. Ред. 91 , 555–602 (2011).
CAS
Статья
Google Scholar
Коле, М. Х. и Стюарт, Г. Дж. Обработка сигналов в начальном сегменте аксона. Нейрон 73 , 235–247 (2012).
CAS
Статья
Google Scholar
Дево, Дж. Дж., Клеопа, К. А., Купер, Э. С. и Шерер, С. С. KCNQ2 представляет собой узловой K + канал. J. Neurosci. 24 , 1236–1244 (2004).
CAS
Статья
Google Scholar
Pan, Z. et al. Общий механизм на основе анкирина-G сохраняет каналы KCNQ и NaV в электрически активных доменах аксона. J. Neurosci. 26 , 2599–2613 (2006).
CAS
Статья
Google Scholar
Huang, H. & Trussell, L.O. Каналы KCNQ5 управляют свойствами покоя и вероятностью высвобождения синапса. Nat. Neurosci. 14 , 840–847 (2011).
CAS
Статья
Google Scholar
Fidzinski, P. et al. Каналы KCNQ5 K + контролируют синаптическое торможение в гиппокампе и быстрые сетевые колебания. Nat. Commun. 6 , 6254 (2015).
CAS
Статья
Google Scholar
Cooper, E. C., Harrington, E., Jan, Y. N. & Jan, L. Y. Субъединицы KCNQ2 канала M локализованы в ключевых местах для контроля колебаний и синхронизации нейронной сети в мозге мыши. J. Neurosci. 21 , 9529–9540 (2001).
CAS
Статья
Google Scholar
Гейгер, Дж., Вебер, Ю. Г., Ландвермейер, Б., Соммер, К. и Лерче, Х. Иммуногистохимический анализ калиевых каналов KCNQ3 в мозге мыши. Neurosci. Lett. 400 , 101–104 (2006).
CAS
Статья
Google Scholar
Weber, Y.G. et al. Иммуногистохимический анализ калиевых каналов KCNQ2 в мозге взрослых и развивающихся мышей. Brain Res. 1077 , 1–6 (2006).
CAS
Статья
Google Scholar
Acsady, L. & Kali, S. Модели, структура, функция: преобразование корковых сигналов в зубчатой извилине. Прог. Brain Res. 163 , 577–599 (2007).
CAS
Статья
Google Scholar
Хенце Д. А., Урбан Н. Н. и Баррионуево Г. Многообразный путь мшистых волокон гиппокампа: обзор. Неврология 98 , 407–427 (2000).
CAS
Статья
Google Scholar
Яффе, Д. Б. и Гутьеррес, Р.Синаптическая передача по мшистым волокнам: сообщение от зубчатой извилины к области CA3. Прог. Brain Res. 163 , 109–132 (2007).
CAS
Статья
Google Scholar
Pelkey, K. A. & McBain, C.J. Зависимая от клеток-мишеней пластичность в цепи замшелых волокон-CA3 обнаруживает разделенную регуляцию пресинаптической функции в дивергентных сайтах высвобождения. J. Physiol. 586 , 1495–1502 (2008).
CAS
Статья
Google Scholar
Ребола, Н., Карта, М. и Мюлле, С. Функционирование и пластичность схем гиппокампа CA3: последствия для кодирования памяти. Nat. Rev. Neurosci. 18 , 208–220 (2017).
CAS
Статья
Google Scholar
Леао, Р. Н., Тан, Х. М. и Фисан, А. Каналы Kv7 / KCNQ контролируют фазировку потенциала действия пирамидных нейронов во время гамма-колебаний гиппокампа in vitro. J. Neurosci. 29 , 13353–13364 (2009).
CAS
Статья
Google Scholar
Петерс, Х.С., Ху, Х., Понгс, О., Сторм, Дж. Ф. и Исбрандт, Д. Условное трансгенное подавление М-каналов в мозге мыши выявляет функции нейрональной возбудимости, резонанса и поведения. Nat. Neurosci. 8 , 51–60 (2005).
CAS
Статья
Google Scholar
Singh, N.A. et al. Мышиные модели мутаций KCNQ2 и KCNQ3 человека при доброкачественных семейных неонатальных судорогах демонстрируют судороги и пластичность нейронов без синаптической реорганизации. J. Physiol. 586 , 3405–3423 (2008).
CAS
Статья
Google Scholar
Delvendahl, I., Weyhersmuller, A., Ritzau-Jost, A. & Hallermann, S. Бутоны из мшистых волокон гиппокампа и мозжечка — одно и то же название, различное назначение. J. Physiol. 591 , 3179–3188 (2013).
CAS
Статья
Google Scholar
Браун Д. А. и Адамс П. Р. Мускариновое подавление нового чувствительного к напряжению тока K + в нейроне позвоночных. Nature 283 , 673–676 (1980).
CAS
Статья
Google Scholar
Шах, М. М., Мильоре, М., Валенсия, И., Купер, Э. С. и Браун, Д. А. Функциональное значение аксональных каналов Kv7 в пирамидных нейронах гиппокампа. Proc. Natl. Акад. Sci. США 105 , 7869–7874 (2008).
CAS
Статья
Google Scholar
Martinello, K. et al. Холинергическая афферентная стимуляция вызывает пластичность функции аксонов в гранулярных клетках гиппокампа взрослых. Нейрон 85 , 346–363 (2015).
CAS
Статья
Google Scholar
Wang, H. S. et al. Субъединицы калиевых каналов KCNQ2 и KCNQ3: молекулярные корреляты М-канала. Наука 282 , 1890–1893 (1998).
CAS
Статья
Google Scholar
Young, C.C. et al. Повышение регуляции внутренних выпрямительных каналов K + (Kir2) в гранулярных клетках зубчатой извилины при височной эпилепсии. J. Physiol. 587 , 4213–4233 (2009).
CAS
Статья
Google Scholar
Gonzalez, J.C., Epps, S.A., Markwardt, S.J., Wadiche, J. I. & Overstreet-Wadiche, L. Конститутивная и синаптическая активация каналов GIRK дифференцирует зрелые и новорожденные зубчатые гранулярные клетки. J. Neurosci. 38 , 6513–6526 (2018).
CAS
Статья
Google Scholar
Энгель, Д. и Йонас, П. Усиление пресинаптического потенциала действия с помощью потенциал-управляемых Na + -каналов в бутонах из мшистых волокон гиппокампа. Нейрон 45 , 405–417 (2005).
CAS
Статья
Google Scholar
Vyleta, N.P. и Jonas, P. Слабая связь между Ca2 + -каналами и датчиками высвобождения в пластиковом синапсе гиппокампа. Наука 343 , 665–670 (2014).
CAS
Статья
Google Scholar
Матеос-Апарисио, П., Мерфи, Р. и Сторм, Дж. Ф. Дополнительные функции калиевых каналов SK и Kv7 / M в контроле возбудимости и синаптической интеграции в зубчатых гранулярных клетках гиппокампа крысы. J. Physiol. 592 , 669–693 (2014).
CAS
Статья
Google Scholar
Джексон, М. Б. и Редман, С. Дж. Динамика кальция, буферизация и насыщение буфера в бутонах зубчатых аксонов гранулярных клеток в воротах ворот. J. Neurosci. 23 , 1612–1621 (2003).
CAS
Статья
Google Scholar
Скотт Р. и Русаков Д. А. Основные детерминанты пресинаптической динамики Ca 2+ в индивидуальных синапсах пирамидных клеток мшистых волокон и CA3. J. Neurosci. 26 , 7071–7081 (2006).
CAS
Статья
Google Scholar
Чемберленд, С., Тимофеева, Ю., Евстратова, А., Волынски, К., Тот, К. Подсчет потенциала действия на гигантских замшелых волоконно-оптических терминалах, передающих информацию в гиппокампе. Proc. Natl. Акад. Sci. США 115 , 7434–7439 (2018).
CAS
Статья
Google Scholar
Бишофбергер, Дж., Гейгер, Дж. Р. и Йонас, П. Время и эффективность активации канала Са 2+ в бутонах из мшистых волокон гиппокампа. J. Neurosci. 22 , 10593–10602 (2002).
CAS
Статья
Google Scholar
Li, L., Bischofberger, J. & Jonas, P. Дифференциальное стробирование и рекрутирование каналов Ca 2+ P / Q-, N- и R-типов в бутонах из мшистых волокон гиппокампа. J. Neurosci. 27 , 13420–13429 (2007).
CAS
Статья
Google Scholar
Holderith, N. et al. Вероятность высвобождения глутаматергических окончаний гиппокампа зависит от размера активной зоны. Nat. Neurosci. 15 , 988–997 (2012).
CAS
Статья
Google Scholar
Кастильо, П. Э., Вайскопф, М. Г. и Николл, Р. А. Роль каналов Ca 2+ в синаптической передаче гиппокампа, покрытой мхом, и долгосрочном потенцировании. Neuron 12 , 261–269 (1994).
CAS
Статья
Google Scholar
Джонс, Р. Т., Фаас, Г. К. и Моди, И. Внутриклеточный бикарбонат регулирует генерацию потенциала действия посредством модуляции канала KCNQ. J. Neurosci. 34 , 4409–4417 (2014).
Артикул
Google Scholar
Pernia-Andrade, A. J. & Jonas, P. Модулируемые тета-гамма синаптические токи в гранулярных клетках гиппокампа in vivo определяют механизм сетевых колебаний. Нейрон 81 , 140–152 (2014).
CAS
Статья
Google Scholar
Vyleta, N. P., Borges-Merjane, C. & Jonas, P. Зависимая от пластичности, полная детонация в синапсах пирамидных нейронов гиппокампа, покрытых мхом, и CA3. Elife 5 , pii: e17977 (2016).
Артикул
Google Scholar
Хенце, Д. А., Виттнер, Л.& Buzsaki, G. Отдельные гранулярные клетки надежно разряжают мишени в сети CA3 гиппокампа in vivo. Nat. Neurosci. 5 , 790–795 (2002).
CAS
Статья
Google Scholar
Battefeld, A., Tran, B.T., Gavrilis, J., Cooper, E.C. и Kole, M.H. Гетеромерные каналы Kv7.2 / 7.3 по-разному регулируют инициирование потенциала действия и проведение в миелинизированных аксонах неокортекса. Дж.Neurosci. 34 , 3719–3732 (2014).
CAS
Статья
Google Scholar
Schwarz, J. R. et al. Каналы KCNQ опосредуют IK, медленный ток K + , регулирующий возбудимость в крысином узле Ранвье. J. Physiol. 573 , 17–34 (2006).
CAS
Статья
Google Scholar
Stegen, M. et al.Адаптивная внутренняя пластичность в клетках гранул зубчатой извилины человека при височной эпилепсии. Cereb. Cortex 22 , 2087–2101 (2012).
Артикул
Google Scholar
Greene, D. L., Kang, S. & Hoshi, N. XE991 и линопирдин являются зависимыми от состояния ингибиторами каналов Kv7 / KCNQ, которые благоприятствуют активированным одиночным субъединицам. J. Pharmacol Exp Ther 362 , 177–185 (2017).
CAS
Статья
Google Scholar
Yue, C. & Yaari, Y. Аксо-соматические и апикальные дендритные каналы Kv7 / M по-разному регулируют внутреннюю возбудимость пирамидных клеток СА1 взрослых крыс. J. Neurophysiol. 95 , 3480–3495 (2006).
CAS
Статья
Google Scholar
Ху, В. и Бин, Б. П. Дифференциальный контроль аксонального и соматического потенциала покоя с помощью зависимой от напряжения проводимости в пирамидных нейронах коркового слоя 5. Neuron 97 , 1315–1326 e1313 (2018).
CAS
Статья
Google Scholar
Elmedyb, P. et al. Модуляция каналов ERG XE991. Basic Clin. Pharmacol. Toxicol. 100 , 316–322 (2007).
CAS
Статья
Google Scholar
Yue, C., Remy, S., Su, H., Beck, H. & Yaari, Y. Проксимальные персистентные каналы N a + вызывают спайк постдеполяризации и связанный с этим разрыв пирамидных клеток CA1 взрослых. J. Neurosci. 25 , 9704–9720 (2005).
CAS
Статья
Google Scholar
Vervaeke, K., Gu, N., Agdestein, C., Hu, H. & Storm, JF Kv7 / KCNQ / M-каналы в глутаматергических аксонах гиппокампа крыс и их роль в регуляции возбудимости и передатчика выпуск. J. Physiol. 576 , 235–256 (2006).
CAS
Статья
Google Scholar
Brown, J. T. & Randall, A. D. Зависимая от активности депрессия спайка после деполяризации генерирует длительную внутреннюю пластичность в пирамидных нейронах CA3 гиппокампа. J. Physiol. 587 , 1265–1281 (2009).
CAS
Статья
Google Scholar
Metz, A.E., Jarsky, T., Martina, M. & Spruston, N. Кальциевые каналы R-типа вносят вклад в постдеполяризацию и разрушение пирамидных нейронов CA1 гиппокампа. J. Neurosci. 25 , 5763–5773 (2005).
CAS
Статья
Google Scholar
Yan, H. D., Villalobos, C. & Andrade, R. Каналы TRPC опосредуют индуцированную мускариновыми рецепторами постдеполяризацию в коре головного мозга. J. Neurosci. 29 , 10038–10046 (2009).
CAS
Статья
Google Scholar
Tai, C., Hines, D. J., Choi, H. B. & MacVicar, B. A. Внедрение каналов TRPC5 в плазматическую мембрану вносит вклад в потенциал холинергического плато в пирамидных нейронах CA1 гиппокампа. Гиппокамп 21 , 958–967 (2011).
CAS
PubMed
Google Scholar
Park, J. Y. & Spruston, N. Синергетическое действие метаботропных рецепторов ацетилхолина и глутамата на возбудимость пирамидных нейронов CA1 гиппокампа. J. Neurosci. 32 , 6081–6091 (2012).
CAS
Статья
Google Scholar
Nagy, G.A. et al. Чувствительные к DAG и проницаемые для Ca 2+ каналы TRPC6 экспрессируются в зубчатых гранулярных клетках и интернейронах в образовании гиппокампа. Гиппокамп 23 , 221–232 (2013).
CAS
Статья
Google Scholar
Алле, Х. и Гейгер, Дж. Р. Комбинированное кодирование аналога и потенциала действия в мшистых волокнах гиппокампа. Наука 311 , 1290–1293 (2006).
CAS
Статья
Google Scholar
Chicurel, M. E. и Harris, K. M. Трехмерный анализ структуры и состава CA3 разветвленных дендритных шипов и их синаптических отношений с бутонами из мшистых волокон в гиппокампе крыс. Дж.Комп. Neurol. 325 , 169–182 (1992).
CAS
Статья
Google Scholar
Rollenhagen, A. et al. Структурные детерминанты передачи в больших синапсах гиппокампа, покрытых мхом. J. Neurosci. 27 , 10434–10444 (2007).
CAS
Статья
Google Scholar
Бишофбергер, Дж., Энгель, Д., Ли, Л., Гейгер, Дж. Р. и Джонас, П. Запись патч-зажима с кончиков мшистых волокон в срезах гиппокампа. Nat. Protoc. 1 , 2075–2081 (2006).
CAS
Статья
Google Scholar
Huang, Z. et al. TRIP8b-независимый трафик и пластичность корковых пресинаптических каналов HCN1 взрослых. J. Neurosci. 32 , 14835–14848 (2012).
CAS
Статья
Google Scholar
Хайнс, М. Л. и Карневале, Н. Т. Среда моделирования NEURON. Neural Comput. 9 , 1179–1209 (1997).
CAS
Статья
Google Scholar
Границы | Подпороговые мембранные токи придают особые свойства настройки, которые позволяют нейронам кодировать интеграл или производную от своих входных данных
Введение
Почти все нейроны используют потенциалы действия или пики для передачи информации.Но это не означает, что все нейроны генерируют спайки одинаково или что все спайки передают одинаковую информацию. Свойства настройки различаются между нейронами. Часто говорят, что нейроны действуют как интеграторы или детекторы совпадений в зависимости от временного интервала, в течение которого они суммируют входные данные (Abeles, 1982; König et al., 1996; Ratté et al., 2013). Когда рассредоточенные во времени входные данные кумулятивно деполяризуют нейрон до порогового значения, время интеграции велико; с другой стороны, когда совпадающие по времени входы быстро деполяризуют нейрон до порогового значения, время интеграции невелико.Согласно этому объяснению, рабочий режим зависит от временного паттерна входных сигналов (Segundo et al., 1963) и других входных свойств, таких как кинетика возбуждающего постсинаптического тока (EPSC), которые влияют на временное суммирование (König et al., 1996). Однако определенные нейроны реагируют преимущественно или даже исключительно на определенные типы входных сигналов, подразумевая, что такие нейроны имеют предпочтительный режим работы из-за присущих им свойств (Lundstrom et al., 2009; Ratté et al., 2013; Gjorgjieva et al., 2014). Настоящее исследование фокусируется на взаимосвязи между внутренними свойствами нейронов и режимом работы, и, в частности, на том, как рабочий режим вытекает из операции — интеграции или дифференцирования — выполняемой на входе ключевыми токами, зависящими от напряжения.
Напряжение-зависимые токи, действующие при подпороговых напряжениях, влияют на кинетику возбуждающего постсинаптического потенциала (ВПСП) (Fricker and Miles, 2000; Magee, 2000; Prescott and De Koninck, 2005; Gastrein et al., 2011; Remme and Rinzel, 2011), что в свою очередь влияют на временное суммирование входных данных.Но кодирование — это двухэтапный процесс, поскольку деполяризация, управляемая вводом, должна быть преобразована в спайки «все или ничего». Это цифровое преобразование также различается для разных нейронов, что очевидно из разнообразия паттернов всплесков, наблюдаемых при применении эквивалентной стимуляции к разным нейронам (Hodgkin, 1948; Llinás, 1988; Connors and Gutnick, 1990; Prescott et al., 2008a). Действительно, интеграция и обнаружение совпадений лучше всего подходят для различных стратегий преобразования: интеграторы должны повторять всплески со скоростью, пропорциональной их ступенчатой деполяризации, тогда как детекторы совпадений должны надежно вспыхивать при резкой деполяризации (что лучше всего достигается при отсутствии повторяющихся всплесков) (Gutkin et al. al., 2003; Прескотт и др., 2006; Прескотт и Сейновски, 2008). Скромное объяснение кодирования, включая различия в настройке интеграторов и детекторов совпадений, должно поэтому касаться как аналоговой обработки, так и цифрового преобразования.
Мы предположили, что те же подпороговые токи активации, ответственные за различную динамику инициирования всплесков (Prescott et al., 2008a), также модулируют траектории подпорогового напряжения, и что именно эта неразрывная комбинация перипороговых эффектов контролирует режим работы нейронов.Ранее мы сосредоточились на объяснении того, как три качественно различных паттерна пиков, каждый из которых соответствует одному из трех классов возбудимости Ходжкина (Hodgkin, 1948), возникли из континуума основных биофизических свойств (Prescott et al., 2008a). Выходя за рамки пиков, настоящее исследование стремилось определить, возникают ли кажущиеся независимыми свойства реакции, связанные с различными режимами работы, из общих биофизических механизмов. С этой целью мы построили минимальные компьютерные модели, воспроизводящие свойства реакции, наблюдаемые в двух типах спинных сенсорных нейронов, один из которых естественным образом ведет себя как интегратор, а другой как детектор совпадений.Включение медленно срабатывающего подпорогового входящего или выходящего тока было достаточным для воспроизведения полного набора характеристик отклика в интеграторах и детекторах совпадений, соответственно. Дальнейший анализ показал, как такие токи согласованно регулируют траекторию подпорогового напряжения и динамику возникновения пиков. Ключевые предсказания моделирования были экспериментально подтверждены, таким образом продемонстрировав, что медленно активирующийся подпороговый ток, в зависимости от его направления, позволяет нейрону вычислять (и кодировать) интеграл или производную от своего входного сигнала.
Методы
Нарезка и электрофизиология
Все экспериментальные протоколы были одобрены Комитетом по уходу за больными детьми и IACUC Университета Питтсбурга и были подробно описаны ранее (Prescott and De Koninck, 2002). Вкратце, взрослых самцов крыс Sprague Dawley подвергали глубокой анестезии и внутрисердечной перфузии ледяной оксигенированной (95% O 2 и 5% CO 2 ) сахарозозамещенной искусственной спинномозговой жидкостью (S-ACSF), содержащей (в мМ) 252 сахароза, 2.5 KCl, 2 CaCl 2 , 2 MgCl 2 , 10 глюкоза, 26 NaHCO 3 , 1,25 NaH 2 PO 4 и 5 кинуреновая кислота; pH 7,35; 340–350 мОсм. Спинной мозг был удален гидравлической экструзией и нарезан в парасагиттальной плоскости толщиной 300 мкм. Срезы хранили в ACSF при комнатной температуре (126 мМ NaCl вместо сахарозы и без кинуреновой кислоты; 300–310 мОсм) до записи. Срезы переносили в записывающую камеру с постоянной перфузией ~ 2 мл / мин ACSF при комнатной температуре.Нейроны просматривали с помощью градиентно-контрастной оптики с использованием микроскопа Zeiss AxioExaminer и регистрировали с помощью зажима для целых клеток с использованием усилителя Axopatch 200B (Molecular Devices, Пало-Альто, Калифорния) и электродов, заполненных (в мМ) 135 KMeSO 4 , 5 KCl , 10 HEPES и 2 MgCl 2 , 4 АТФ (Sigma, St Louis, MO), 0,4 GTP (Sigma). Трассы были отфильтрованы низкими частотами на 3-10 кГц и дискретизированы на 10 кГц.
Вычислительное моделирование
Наша модель основана на модели Морриса-Лекара (Morris and Lecar, 1981; Rinzel and Ermentrout, 1989) и подробно описана (Prescott et al., 2008а). Если не указано иное, все моделирование основано на следующих уравнениях:
CdV / dt = Istim − gleak (V − Eleak) −g¯Nam∞ (V) (V − ENa) −g¯K, drw (V − EK) −g¯subz (V − Esub) −g¯adapta ( В − ЭК) (1)
dw / dt = ϕw∞ (V) −wτw (V) (2)
dz / dt = (11 + e (βz − V) / γz − z) / τz (3)
da / dt = (11 + e (βa − V) / γa − a) / τa (4)
m∞ (V) = 0,5 [1 + tanh (V − βmγm)] (5)
w∞ (V) = 0,5 [1 + tanh (V − βwγw)] (6)
τw (V) = 1 / ch (V − βw2 · βw) (7)
, где V — напряжение, а w, z и a управляют зависимой от времени и напряжения активацией g K, dr , g sub и g Adaptor соответственно; Предполагается, что г Na активируется мгновенно, и поэтому м всегда находится в устойчивом состоянии.Следующие параметры одинаковы для интеграторов и дифференциаторов: C = 2 мкФ / см 2 ; проводимость утечки г утечка = 2 мСм / см 2 , E утечка = −70 мВ; проводимость натрия г Na = 20 мСм / см 2 , E Na = 50 мВ, β м = -1,2 мВ, γ м = 18 мВ; Калийная проводимость выпрямителя с задержкой г K, dr = 20 мСм / см 2 , E K = −100 мВ, ϕ = 0.15, β w = -10 мВ, γ w = 10 мВ; Адаптивная проводимость типа AHP г Adapt = 5 мСм / см 2 , β a = 0 мВ, γ a = 5 мВ и τ a = 20 мс. Зависимость от напряжения подпороговой проводимости g sub для интегратора и дифференциатора была эквивалентна: β z = -40 мВ и γ z = 10 мВ. Для интегратора g sub = 0,7 мСм / см 2 , τ z = 2 мс, E sub = E Na .Для дифференциатора g sub = 1,5 мСм / см 2 , τ z = 10 мс и E sub = E K . Мы не моделировали процессы, которые медленно изменяются в масштабе времени одного всплеска (например, медленная инактивация натриевых или калиевых каналов), потому что эти сверхмедленные процессы слишком медленные, чтобы динамически взаимодействовать с более быстрыми процессами, контролирующими инициирование всплеска; соответственно, эти процессы могут влиять на взаимодействия, управляющие инициированием пиков, модулируя эффективную плотность (или доступность) быстро активируемых каналов, но в остальном они не вносят вклад в соответствующие нелинейности.
Моделирование проводилось в XPP (Ermentrout, 2002) с использованием метода Эйлера с шагом по времени 0,05 или 0,1 мс. Анализ фазовой плоскости также проводился в XPP. Для расчета нулевых наклонов в момент времени t все переменные, не связанные с нулевым наклоном, сохранялись постоянными на их значениях в момент времени t . Моменты времени указаны на связанных цифрах. Бифуркационный анализ проводился с AUTO с использованием интерфейса XPP.
Анализ шумовых стимулов и спайков
Для электрофизиологических экспериментов стимуляцию применяли путем подачи тока через пипетку.Для моделирования были применены эквивалентные формы сигналов стимула через I tim . Это включало зашумленную форму волны, сгенерированную с помощью процесса Орнштейна-Уленбека (Уленбек и Орнштейн, 1930),
dζ = −ζτnoisedt + σnoiseNτξ (0,1) dt (8)
, где ξ (0,1) — случайное число, полученное из распределения Гаусса со средним 0 и единичной дисперсией, а N τ — коэффициент масштабирования 2 / τnoise, так что ζ ( t ) имеет единичную дисперсию перед масштабирование по σ шум .ξ — белый шум, а ζ — цветной шум, спектральные свойства которого контролируются временем автокорреляции τ noise . Среднее значение стимула, инициируемого спайком (STA- I стим ), рассчитывалось путем взятия среднего значения по форме волны стимула, предшествующей каждому спайку, вызванному входным цветным шумом; STA- I стим , вычисленный на основе перетасованной последовательности всплесков, был вычтен, так что окончательный STA- I стим отражает колебания стимула вокруг среднего устойчивого входного сигнала.
Ковариационный анализ проводился в соответствии с методами и условностями, описанными Fairhall et al. (2006). Цветной шум применялся к каждому модельному нейрону, как описано выше, но лежащий в основе белый шум использовался для расчета STA и ковариантности, запускаемой спайком (STC), чтобы избежать автокорреляции в стимуле. Суррогатные данные были получены с помощью того же процесса перетасовки, который описан выше.
Результаты
Экспериментальная характеристика интеграторов и дифференциаторов
Мы начали с сопоставления свойств реакции сенсорных нейронов второго порядка в пластинке I спинного дорсального рога, сосредоточив внимание на нейронах с тоническими и одиночными шипами, потому что эти нейроны олицетворяют черты (и в дальнейшем именуются) интеграторами и дифференциаторами. , соответственно.Тонические и одиночные спайковые нейроны составляют отдельные типы клеток, которые вместе составляют большинство нейронов пластинки I (Prescott and De Koninck, 2002). Скорее всего, они представляют собой тормозные и возбуждающие интернейроны соответственно (Prescott and Ratté, 2012), но это различие не влияет на настоящее исследование того, почему каждый тип клеток ведет себя так, как он ведет себя, и не было целью этого исследования изучить, как клеточные свойства влияют на сетевую функцию и обработку боли. Вместо этого мы сосредоточимся здесь на биофизической основе свойств клеток с целью определения того, как биофизически реализуются ключевые вычислительные операции.
Интеграторы многократно увеличивали пики во время длительных шагов тока, в отличие от дифференциаторов, которые увеличивали пики только в начале стимула (рис. 1A). Тем не менее, дифференциаторы могут реагировать на увеличение интенсивности стимула, не требуя промежуточной гиперполяризации, о чем свидетельствует их реакция на двухфазные шаги тока (рис. 1В). Поскольку интеграторы срабатывают повторно, они могут кодировать интенсивность стимула, модулируя частоту срабатывания, тогда как дифференцирующие не могут (рис. 1C).Интенсивность стимула также влияла на задержку всплеска в интеграторах, в то время как дифференцирующие факторы увеличивались с постоянно короткой задержкой (рис. 1D). Вариабельность латентности спайков имела важные последствия для того, как каждый тип клеток реагировал на цепочки стимулов: дифференциаторы отвечали спайками, синхронизированными по времени, на отдельные импульсы, тогда как интеграторы демонстрировали нерегулярное время спайков (рис. 1E). Эти эффекты визуализированы здесь с помощью графиков возврата, которые показывают задержку (от начала стимула) до всплеска n vs.латентность предыдущего спайка n-1 . Паттерн, наблюдаемый для интегратора, указывает на то, что на выбор времени всплеска влияют предыдущие всплески, что ставит под угрозу синхронизацию фазы с последовательностью стимулов; для сравнения, дифференциатор с его неизменно короткой задержкой пиков продемонстрировал хорошую фазовую синхронизацию. Когда клетки получали более слабые импульсы стимула, интегратор демонстрировал временное суммирование, тогда как дифференцирующий элемент — нет (рис. 1F). Отсутствие суммирования в ответе напряжения дифференциатора, несмотря на очень короткий межимпульсный интервал, примечательно, поскольку временное суммирование мембранного потенциала могло бы происходить без возникновения всплесков исключительно из-за процесса инициирования всплеска (например,g., потому что всплески должны происходить в течение короткого времени ожидания начала стимула; см. рисунок 1E).
Рис. 1. Сравнение свойств интегратора и дифференциатора в нейронах спинного мозга . Все следы пробы взяты из одной тонической или однократной клетки, которая является репрезентативной для этого типа клеток. (A) Интегратор повторяет всплески во время стимуляции, тогда как дифференцирующий элемент дает всплеск только один или два раза в начале стимула, даже в ответ на сильную стимуляцию, как показано здесь со стимуляцией 2.5 × реобаза (верхняя правая кривая). (B) Дифференциатор может реагировать на последующее увеличение интенсивности стимула. (C) Интегратор кодирует интенсивность стимула, модулируя его частоту срабатывания (здесь указывается как количество пиков на стимул длительностью 900 мс), тогда как дифференцирующее устройство неспособно к такому кодированию скорости. (D) Среднее время ожидания от начала стимула до первого всплеска (± sem, n = 5 клеток каждого типа) для интенсивностей стимула, выраженных относительно реобазы (т.е.е., минимум I стим , необходимый для вызова пиков), идентифицированный независимо для каждого нейрона. (E) Цепочки коротких импульсов умеренной амплитуды вызывают нерегулярную последовательность импульсов в интеграторе по сравнению с очень регулярной цепочкой импульсов в дифференциаторе. Точность синхронизации всплеска показана с помощью графиков возврата, на которых задержка (от начала импульса стимула) одного всплеска ( n ) отображается в зависимости от задержки предыдущего всплеска ( n -1). Открытый кружок указывает на первую точку данных.Задержка пиков в интеграторе чувствительна к прошлым пикам, в отличие от дифференциатора, где пики привязаны по времени к основному стимулу. (F) В ответ на импульсы стимула, более слабые, чем в (E) , интегратор демонстрирует временное суммирование, тогда как дифференцирующее устройство — нет, даже при очень коротких интервалах между импульсами.
Временное суммирование, наблюдаемое в нейронах с тоническими пиками, согласуется с длительным временем интеграции, в то время как его отсутствие в нейронах с одиночными пиками соответствует короткому времени интеграции.Помимо этих различий в аналоговой обработке, нейроны с тоническими и одиночными пиками, очевидно, различаются по своим свойствам цифрового преобразования не только в одноименных паттернах пиков. Паттерн пиков, задержка / точность пиков и временное суммирование представляют собой триаду размерностей отклика, по которым интеграторы и дифференциаторы находятся на противоположных крайностях, но остается без ответа, являются ли эти свойства производными от одних и тех же или разных биофизических механизмов.
Воспроизведение интегратора и дифференциатора в минимальной модели
Чтобы определить, какие биофизические механизмы определяют свойства отклика, которые различают интеграторы и дифференциаторы, мы стремились (1) воспроизвести каждый образец отклика в простейшей возможной компьютерной модели и (2) преобразовать эту модель между режимами интегратора и дифференциатора, варьируя как можно меньше параметры по возможности.Начиная с «базовой» модели, мы добавили направленный внутрь или наружу ток для воспроизведения тонического и одиночного импульса соответственно; Расчет этих подпороговых токов был основан на предыдущей работе по пиковой схеме (см. Методы). Зависимость добавленных проводимостей от напряжения была эквивалентной, таким образом выделяя реверсивный потенциал и максимальную проводимость как ключевые различия между моделями интегратора и дифференциатора. Максимальная проводимость была скорректирована для получения желаемой картины выброса (рис. 2А). Мы знали, что таким образом можно воспроизвести паттерны пиков, основываясь на нашей предыдущей работе (Prescott et al., 2008a), но здесь важно то, что все других свойств отклика были воспроизведены без дальнейших изменений параметров (Рисунки 2B – F). Это свидетельствует о том, что триада свойств отклика — паттерн пиков, задержка / точность пиков и временное суммирование — проистекает из общего биофизического механизма, а именно подпорогового тока, который был добавлен к базовой модели.
Рис. 2. Воспроизведение характеристик отклика интегратора и дифференциатора в минимальной модели .Добавление подпорогового входящего или выходящего тока к базовой модели было достаточным для воспроизведения ответов на текущие шаги (A) , ответы на двухфазные шаги (B) , кривые стимул-ответ (C) , задержка импульса (D) , точность синхронизации импульсов (E) и временное суммирование (F) , характеристика интеграторов или дифференциаторов, соответственно. Шум малой амплитуды (τ шум = 5 мс, σ шум = 1 мкА / см 2 ) был добавлен для учета влияния собственных источников шума на синхронизацию всплесков.
Как более подробно объясняется в следующем разделе, добавленный ток придает определенные свойства отклика за счет реализации отложенной обратной связи. Зависимость этого тока от напряжения определяет диапазон напряжений, в котором работает эта обратная связь; обратная связь должна работать в относительно узком диапазоне напряжений около порогового значения. Кинетика этого тока определяет, как обратная связь реагирует на входные данные с различной кинетикой; обратная связь должна действовать в первую очередь на медленных входах и, следовательно, имеет медленную постоянную времени от 2 до 10 мс, что, хотя и довольно быстро, медленнее, чем активация быстрого натриевого канала и самых быстрых компонентов входа.Медленно активирующийся подпороговый входящий или исходящий ток достаточен для наделения интеграторами или дифференциаторами, соответственно, но эти данные не исключают, что нейроны отличаются другими способами [например, нейроны с тоническим и одиночным спайком различаются по своей дендритной морфологии (Прескотт и Де Koninck, 2002) и в степени адаптации высоты шипа], но любую связь с рабочим режимом следует рассматривать как коррелятивную, пока не будет доказано обратное. Тем не менее, другие факторы могут, по крайней мере теоретически, вносить вклад в интеграторные или дифференцирующие черты (см. Ниже).Что касается этих спинномозговых нейронов, то подпороговые токи, идентифицированные здесь как передающие интеграторные и дифференцирующие признаки, как известно, существуют в нейронах с тоническими и одиночными пиками, соответственно, необходимы для связанных с ними паттернов пиков и участвуют в формировании кинетики ВПСП ( Прескотт и Де Конинк, 2005; Прескотт и др., 2008a). В частности, интеграторы выражают постоянный ток натрия и кальция, тогда как дифференциаторы выражают ток калия типа K v 1.
Различия в динамике инициирования спайков
Затем мы попытались объяснить, как медленно активирующийся подпороговый ток ( I sub ) взаимодействует с другими токами, чтобы повлиять на процесс инициирования всплеска. Нелинейная динамика, лежащая в основе инициирования спайков, была подробно описана ранее (Rinzel and Ermentrout, 1989; Borisyuk, Rinzel, 2005; Ижикевич, 2007; Prescott et al., 2008a), но здесь приводится описание для объяснения конкретных моделей, разработанных в этой статье. изучить и помочь установить экспериментально проверяемые прогнозы относительно различий между интеграторами и дифференциаторами.Более того, наши демонстрации подчеркивают биофизическую основу нелинейных взаимодействий и приводят к интерпретациям, которые отличаются от ранее опубликованных (см. Также Обсуждение).
Вкратце, бифуркационный анализ описывает, как поведение системы качественно изменяется (например, переключается с состояния покоя на повторяющиеся всплески) при изменении параметра. Такие изменения отражают нелинейное взаимодействие между системными переменными. Эти взаимодействия можно визуализировать, сопоставив одну переменную с другими переменными, чтобы создать фазовую плоскость.Нулевые линии представляют всюду вдоль фазовой плоскости, где заданная переменная не изменяется. Определение того, как пересекаются нулевые линии и как эти так называемые фиксированные точки изменяются во время бифуркации, может предсказать, как будут развиваться системные переменные. Для более подробного объяснения такого рода анализа см. Ижикевич (2007).
Начиная с интегратора, на рисунке 3A показано, как переменные V, w и z развиваются во время ступенчатого деполяризующего стимула, где w и z управляют активацией g K, dr и g. саб соответственно.Чтобы количественно оценить влияние подпорогового входящего тока, мы провели бифуркационный анализ, систематически увеличивая активацию этого тока (контролируемую z ), вместо того, чтобы позволить z свободно развиваться как переменной. Примечательно, что этот бифуркационный анализ отличается от более типичного анализа, в котором изменяется стимулирующий ток I tim . Поскольку z систематически увеличивалось, стабильная фиксированная точка дестабилизировалась посредством бифуркации Хопфа и была заменена стабильным предельным циклом; значение z , при котором это происходит, зависело от интенсивности стимула (рис. 3B).Чтобы предсказать поведение полной модели (т.е. с z , рассматриваемым как переменная), мы спроецировали зависимую от напряжения кривую активации для I sub на бифуркационную диаграмму. Перед стимуляцией (рис. 3В вверху) кривая активации пересекала бифуркационную диаграмму в области, соответствующей стабильной фиксированной точке, которая точно предсказывает, что полная модель будет находиться при подпороговом напряжении с минимальной активацией I sub . Во время стимуляции (рис. 3B внизу) бифуркационная диаграмма смещается так, что она больше не пересекает кривую активации в устойчивой фиксированной точке; в этих условиях полная модель периодически подскакивает из-за неограниченной активации I sub .Это дополнительно проиллюстрировано анализом фазовой плоскости. В фазовой плоскости Vw (рис. 3C вверху) стимуляция сдвигает нулевую линию V вверх (из красного в синее положение), но этого недостаточно, чтобы дестабилизировать пересечение между V — и w — nullclines; бифуркация Хопфа происходит только после того, как нулевая линия V сдвигается дальше (в пурпурное положение) за счет вклада I sub . Активацию I sub можно понять из фазовой плоскости Vz (рис. 3C внизу), где стимуляции достаточно, чтобы сместить нулевую линию V достаточно далеко вниз, чтобы ее устойчивое пересечение с z — nullcline разрушается через бифуркацию седло-узел на бифуркации инвариантной окружности (SNIC).Эта бифуркация не ограничивает активацию входящей внутрь субсистемы I , что приводит к деполяризации положительной обратной связи и бифуркации Хопфа в подсистеме V-w , после чего возникает повторяющийся выброс.
Рисунок 3. Динамика возникновения всплесков в интеграторе. (A) Отклик напряжения ( В ), активация I K, dr ( w ) и активация I sub ( z ) на стимулирующий ток ( I ) стим ) в зависимости от времени.Цветные точки и стрелки указывают моменты времени, проиллюстрированные в (B, C) . (B) Бифуркационные диаграммы в состоянии покоя (вверху) и во время стимуляции (внизу) с z , рассматриваемым как параметр и систематически изменяющимся от 0 до 1. Зеленые кривые показывают фиксированную точку; оранжевые кривые показывают максимум и минимум предельного цикла. Бифуркационные диаграммы показывают, что бифуркация Хопфа происходит (фиксированная точка становится нестабильной и появляется устойчивый предельный цикл), если z увеличивается достаточно высоко.Чтобы предсказать, насколько сильно возрастет z в полной модели, кривая активации, зависящая от напряжения, для I sub (серая) была наложена на бифуркационную диаграмму; отклик полной модели показан черным цветом. Без стимуляции серая кривая пересекает бифуркационную диаграмму в устойчивой фиксированной точке (красная точка), где z остается в полной модели. Во время стимуляции фиксированная точка разрушается, и система перемещается от синей точки к фиолетовой, после чего возникает всплеск. (C) Фазовые плоскости, показывающие взаимодействие V-w (вверху) и взаимодействие V-z (внизу). Нулевые линии показывают, где переменная остается постоянной; w — и z-нулевые линии показаны серым цветом; V -нулевые линии меняются со временем и показаны разными цветами, соответствующими моментам времени, указанным в (A) . В фазовой плоскости V — w стимуляция сдвигает нулевую линию V вверх (из красного положения в синее), но пересечение между линиями нуля V и w не дестабилизируется до . V -nullcline смещается дальше (в пурпурное положение) вторично при активации I sub .Активацию I sub можно понять из фазовой плоскости V — z , где стимуляция сдвигает нулевую линию V настолько вниз, что она больше не пересекает нулевую линию z . Таким образом, инициирование всплеска в интеграторе можно описать как последовательность двух бифуркаций: бифуркации SNIC в плоскости V-z , которая способствует последующей бифуркации Хопфа в плоскости V-w . Желтые прямоугольники показывают увеличенный вид критических областей. (D) Влияние постоянной времени включения подпорогового тока (τ z ) и адаптации, зависящей от пиков. (a) Несмотря на медленный запуск, пиковые значения очень быстрые для τ z = 2 мс, потому что g sub остается активным между всплесками. Активация g sub прерывается между пиками, если τ z удлиняется (b) и / или включена адаптация, зависящая от пиков (c, d) . Адаптация также служит для увеличения динамического диапазона.
Ток адаптации был включен в модель интегратора, чтобы помочь воспроизвести экспериментальные данные. На рисунке 3D показано влияние тока адаптации и изменения постоянной времени активации для подпорогового тока τ z . Если τ z длинное, первый всплеск возникает медленно, но скорость срабатывания после этого высока, потому что положительная обратная связь сначала медленно активируется, но затем остается активированной между всплесками (рис. 3Da). Это несовместимо с экспериментальными данными (рис. 1A) и исправляется путем сокращения τ z так, чтобы положительная обратная связь (т.е.е. подпороговый входящий ток) деактивируется между всплесками (рис. 3Db). Однако положительная обратная связь приводит к тому, что даже слабые надпороговые стимулы вызывают высокую частоту возбуждения, что приводит к сужению динамического диапазона. Динамический диапазон расширяется за счет включения зависимой от спайка адаптации, которая резко активируется с каждым всплеском и медленно затухает, таким образом периодически прерывая процесс положительной обратной связи (рисунки 3Dc, d). Выраженная постгиперполяризация (AHP), наблюдаемая в нейронах с тоническими импульсами, согласуется с зависимой от спайков адаптацией в этом типе клеток (Prescott and De Koninck, 2002).Эквивалентная адаптация была включена в модель дифференциатора для сравнения, но она не дала заметных эффектов.
Динамика инициирования всплеска в дифференциаторе полностью отличается от таковой в интеграторе, поскольку всплеск дифференциатора должен происходить несмотря на подпороговый внешний ток, а не с помощью подпорогового внутреннего тока. На рисунке 4A показано, как переменные V, w и z развиваются во время ступенчатого деполяризующего стимула. Бифуркационный анализ с использованием z в качестве параметра бифуркации показывает, что нет предельного цикла в отсутствие стимуляции (рис. 4B вверху).При достаточно сильной стимуляции стабильный предельный цикл существует, но только при низких значениях z (рис. 4B внизу), что подразумевает, что повторяющиеся выбросы возможны, но будут предотвращены, если I sub активируется достаточно сильно. Примечательно, что кривая активации для I sub пересекает нижнюю бифуркационную диаграмму в области, соответствующей стабильной фиксированной точке. Моделирование в полной модели показывает, что модель стабилизируется на этом перекрестке, но не раньше, чем образуется один пик.Это говорит о том, что если дифференцирующий элемент должен генерировать всплеск, он должен сделать это до того, как I sub станет слишком сильно активированным, что согласуется с результатами анализа фазовой плоскости. В плоскости Vw (рис. 4C вверху) стимуляция сдвигает нулевую линию V вверх (из красного в синее положение), что достаточно для возникновения бифуркации Хопфа, но активация I sub сдвигает . V — нулевая линия вниз, так что она снова пересекает нулевую линию w в устойчивой фиксированной точке.Активацию I sub можно понять по нулевой линии V-z (рисунок 4C внизу). Стимуляция сдвигает нулевую линию V вверх, но ее пересечение с нулевой линией z остается стабильным; Несмотря на это, симуляции в полной модели показывают, что траектория следует косвенным путем от первоначально установленной устойчивой фиксированной точки к вновь установленной устойчивой фиксированной точке. Эта обходная траектория может возникнуть из-за того, что z не изменяется мгновенно, что позволяет системе временно покинуть устойчивую фиксированную точку, когда эта точка перемещается слишком быстро (например,г., при резком начале шага раздражителя). Ускользнет ли система на достаточно большое расстояние от фиксированной точки, чтобы вызвать всплеск, объясняется тем, пересекает ли траектория квазисепаратрису — границу, разделяющую поток на фазовой плоскости. Соответственно, мы называем инициирование спайков посредством этого механизма квазисепаратрисным пересечением (Prescott et al., 2008a).
Рисунок 4. Динамика возникновения спайков в дифференциаторе. (A) переменных в зависимости от времени, как на рисунке 3A.Цветные точки и стрелки указывают моменты времени, проиллюстрированные в (B, C) . (B) Бифуркационные диаграммы в состоянии покоя (вверху) и во время стимуляции (внизу). Для существования предельного цикла требуется стимуляция, но этот предельный цикл теряется, если z увеличивается достаточно высоко. Чтобы предсказать, насколько сильно увеличится z в полной модели, кривая активации, зависящая от напряжения, для I sub (серая) была наложена на бифуркационную диаграмму; отклик полной модели показан черным цветом.Без стимуляции серая кривая пересекает бифуркационную диаграмму в устойчивой фиксированной точке (красная точка). Во время стимуляции серая кривая пересекает бифуркацию в устойчивой фиксированной точке, но система должна перемещаться от синей точки к фиолетовой, и на этом пути может образоваться всплеск. (C) Фазовые плоскости, показывающие взаимодействие V-w (вверху) и взаимодействие V-z (внизу). В фазовой плоскости V — w стимуляция сдвигает нулевую линию V вверх (из красного положения в синее), дестабилизируя пересечение между нулевыми линиями V и w в процессе, но это пересечение повторно стабилизируется, когда активация переходника I смещает нуль-линию V вниз.Активацию I sub можно понять из фазовой плоскости V — z , где стимуляция сдвигает нулевую линию V вверх, хотя ее пересечение с нулевой линией z остается стабильным. Одиночный всплеск генерируется из-за того, что квазисепаратриса смещается над синей точкой, что означает, что траектория должна следовать непрямым путем к фиолетовой точке. Другими словами, пересечение квази-сепаратрисы в подсистеме V-z соответствует фиксированной точке в подсистеме V-w , которая остается нестабильной достаточно долго для образования пика. (D) Эффекты сверхмедленной инактивации натриевых каналов моделируются как статическое изменение плотности натриевых каналов ( г Na ). Синяя и серая области показывают комбинации параметров, дающие повторяющиеся или только начинающиеся пики соответственно. График показывает, как плотность подпорогового выходящего тока ( г sub ), необходимого для начального выброса, зависит от г Na . Данные приведены для I стим = 60 мкА / см 2 ; области сдвигаются в зависимости от I стим (не показано).Всплеск только в начале может быть произведен при отсутствии подпорогового выходящего тока, если г Na достаточно низкое, но только для узкого диапазона параметров (т. Е. Серая область становится уже, поскольку г sub уменьшается), тогда как начало — только шип производится в широком диапазоне: г Na , когда наружу г sub выше. (E) Эффекты медленной инактивации натриевых каналов моделируются динамически с помощью переменной z , контролирующей инактивацию (см. Результаты).Образец ответа на двухфазные шаги стимула воспроизводит экспериментальные данные на рисунке 1B. Поскольку изменения в z описываются тем же дифференциальным уравнением, что и в исходной модели (см. Уравнение 3), его взаимодействия с другими переменными такие же, как описано на панели C для исходной модели дифференциатора.
Согласно приведенному выше описанию, инициирование всплеска в дифференциаторе зависит от конкуренции между медленно активирующимся калиевым током и быстро активирующимся натриевым током.Таким образом, примечательно, что инактивация натриевых каналов более заметна среди дифференциаторов (см. Рисунок 1), так как это эффективно снижает г Na и смещает конкуренцию в пользу пиков, возникающих только в начале (Prescott et al., 2008a; Rho и Прескотт, 2012). Если такая инактивация должна происходить медленно — настолько медленно, что инактивация фактически постоянна во время любого одного всплеска — то ее можно смоделировать как статическое изменение г Na , тогда как если инактивация происходит быстрее, ее следует моделировать как динамическая переменная.Чтобы проверить эффекты медленной инактивации, мы совместно варьировали г Na и г sub в нашей модели дифференциатора и подтвердили, что всплески только в начале могут быть произведены без подпорогового внешнего тока, но только для узкого диапазона быстрая натриевая проводимость; пики только в начале были более устойчивыми, когда модель содержала сильный подпороговый выходной ток (рис. 4D). Чтобы проверить эффекты более быстрой инактивации, мы изменили уравнение 1 (см. Методы), чтобы получить новое уравнение текущего баланса,
.
CdVdt = Istim − gleak (V − Eleak) −g¯Na m∞ (V) (1 − z) (V − ENa) −g¯K, dr w (V − EK) −g¯adapt (V − EK)
, где z теперь контролирует инактивацию быстрого натриевого тока.Все остальные параметры не изменились по сравнению с исходной моделью дифференциатора — z имеет ту же кинетику и зависимость от напряжения, что и в исходной модели дифференциатора, — но g sub отсутствует. Эта модель «инактивации» точно воспроизводила паттерн «только начало» пиков, включая реакцию на двухфазные шаги стимула (рис. 4E). Происходит такая же конкуренция между положительной и отрицательной обратной связью, что и в нашей исходной модели дифференциатора, но теперь она реализуется посредством быстрой активации и более медленной инактивации, соответственно, быстрого натриевого тока.Этот механизм может помочь придать дифференцирующие черты, но тот факт, что дифференциаторы экспрессируют каналы K v 1 и повторяются после блокады этих каналов (Prescott et al., 2008a), свидетельствует о том, что активация подпорогового выходящего тока является доминирующим (и необходимым) отрицательным механизм обратной связи в дифференциаторах в пластинке I спинного рога спинного мозга.
Подводя итог, интегратор повторяет всплески, когда бифуркация SNIC позволяет активировать положительную обратную связь подпорогового внутреннего тока, тогда как дифференциатор всплескивает только в том случае, если его квазисепаратриса пересекается, что соответствует вызванной стимулом деполяризации, опережающей отрицательную обратную связь, опосредованную (преимущественно) подпороговой. внешний ток.В обеих моделях также должна происходить бифуркация Хопфа, чтобы инициировать активацию положительной обратной связи быстро активирующегося натриевого тока. Согласно этому объяснению, процесс инициирования всплеска включает два пороговых механизма. В интеграторе оба порога (бифуркация Хопфа и бифуркация SNIC) зависят от интенсивности стимула. В дифференциаторе один порог (бифуркация Хопфа) зависит от интенсивности стимула, а другой (пересечение квазисепаратрисы) зависит от скорости изменения интенсивности стимула.Эти различия, которые имеют очевидное значение для настройки нейронов (см. Ниже), в конечном итоге сводятся к тому, взаимодействует ли медленный процесс обратной связи с быстрой положительной обратной связью, непосредственно ответственной за выбросы, или конкурирует с ней.
Дифференциальные ответы на стимулы ускорения
Пороговые механизмы, описанные выше, предсказывают различия в настройке, которые имеют решающее значение для различия между интеграцией и дифференцированием. Если всплеск дифференциатора происходит только тогда, когда вызванная стимулом деполяризация опережает отрицательную обратную связь, реализуемую внешним током, то скорость изменения интенсивности стимула будет решающим фактором в их решении о всплеске.Для интеграторов этого не предсказывают; фактически, положительная обратная связь, реализуемая подпороговым входящим током, должна смягчать внутреннюю проницаемость клеточной мембраны и тем самым поощрять (а не активно препятствовать) временное суммирование.
Дифференциальная чувствительность к скорости изменения интенсивности стимула была проверена путем применения стимулов с изменяющимся углом наклона. Как и ожидалось, модель интегратора реагировала на чрезвычайно мелкие наклоны повторяющимися пиками, тогда как модели дифференциатора реагировали только на линейные изменения, которые превышали пороговый наклон (рис. 5A).Примечательно, что подсистема V-z модели дифференциатора может испытывать бифуркацию Хопфа при высоких интенсивностях стимула; соответственно, пиковая интенсивность каждого линейного изменения была ограничена значением ниже этого порогового значения интенсивности. Мы предположили, что замедление отрицательной обратной связи (путем увеличения τ z , постоянной времени, контролирующей активацию подпорогового тока) уменьшит наклон порогового значения, что действительно произошло (рис. 5A). Более того, быстрые нарастания вызвали> 1 всплеска, как и ожидалось, пока скорость деполяризации, вызванной стимулом, продолжает превышать скорость, с которой активируется отрицательная обратная связь.
Рис. 5. Ответы на стимулы рампы. (A) Модель интегратора реагировала на чрезвычайно мелкие наклоны с помощью повторяющихся пиков, тогда как модели с дифференциатором реагировали только на крутые подъемы. Все линейные изменения, применяемые к моделям дифференциатора, были ограничены уровнем ниже интенсивности, вызывающей бифуркацию Хопфа, таким образом гарантируя, что всплески происходят через квазисепаратрисное пересечение. Модель дифференциатора была протестирована с постоянной времени активации для его подпорогового тока τ z = 10 мс (слева) и 20 мс (справа).Как и ожидалось, снижение скорости активации отрицательной обратной связи уменьшало наклон порогового стимула. Модель дифференциатора может всплывать более одного раза в ответ на надпороговые наклоны. (B) Образцы графиков от каждого типа ячеек, подтверждающие прогнозы модели. Показаны образцы ответов от двух нейронов-дифференциаторов. Эксперименты повторяли на трех клетках каждого типа.
Этот управляемый нарастанием повторяющийся всплеск, а также основной прогноз — что линейные изменения должны превышать пороговый наклон, чтобы вызвать всплеск дифференциатора — были подтверждены в 3 из 3 протестированных дифференцирующих нейронов, тогда как произвольно неглубокие линейные изменения вызвали повторяющиеся всплески в 3 из 3 протестированных нейронов интегратора ( Рисунок 5B).Примечательно, что дифференцирующие нейроны никогда не вызывали> 2 спайков в ответ на шаги деполяризации с амплитудой 2–3 × реобазы, и поэтому наблюдение 4-5 спайков во время линейного нарастания явно не связано с интенсивностью стимула. Исходя из затухания амплитуды всплеска, отказ от продолжения всплеска во время линейного изменения, вероятно, связан с инактивацией натриевого канала.
Дифференциальная обработка синаптических входов
Чувствительность к интенсивности стимула или к скорости изменения интенсивности стимула влияет на то, как обрабатывается синаптический ввод.Чтобы исследовать это, мы начали с измерения времени интеграции ( T int ) на основе возбуждающих постсинаптических токов (EPSC), применяемых через равные промежутки времени. Используя EPSC с фиксированной амплитудой, мы скорректировали интервал, чтобы определить максимальную задержку до всплеска, которая была в 8 раз меньше в модели дифференциатора по сравнению с моделью интегратора, в то время как базовая модель была промежуточной (рисунок 6A). Чтобы проверить реакцию на нерегулярный входной сигнал, содержащий широкий набор частот, мы сгенерировали цветной шум, используя процесс Орнштейна-Уленбека, и измерили T int путем вычисления среднего стимула, вызванного всплесками (STA- I стим ). .Медленно флуктуирующий вход (τ шум = 100 мс) вызвал сильные всплески в интеграционной и базовой моделях, но не в модели дифференциатора (рис. 6В), что согласуется с потребностью последней в быстрой деполяризации, которая не достигается с помощью такого рода Вход. В интеграторе и базовой модели T int составляло ~ 200 мс, что согласуется с увеличенным временем автокорреляции входных данных (рис. 6B). Все модели реагировали на более быстрый флуктуирующий вход (τ шум = 5 мс), но в то время как модель интегратора демонстрировала однофазный STA- I стим с T int = 16 мс, модель дифференциатора демонстрировала двухфазный STA- I tim , положительная фаза которого соответствует T int = 6 мс, а базовая модель была промежуточной (рис. 6C).Точное время интегрирования зависит от параметров стимула, таких как средняя интенсивность стимула, но дифференцирующий элемент всегда имел самое короткое значение, а интегратор — самое длинное. Для данных, представленных здесь, средняя интенсивность стимула была скорректирована для обеспечения сопоставимой скорости возбуждения в каждом типе клеток (см. Легенду на Рисунке 6).
Рис. 6. Ответы на синаптические стимулы. (A) Отклик модельных нейронов на регулярно разнесенные синаптические входы, смоделированные как I syn = g syn (1-e −t / τ рост ) e −t / τ распад (VE rev ), где g syn = 0.7 мСм / см 2 , нарастание τ = 0,5 мс, затухание τ = 5 мс и E об. = 0 мВ. Интервал между событиями варьировался, чтобы найти минимальную скорость, способную вызвать всплеск, и время интеграции ( T int ) оценивалось от задержки до всплеска. Чтобы смоделировать нерегулярный синаптический вход (B – E) , медленные и быстрые флуктуирующие входы были сгенерированы с использованием процесса Орнштейна-Уленбека с шумом τ = 100 мс или 5 мс. T int также измерялась как продолжительность положительной фазы стимула, вызванного спайк-триггером среднего (STA). (B) Интегратор и базовая модель ответили на медленные колебания входного сигнала с T int ≈ 200 мс, а дифференцирующий — нет, что согласуется с его потребностью в быстрой деполяризации, которая не достигается с таким типом входных сигналов. При σ шум = 10 мкА / см 2 , среднее значение I Stim составляло 10 и 25 мкА / см 2 для интегратора и базовой модели соответственно, чтобы получить частоту срабатывания около 10 Гц. (C) Все три модели реагировали на быстро меняющиеся входные данные, но в то время как модель интегратора показывала однофазный STA с T int = 16,0 мс, модель дифференциатора демонстрировала двухфазный STA с T int = 5,8 РС. Базовая модель была промежуточной с T int = 9,9 мс. При σ шум = 10 мкА / см 2 , среднее значение I Stim составляло 3, 20 и 48 мкА / см 2 для интегратора, базовой модели и дифференциатора соответственно, чтобы получить срабатывание частота около 10 Гц. (D) Примеры STA показаны для типичного интегрирующего нейрона ( T int = 30,8 мс) и дифференцирующего нейрона ( T int = 18,7 мс), измеренных с быстро меняющимся входным сигналом. Среднее значение T int ± SEM, измеренное на трех интеграторах (30,4 ± 0,4 мс), было значительно больше, чем у трех дифференциаторов (19,4 ± 1,3 мс) (* p <0,005, t -тест). Среднее значение I tim было скорректировано для каждой ячейки, чтобы обеспечить срабатывание от 5 до 10 Гц. (E) Растры показывают время всплеска в ответ на «замороженный шум», повторяющийся пять раз в одной ячейке каждого типа. Интегратор и дифференциатор часто реагировали на одни и те же входные события, хотя есть явные примеры (выделенные серым цветом), в которых реагировал только один или другой. Стрелками выделены примеры (наблюдаемые только в интеграторах), в которых реакция на предыдущее событие ввода предотвратила реакцию на последующее событие ввода, которое в противном случае вызвало бы всплеск.
На основе отличительных форм STA- I tim , выявленных при тестировании с быстро меняющимся входным сигналом, мы проверили тот же вход экспериментально.Как и в симуляциях, нейроны-интеграторы и дифференциаторы демонстрировали монофазные и двухфазные STA- I стим с, соответственно (рис. 6D). Эти данные подтверждают, что дифференциаторы предпочитают более быстрый ввод деполяризации, чем интеграторы. На основании тестирования трех нейронов каждого типа, среднее значение T int ± SEM было значительно больше в нейронах-интеграторах (30,4 ± 0,4 мс), чем в нейронах-дифференциаторах (19,4 ± 1,3 мс) ( p <0,005, т -тест).Повторение одного и того же быстро меняющегося стимула несколько раз в одном и том же нейроне позволило выявить некоторые дополнительные наблюдения (рис. 6Е). Во-первых, нейроны-интеграторы и дифференцирующие нейроны имели тенденцию надежно отвечать в ходе испытаний, но два типа нейронов не обязательно реагировали на одни и те же входные колебания. Кроме того, нейрон интегратора демонстрирует паттерн, в котором реакция на более раннее колебание может помешать ему реагировать на последующие колебания, что напоминает данные на рисунках 1E, 2E.
Мы вернулись к модели, чтобы сравнить STA- I стим с активацией I sub , который мы изображаем здесь как средний подпороговый ток, инициированный всплесками (STA- I sub ). Мы пришли к выводу, что положительная фаза дифференциатора STA- I tim является короткой, потому что это минимизирует активацию внешнего I sub , тогда как положительная фаза интегратора STA- I tim относительно длинна, потому что это использует активацию входящего I sub .Сравнение STA- I sub для интеграторов и дифференциаторов подтвердило эту предсказанную закономерность (рис. 7A). Поскольку изменение напряжения зависит от суммы I tim и I sub (вместе с другими трансмембранными токами), мы построили график STA- I tim против STA- I sub . (Рисунок 7B). Точки нанесены с интервалом в 1 мс, чтобы помочь визуализировать скорость изменения. Траектория интегратора изгибается против часовой стрелки, подтверждая, что медленная деполяризация, управляемая I стимулом , вызывает активацию внутреннего I sub , так что к моменту возникновения всплеска (в конце траектории) I стим и I sub вносят равный вклад в деполяризацию.С другой стороны, траектория дифференциатора вращается по спирали в направлении по часовой стрелке, показывая, как медленная начальная гиперполяризация I стим вызывает чистый внутренний ток за счет деактивации внешнего I sub ; за этим следует быстрая деполяризация, так что внешняя I sub превышает его базовую активацию всего за 3 мс до пика. Если бы деполяризация была медленнее, I sub активировался бы сильнее, что привело бы к более плотной спирали, меньшей деполяризации и отсутствию всплеска.Пороговое значение напряжения соответствует вертикальной границе на Рисунке 7B. Следует понимать, что траектория дифференциатора может не пересекать эту границу (даже если I Stim очень сильна), потому что активация направленного наружу I sub заставляет траекторию отклоняться. Напротив, активация внутреннего переходника I заставляет траекторию отклоняться к такой границе, таким образом увеличивая вероятность достижения порогового значения напряжения.Эти результаты подчеркивают, как избирательность для различных характеристик стимула (то есть способность различных форм волны стимула вызывать спайк) зависит от того, как стимул взаимодействует с субпороговым током.
Рисунок 7. Избирательность функций зависит от того, как I SIM взаимодействует с I sub . (A) Средние значения, вызванные выбросами, были рассчитаны для стимула (STA- I стим ) и для активированного подпорогового тока (STA- I sub ).Деполяризационный ток показан как отклонение вверх (положительное). Среднее значение I tim составило 5 мкА / см 2 и 80 мкА / см 2 для интегратора и дифференциатора соответственно; σ шум составил 10 мкА / см 2 для обоих. (B) Те же данные, что и в A, но теперь с графиками I tim и I sub относительно друг друга, а не относительно времени. Точки нанесены с интервалом в 1 мс, чтобы проиллюстрировать скорость, с которой изменяется каждая переменная, где большой интервал означает быстрое изменение.Для интегратора I стим вызывает относительно медленную деполяризацию, тем самым давая время на активацию внутрь I sub , что приводит к изгибу траектории против часовой стрелки. Изгиб против часовой стрелки отражает кооперативность между I стимулом и внутренним I sub , что увеличивает способность медленных колебаний стимула управлять надпороговой деполяризацией. Для дифференциатора начальное медленное уменьшение I стим вызывает отключение внешнего переходника I , создавая таким образом чистый входящий ток.Последующее быстрое увеличение I стимула вызывает деполяризацию, сводя к минимуму время, доступное для активации внешнего переходника I до того, как будет инициирован всплеск. Изгиб этой траектории по часовой стрелке отражает конкуренцию между I стимулом и внешним I sub , что гарантирует, что только быстрые флуктуации стимула вызывают надпороговую деполяризацию.
Спайк-триггерный ковариационный анализ чувствительности к характеристикам стимула
Как показывают ответы на линейную стимуляцию (рис. 5), пиковое значение дифференциатора чувствительно к скорости изменения интенсивности стимула и к самой интенсивности стимула.В этом отношении ни наша модель дифференциатора, основанная на проводимости, ни наши дифференцирующие нейроны не являются чистыми дифференциаторами. Мы предположили, что эта совместная чувствительность к интенсивности стимула и скорости ее изменения возникает из двух требований для инициирования спайка дифференциатора: бифуркация Хопфа в подсистеме Vw зависит от интенсивности стимула, в то время как пересечение квази-сепаратрисы в Vz Подсистема зависит от скорости изменения интенсивности стимула (см. рисунок 4).Оба порога должны быть соблюдены. Для сравнения, инициирование всплеска интегратора зависит от бифуркации Хопфа в подсистеме V-w или бифуркации SNIC в подсистеме V-z (см. Рисунок 3). Бифуркация SNIC несущественна, если интенсивность стимула достаточна, чтобы вызвать бифуркацию Хопфа сама по себе; с другой стороны, бифуркация Хопфа становится неизбежной, как только происходит бифуркация SNIC, пока стимуляция не прекращается резко. Другими словами, два порога интенсивности составляют один порог интенсивности, точное значение которого может варьироваться, в отличие от двойного порога, предсказанного для дифференциаторов.Подводя итог, мы пришли к выводу, что чувствительность к различным характеристикам стимула, а именно к интенсивности стимула и ее производной, является прямым следствием двух динамических процессов, составляющих процесс инициирования спайков.
Для дальнейшего изучения этого мы провели анализ ковариации, запускаемой спайками, чтобы определить особенности стимула, к которым чувствителен каждый механизм инициирования спайков (см. Методы). Как объяснили Fairhall et al. (2006), этот анализ определяет направления в пространстве стимулов, по которым отклонения различаются по сравнению с предыдущим распределением стимулов; собственное значение каждого направления раскрывает особенности стимула (т.е., направления), к которым нейрон чувствителен. Ковариация, вызванная всплеском, выявила только одно значимое собственное значение для модели интегратора, тогда как два значимых собственных значения были обнаружены для модели дифференциатора (рис. 8A). Для обеих моделей собственный вектор наименьшего собственного значения (обозначенный как признак 1 ) напоминает STA для этого типа ячейки (рисунок 8B). Для дифференциатора собственный вектор для второго наименьшего собственного значения ( признак 2 ) напоминает производную признака 1, но неструктурирован в случае интегратора (рисунок 8B).Затем мы спроецировали ансамбль стимулов, запускаемых спайком, на две функции (рис. 8C). Спайковые стимулы образуют отчетливое облако, которое можно сравнить с предыдущим. STA- I tim описывает, как среднее значение этого облака смещено относительно предыдущего, в то время как сжатие облака отражает информацию, предоставляемую каждой функцией. Для интегратора STA- I tim указывает горизонтально, и только функция 1 предоставляет информацию. Для дифференциатора STA- I tim точек по диагонали, и обе функции 1 и 2 предоставляют информацию, таким образом подтверждая наш прогноз.Применяя такой же анализ к экспериментальным данным, полученным от дифференциаторов в слуховом стволе мозга, Slee et al. (2005) обнаружили аналогичные результаты, в частности, что блокада низкопорогового калиевого тока изменила проекцию на вторую особенность (и прогнозируемый оттуда пик), хотя вторая особенность все еще была очевидна.
Рис. 8. Различия в свойствах кодирования, выявленные методом ковариационного анализа . Параметры стимула были такими, как показано на рисунке 7, но основной белый шум использовался для всего анализа. (A) Собственные значения, упорядоченные по рангу, показаны для данных, запускаемых спайком (кружки), и для суррогатных данных (черная линия), основанных на ансамбле стимулов, запускаемых спайком, найденном после перетасовки времен спайков. Десять наборов суррогатных данных были сгенерированы путем перетасовки времен всплесков. Две наименьшие собственные моды значительно отклонялись от суррогатных данных ( p <0,01; однократные t -тесты) для дифференциатора, тогда как только одна собственная мода значительно отклонялась в случае интегратора. (B) STA- I tim и собственные векторы, соответствующие двум наименьшим собственным модам, которые обозначены как feature 1 и 2 . (C) Когда стимулы, вызванные спайком, проецируются на объекты 1 и 2, они образуют облако (черные точки), которое можно сравнивать с суррогатными данными, проецируемыми на те же два объекта (серые точки). Розовые стрелки, соответствующие STA- I tim , показывают разницу в центре масс каждого облака.Размеры черного облака относительно серого облака показывают особенности стимула, о которых шип передает информацию. Эти данные утверждают, что интеграторы кодируют информацию об одной характеристике стимула, а именно об интенсивности стимула, тогда как дифференциаторы кодируют информацию как об интенсивности стимула, так и о скорости его изменения.
Обсуждение
Используя комбинацию моделирования и экспериментов, мы исследовали биофизические механизмы, с помощью которых нейроны вычисляют интеграл или производную своих входных данных.Мы обнаружили, что интеграторы и дифференциаторы различаются свойствами множественного отклика — временным суммированием, задержкой / точностью пиков и паттерном пиков — но все эти различия имеют общее биофизическое объяснение, а именно направление медленно активирующегося подпорогового тока. Направление этого тока определяет, будет ли медленная обратная связь по субпороговому напряжению положительной или отрицательной. Это, в свою очередь, определяет, поощряется или не приветствуется временное суммирование, а также приводит к принципиально другим механизмам установления пороговых значений, основанным на том, как медленно активирующийся подпороговый ток взаимодействует с быстро активирующейся положительной обратной связью, ответственной за выбросы.
В интеграторах подпороговое значение , направленное внутрь тока, создает петлю положительной обратной связи, которая поддерживает деполяризацию и взаимодействует совместно с быстро активирующимся натриевым током, создавая повторяющиеся выбросы со скоростью, пропорциональной интенсивности стимула. Петля положительной обратной связи инициируется, когда интенсивность стимула превышает порог, необходимый для создания бифуркации SNIC во взаимодействии между напряжением и переменной активации для подпорогового тока. Это отличается от стандартного динамического объяснения возбудимости класса 1, которое объясняет медленные повторяющиеся всплески как происходящие из-за бифуркации SNIC во взаимодействии между напряжением и переменной восстановления w (где w представляет активацию калиевого тока выпрямителя с задержкой) ( Ижикевич, 2007; Прескотт и др., 2008a; Прескотт, 2015). В нашей модели интегратора эффекты положительной обратной связи контролируются постгиперполяризацией (AHP), вызванной спайк-зависимой адаптацией. Это составляет двухтактный механизм: подпороговый входящий ток «подталкивает» мембранный потенциал вверх (ко второму порогу, определяемому бифуркацией Хопфа во взаимодействии между напряжением и переменной восстановления), в то время как AHP «подталкивает» мембранный потенциал вниз. Это приводит к сильной внутренней динамике, которая зависит от кинетики AHP.Ранее было показано, что эта внутренняя динамика линеаризует кривую ввода-вывода (Ermentrout, 1998) и вызывает формирование шума за счет введения отрицательных корреляций между спайковыми интервалами (Prescott and Sejnowski, 2008). Оба эффекта избирательно улучшают скоростное кодирование медленных сигналов с плотными выбросами (Chacron et al., 2004) и, таким образом, актуальны для интеграторов, работающих в режиме, управляемом средним или колеблющимся. В режиме, управляемом флуктуациями, который характеризуется редкими всплесками, каждый из которых вызывается отдельными флуктуациями стимула, AHP может предотвращать взрыв, но его можно пренебречь от одного всплеска, вызванного флуктуациями, к другому.В этих условиях управление траекторией подпорогового напряжения с положительной обратной связью — «толкающий» механизм — увеличивает временное суммирование. Наш анализ на основе шума был сосредоточен на области, вызванной колебаниями.
Для сравнения, подпороговый , направленный наружу ток, выражаемый дифференциаторами, создает петлю отрицательной обратной связи, которая усекает деполяризацию и предотвращает повторяющиеся выбросы за счет конкурентного взаимодействия с быстро активирующимся натриевым током. Эквивалентная отрицательная обратная связь в принципе может быть достигнута путем медленной инактивации быстро активирующегося натриевого тока.Можно сказать, что любой механизм опосредует форму аккомодации порога спайков . Изолированный спайк возникает, когда опосредованная стимулом деполяризация вызывает квазисепаратрисное пересечение, что равносильно уходу от отрицательной обратной связи путем опережения ее активации. Согласно этому объяснению, инициирование всплеска зависит от скорости изменения интенсивности стимула, превышающей некоторый порог, а крутизна порога, естественно, зависит от кинетики активации подпорогового внешнего тока.Lundstrom et al. (2009) тщательно продемонстрировали, как кинетика этой обратной связи настраивает чувствительность нейронов к колеблющимся входным сигналам. Наши модели предсказали, и эксперименты подтвердили, что дифференциаторы повторяют всплески, пока скорость изменения интенсивности стимула превышает пороговую крутизну. По крайней мере теоретически, частота таких всплесков может кодировать производную от стимула. На практике быстро деполяризующий стимул (например, вызванный залпом синхронных входных сигналов), вероятно, будет настолько коротким, что во время него может произойти только один всплеск.Время таких всплесков указывает на то, когда произошли события надпорогового стимула, а частота этих всплесков указывает на частоту событий надпорогового стимула. Наиболее важно то, что дифференциаторы из-за своей избирательности в отношении быстро деполяризованного входа избирательно реагируют на синхронные входные данные; кроме того, набор таких нейронов, получающих общий входной сигнал, может отвечать синхронными импульсами, обеспечивая тем самым хорошее синхронное кодирование (Hong et al., 2012; Ratté et al., 2013). Очевидно, что интеграторы могут реагировать на синхронные входы, но набор таких нейронов не обязательно отвечает хорошей синхронизацией выходных сигналов.Фактически, дифференциаторы всегда работают в режиме, управляемом флуктуациями, и, более того, избирательно реагируют на флуктуации, содержащие относительно высокие частоты, из-за фильтрации верхних частот, осуществляемой подпороговым выходным током.
Согласно пояснениям, приведенным выше (см. Также рисунки 3, 4), пиковое значение имеет два пороговых значения. В интеграторе один порог (бифуркация SNIC) управляет активацией подпорогового внутреннего тока, а другой (бифуркация Хопфа) управляет активацией быстро активирующегося натриевого тока.В дифференциаторе один порог (пересечение квазисепратрисы) описывает, опережает ли вызванная стимулом деполяризация активацию подпорогового внешнего тока, в то время как другой является той же бифуркацией Хопфа, наблюдаемой в интеграторе. Наличие двух пороговых значений вызывает вопрос о том, преобладает ли один порог над другим, и если нет, то как они сочетаются. В случае интеграторов достижение первого порога инициирует процесс положительной обратной связи, который при наличии достаточного времени гарантирует достижение второго порога.Соответственно, важность второго порога зависит от кинетики входа относительно кинетики активации подпорогового тока; другими словами, второй порог становится важным только в том случае, если вызванная стимулом деполяризация настолько коротка, что второй порог должен быть достигнут без помощи подпорогового входящего тока; в этих условиях стимула нейрон вообще не использует свою интегрирующую способность. В дифференциаторе действуют оба порога, поскольку деполяризация должна быть не только быстрой, но и достаточной по амплитуде для возникновения всплеска.Первый механизм пороговой обработки формирует траекторию напряжения до того, как это напряжение достигает второго порога, после чего всплеск становится действительно неизбежным.
Приведенные здесь объяснения инициирования спайков отличаются от предыдущих описаний (Rinzel and Ermentrout, 1989; Borisyuk, Rinzel, 2005; Izhikevich, 2007; Prescott et al., 2008a) и, возможно, обеспечивают более глубокое понимание биофизических основ этого процесса. Это зависит от использования нами модели, основанной на проводимости, которая все еще является низкоразмерной, но немного более сложной, чем простейшие модели, к которым применяется большая часть нелинейного динамического анализа.Тем не менее, наши модели нейронов по-прежнему чрезвычайно просты по сравнению с реальными нейронами, что заставляет подозревать, что пространственно сложные нейроны, которые выражают различные ионные каналы с неоднородной плотностью, могут иметь несколько механизмов пороговой обработки, которые каким-то образом объединяются, чтобы дать глобальный порог (Ashida et al. , 2007; Brette, 2013). Тем не менее, один и тот же нейрон может комбинировать пороги тонко по-разному в зависимости от статистики стимула, подразумевая, что порог может меняться, по крайней мере количественно, в зависимости от стимула (Famulare and Fairhall, 2010) (см. Ниже).Между нейронами существуют качественные различия, примером чему служит наше сравнение нейронов с тоническим и одиночным шипом в спинном роге спинного мозга. Более того, внутренние свойства нейронов подвержены модуляции, что означает, что порог может изменяться (Prescott et al., 2008b; Hong et al., 2012).
Вышеупомянутое обсуждение подчеркивало нейронные свойства и нейрон-центрическое определение рабочего режима. Предыдущие обсуждения рабочего режима чаще всего подчеркивали влияние свойств стимула (Mainen and Sejnowski, 1995; Nowak et al., 1997; Азуз и Грей, 2000, 2003; Рудольф и Дестекс, 2003; Аксмахер и Майлз, 2004 г.). Оба важны. Это проиллюстрировано реакциями на шумные стимулы (см. Рисунок 6): интегратор реагировал как на медленные, так и на быстро меняющиеся входные сигналы с помощью STA, различие которых полностью объясняется разницей во времени автокорреляции стимулов; однако нейроны интегратора и дифференциатора ответили на один и тот же быстро колеблющийся стимул с помощью STA, различие которых полностью объясняется различием в свойствах нейронов, а именно их подпороговом токе.В самом деле, как показано на рисунке 6E, два типа клеток не обязательно реагируют на одни и те же стимулы. Это не меняет того факта, что интеграторы чувствительны к колебаниям стимулов; они просто менее избирательны в отношении этих флуктуаций, чем дифференциаторы (Ratté et al., 2013), что согласуется с наблюдением, что дифференциаторы вообще не реагируют на медленно флуктуирующие стимулы, тогда как интеграторы реагируют. Короче говоря, различия в траектории подпорогового напряжения и инициировании всплеска приводят к различиям в настройке, которые напрямую отражаются в вызванном всплеском среднем значении и ковариации (Aguera y Arcas and Fairhall, 2003; Gutkin et al., 2005; Fairhall et al., 2006; Ermentrout et al., 2007; Hong et al., 2007; Артур и др., 2013). Настоящее исследование расширяет попытки связать свойства нейронального кодирования со свойствами инициирования спайков и, кроме того, с биофизическими свойствами.
Хотя мы подчеркивали влияние различных подпороговых токов, необходимо понимать, что пороги возникают из-за нелинейного взаимодействия между токами; следовательно, подпороговые токи следует рассматривать в контексте других токов, выражаемых ячейкой.Чтобы проиллюстрировать это, предположим, что слабый медленно активирующийся калиевый ток будет представлять небольшую конкуренцию быстро активирующемуся натриевому току, который особенно силен и, таким образом, не смог бы придать дифференцирующие свойства этому нейрону; однако, если плотность натриевых каналов была уменьшена, та же самая плотность калиевых каналов могла бы предотвратить повторяющиеся пики и реализовать отличительные черты (Lundstrom et al., 2008). Точно так же кумулятивная инактивация натриевых каналов может снизить доступность натриевых каналов (Fernandez et al., 2011), а повышенная проводимость утечки может увеличивать перипороговый внешний ток (Zsiros and Hestrin, 2005; Prescott et al., 2006; Broicher et al., 2012), причем оба эффекта имеют тот же эффект, что и уменьшение плотности натриевых каналов или увеличение медленно активирующихся каналов. плотность калиевых каналов. Действительно, моделирование на Рисунке 4 продемонстрировало, что инактивация натриевого канала и активация калиевого канала (с эквивалентной кинетикой и зависимостью от напряжения) могут реализовывать одну и ту же отрицательную обратную связь и, таким образом, давать одни и те же отличительные черты.Lundstrom et al. (2009) смоделировали отрицательную обратную связь как комбинацию двух процессов. Но, как и канонические дифференциаторы в слуховом среднем мозге (Reyes et al., 1996; Slee et al., 2005; Mathews et al., 2010; Higgs and Spain, 2011), некоторые пирамидные нейроны в неокортексе (Guan et al., 2007 ) и нейроны ганглия задних корешков (Ratté et al., 2014), дифференциаторы спинного дорсального рога полагаются на ток K v 1 для их свойств ответа (Prescott et al., 2008a). Точно так же интеграторные черты могут передаваться инактивацией калиевого тока (Connor, Stevens, 1971; Cudmore et al., 2010; Barreiro et al., 2012) или активацией натриевых или кальциевых потоков (Fricker and Miles, 2000; Gonzalez-Burgos and Barrionuevo, 2001; Vervaeke et al., 2006; Carter et al., 2012). Последние встречаются в спинномозговых нейронах, вызывающих тонические импульсы (Prescott, De Koninck, 2005; Prescott et al., 2008a).
Подводя итог, можно сказать, что процесс инициирования всплеска является критическим компонентом нейронного кодирования. Скачки не возникают мгновенно, когда напряжение пересекает заранее установленный порог; Напротив, всплески могут быть инициированы посредством качественно различных механизмов, которые зависят от нелинейного взаимодействия между вносящими вклад мембранными токами.Когда медленно активирующийся подпороговый ток направлен внутрь, он будет взаимодействовать с быстро активирующимся током. В этих условиях инициирование всплеска зависит от кумулятивной деполяризации, достигающей порога, представленного бифуркацией SNIC, после чего активация положительной обратной связи медленного входящего тока может гарантировать активацию быстрого тока. Когда медленно активирующийся подпороговый ток направлен вовне, он будет конкурировать с быстро активирующимся током. В этих условиях инициирование всплеска зависит от деполяризации, активирующей быстрый ток достаточно быстро, чтобы медленный ток не мог догнать, что представлено пересечением квази-сепаратрисы.В обоих случаях стимул должен иметь достаточную интенсивность, чтобы вызвать всплеск, но его взаимодействие с подпороговым током будет управлять предпочтительной формой волны стимула: интеграторы настроены на медленный деполяризационный вход — вид входа, который дает время подпорогового внутреннего тока для облегчения пики — тогда как дифференциаторы настроены на вход с быстрой деполяризацией — вид входа, который дает подпороговому внешнему току слишком мало времени для предотвращения пиков. Таким образом, интеграция и дифференциация возникают из-за противоположно направленных мембранных токов и реализуются как часть процесса инициирования спайков.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта работа была поддержана грантом R01 NS076706 Национального института здравоохранения и грантом на открытие Национального совета по научным и инженерным исследованиям Канады. Стивен А. Прескотт также является 53-м стипендиатом Маллинкродта и новым исследователем Канадского института медицинских исследований.
Список литературы
Борисюк А., Ринзель Дж. (2005). «Понимание динамики нейронов путем геометрического разделения минимальных моделей», в Методы и модели в нейрофизике, Летняя школа Proc Les Houches , редакторы К. Чоу, Б. Гуткин, Д. Хансель, К. Менье и Дж. Далибард (New Йорк, Нью-Йорк: Эльзевир), 19–72.
Google Scholar
Бройхер Т., Малерба П., Дорваль А. Д., Борисюк А., Фернандес Ф. Р. и Уайт Дж. А. (2012). Фазовая синхронизация спайков в пирамидных нейронах CA1 зависит от фоновой проводимости и скорости возбуждения. Дж. Neurosci . 32, 14374–14388. DOI: 10.1523 / JNEUROSCI.0842-12.2012
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кадмор, Р. Х., Фронзароли-Молиньер, Л., Жиро, П., и Дебанн, Д. (2010). Точность импульсов и синхронизация сети контролируются гомеостатическим регулированием калиевого тока D-типа. J Neurosci 30, 12885–12895. DOI: 10.1523 / JNEUROSCI.0740-10.2010
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Эрментроут, Б.(2002). Моделирование, анализ и анимация динамических систем: руководство по XPPAUT для исследователей и студентов . Филадельфия, Пенсильвания: СИАМ.
Google Scholar
Фэрхолл, А. Л., Бурлингейм, К. А., Нарасимхан, Р., Харрис, Р. А., Пучалла, Дж. Л., и Берри, М. Дж. (2006). Селективность множественных стимулов в ганглиозных клетках сетчатки. J. Neurophysiol . 96, 2724–2738. DOI: 10.1152 / jn.00995.2005
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Фернандес, Ф.Р., Бройхер Т., Чыонг А. и Уайт Дж. А. (2011). Колебания напряжения на мембране снижают адаптацию частоты всплесков и сохраняют усиление выходного сигнала в пирамидных нейронах CA1 в состоянии высокой проводимости. Дж. Neurosci . 31, 3880–3893. DOI: 10.1523 / JNEUROSCI.5076-10.2011
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Гастрейн П., Кампанак Э., Гасселин К., Кадмор Р. Х., Бяловас А., Карлье Э. и др. (2011). Роль катионного тока, активированного гиперполяризацией, в точности времени пика и внутреннем резонансе в корковых нейронах in vitro . J. Physiol . 589, 3753–3773. DOI: 10.1113 / jphysiol.2011.209148
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Гонсалес-Бургос, Г., и Баррионуево, Г. (2001). Напряжение-управляемые натриевые каналы формируют подпороговые epsps в пирамидных нейронах слоя 5 префронтальной коры головного мозга крысы. J. Neurophysiol . 86, 1671–1684.
Pubmed Реферат | Pubmed Полный текст | Google Scholar
Гуань, Д., Ли, Дж. К., Хиггс, М.Х., Испания, У. Дж. И Феринг, Р. К. (2007). Функциональная роль каналов Kv1 в пирамидных нейронах неокортекса. J. Neurophysiol . 97, 1931–1940. DOI: 10.1152 / jn.00933.2006
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ижикевич, Э. М. (2007). Динамические системы в неврологии . Кембридж, Массачусетс: MIT Press.
Google Scholar
Лундстрем, Б. Н., Фамуларе, М., Соренсен, Л. Б., Испания, В.Дж. И Фэрхолл А. Л. (2009). Чувствительность скорости возбуждения к входным колебаниям зависит от разделения шкалы времени между быстрыми и медленными переменными в отдельных нейронах. J. Comput. Neurosci . 27, 277–290. DOI: 10.1007 / s10827-009-0142-x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Лундстрем, Б. Н., Хонг, С., Хиггс, М. Х., Фэрхолл, А. Л. (2008). Два вычислительных режима однокамерного нейрона, разделенного плоской границей в пространстве проводимости. Нейронные вычисления . 20, 1239–1260. DOI: 10.1162 / neco.2007.05-07-536
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Мэтьюз П. Дж., Джерког П. Э., Ринзел Дж., Скотт Л. Л. и Голдинг Н. Л. (2010). Управление субмиллисекундной синаптической синхронизацией в детекторах бинауральных совпадений по каналам K (v) 1. Nat. Neurosci . 13, 601–609. DOI: 10.1038 / № 2530
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Прескотт, С.А. (2015). «Возбудимость: классы I, II и III Ходжкина», в Encyclopedia of Computational Neuroscience , ред. Д. Джагер и Р. Юнг (Нью-Йорк, Нью-Йорк: Спрингер), 1–7.
Прескотт, С. А., Де Конинк, Ю. (2005). Время интеграции в подмножестве нейронов пластинки I спинного мозга удлиняется за счет синергетического действия натриевых и кальциевых потоков, продлевая подпороговую деполяризацию. Дж. Neurosci . 25, 4743–4754. DOI: 10.1523 / JNEUROSCI.0356-05.2005
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Прескотт, С.А., Ратте, С., Де Конинк, Ю., и Сейновски, Т. Дж. (2006). Нелинейное взаимодействие между шунтированием и адаптацией управляет переключением между интеграцией и обнаружением совпадений в пирамидных нейронах. Дж. Neurosci . 26, 9084–9097. DOI: 10.1523 / JNEUROSCI.1388-06.2006
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Рейес, А. Д., Рубель, Э. У., и Испания, В. Дж. (1996). In vitro анализ оптимальных стимулов для фазовой синхронизации и отложенной по времени модуляции возбуждения в нейронах ламинарного ядра птичьего ядра. Дж. Neurosci . 16, 993–1007.
Pubmed Реферат | Pubmed Полный текст | Google Scholar
Ро, Ю.А., Прескотт, С.А. (2012). Выявление молекулярных патологий, достаточных для возникновения нейропатической возбудимости в первичных соматосенсорных афферентах, с использованием теории динамических систем. PLoS Comput. Биол . 8: e1002524. DOI: 10.1371 / journal.pcbi.1002524
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ринзель, Дж.и Б. Эрментрут (1989). «Анализ нервной возбудимости и колебаний», в Методы нейронного моделирования , ред. К. Кох и И. Сегев (Кембридж: MIT Press), 251–291.
Второй, Дж. П., Мур, Г. П., Стенсас, Л. Дж., И Буллок, Т. Х. (1963). Чувствительность нейронов аплизии к временному паттерну приходящих импульсов. J. Exp. Биол . 40, 643–667.
Pubmed Реферат | Pubmed Полный текст | Google Scholar
Уленбек, Г. Э., и Орнштейн, Л.С. (1930). К теории броуновского движения. Phys. Ред. . 36, 823–841. DOI: 10.1103 / PhysRev.36.823
CrossRef Полный текст | Google Scholar
Использование подпороговых событий для характеристики функциональной архитектуры электрически связанной сети нижних олив
Существенных изменений:
Эксперименты и моделирование выполнены хорошо, анализ интересен, а выводы исследования потенциально очень важны.Есть несколько опасений по поводу основных результатов — происхождения IRE и анализа сетей (нейронных кластеров), которые необходимо рассмотреть при пересмотре. Подробные комментарии рецензентов дополняют эти основные необходимые изменения, описанные ниже; при пересмотре акцент должен быть сделан на этих основных моментах, а не на незначительных различиях в комментариях рецензента.
Спасибо за обнадеживающий ответ. Рецензенты проделали тщательную работу, которая завершилась важными комментариями, которые, несомненно, приведут к значительному улучшению презентации и заключений этого исследования.В отредактированной рукописи мы реорганизовали описание данных, существенно расширили описание статистических методов, используемых для фармакологического лечения и подхода к моделированию, и постарались ответить на все комментарии рецензентов. Однако это исследование представляет собой попытку исследовать структуру электрически связанных сетей путем анализа подпороговых событий, которые представляют электрическую связь. Таким образом, основной проблемой является идентификация этих событий. В наших записях нейронов нижних олив мы встретили электрически связанные колоски, а также другие подпороговые события, IRE, которые необходимо было идентифицировать, прежде чем исключить их из анализа.Мы предполагаем, что они представляют собой регенеративные ответы в шипах, которые расположены в начальном сегменте аксона. Мы согласны с тем, что это «важное новое открытие», которое требует тщательного изучения. Фактически, мы начали новое направление исследований, в котором анатомические, физиологические и методы визуализации будут использоваться для проверки нашего предположения о происхождении этих событий. Таким образом, в настоящее время мы можем предоставить только анекдотические наблюдения за шипами на аксоне у мыши. Тщательное изучение распределения спайков во многих нейронах, а также корреляция между количеством шипов и количеством IRE в данном нейроне и специфической активацией этих шипов абсолютно необходимы и не могут быть предоставлены на данном этапе.Тем не менее, мы постарались ответить на комментарии рецензентов и надеемся, что они получат ваше одобрение.
1) Все обозреватели согласны с тем, что синаптическая фармакология должна быть лучше как для IRE, так и для рассказов о колосках. Авторы заявляют, что они протестировали 19 ячеек в CNQX / APV, поэтому, возможно, все, что осталось, — это провести более подробный анализ и задокументировать его в документе.
В отредактированной рукописи мы предоставили лучшее и более полное описание фармакологических экспериментов.Сначала мы сравнили частоту возникновения спонтанных событий, демонстрируя, что, хотя частота колосков резко снижена (n = 9 клеток), IRE полностью отсутствуют (n = 4 клетки). Как было предложено, мы добавили подробное описание влияния синаптических блокаторов на форму колосков и появление вызванных колосков (Рисунок 2 — дополнение к рисунку 1; Рисунок 6 — приложение к рисунку 1), чтобы показать, что снижение частоты происходит из-за снижение возбудимости сети, и не потому, что некоторые из регистрируемых нами колосков имеют синаптическое происхождение.
IRE, которые являются внутренними событиями, не встречаются в присутствии синаптических блокаторов, но также не имеют синаптического происхождения, как показано на Рисунке 2 (Рисунок 3 в оригинальной рукописи). Поскольку большая часть потенциалов действия в нейронах IO запускается IRE, в присутствии синаптических блокаторов частота потенциалов действия сильно снижается, и, следовательно, частота колосков. Однако колоски все еще могут быть вызваны синаптическими блокаторами, когда потенциалы действия вызываются в связанной клетке (Рисунок 6 — рисунок в приложении 1).
Мы изменили порядок появления рисунков 2 и 3 исходной рукописи, чтобы объяснить и обсудить эти результаты.
2) Действительно ли аксональные шипы так же обычны и красивы в клетках мыши, как показал Де Зеув у кошек? Авторы уже имели заполненные красителем IO-клетки мыши, но не показали их в статье. Эти шипы должны быть задокументированы в документе (уровень LM), и, возможно, эти данные уже доступны.
Изображение ответа автора 1 представляет собой рисунок, показывающий шипы аксонов оливарного нейрона мыши с использованием процедуры разреженной вирусной маркировки (A).Область, где аксон ответвляется от сомы, показана в большом увеличении (B) и схематически изображены шипы аксонов (C). При необходимости этот рисунок может быть добавлен как дополнительный. На данном этапе мы не можем предоставить более подробное описание шипов аксонов, поскольку идентификация аксона в IO-клетках — непростая задача из-за обширных бифуркаций дендритов и тенденции аксонов выходить из дендрита.
3) Могут ли авторы предоставить более прямые доказательства того, что IRE происходят от шипов? Моделирования недостаточно, особенно когда мы даже не знаем, существуют ли аксонные шипы.Большинство людей считают, что спайки инициируются в начальных сегментах аксона из-за высокой плотности там Na-каналов, и есть хорошие маркеры для начальных сегмент-специфичных белковых комплексов (например, иммуно для анкирин-G или самих Na-каналов). Такой демонстрации в аксональных шипах (уровень LM) было бы достаточно. Лучшая электрофизиология (извлечение глутамата, запись аксонов) также была бы замечательной, но, вероятно, потребовала бы слишком много времени для проверки.
Как было сказано выше, мы работаем над этой проблемой, но в настоящее время не можем предоставить эту информацию.Мы надеемся, что демонстрации их существования будет достаточно для настоящего отчета.
4) Что касается функциональной значимости IRE, авторы никогда не упоминают, запускают ли эти большие подпороговые IRE события, распространяемые электронным путем (маленькие колоски?) В связанных клетках. Это данные, которые авторы уже должны иметь из своих парных записей. В общем, совсем не ясно, как IRE, колоски, спонтанные колебания и полные спайки взаимодействуют в связанных сетях нейронов IO; авторы должны представить более целостный взгляд на то, как взаимодействуют все эти мембранные события.
В рукописи мы специально сообщили, что при парной записи мы никогда не встречали коррелированного сигнала с IRE. Фактически, это было нашим основным побуждением к поиску альтернативной возможности. Если бы IRE происходили на дендритном уровне, они должны были привести к гораздо более высокому сигналу в пост-соединительном нейроне. Мы добавили более четкое описание этой проблемы в раздел «Результаты».
Мы добавили наш целостный взгляд на взаимодействие между этими событиями в Обсуждение.
5) Другая важная и потенциально интересная, но проблемная часть статьи касается колосков, сцепления и кластеризации. Авторы, возможно, смогут усилить это, не проводя дополнительных экспериментов, если они смогут решить некоторые технические проблемы, поднятые в обзорах (о ясности описания, порогах шума и обнаружения, альтернативных схемах подключения в модели, статистической надежности соответствия кривой моделирования. , и т.д).
Мы решили все технические проблемы, как было предложено рецензентами, перечисленными в каждом конкретном пункте ниже.
Рецензент № 1:
Есть несколько проблем:
— В статье два раздела: анализ подпороговых откликов и использование колосков для анализа сетевых подключений. Хотя первый раздел читается очень хорошо, у меня возникли проблемы с переходом к сетевому анализу. Я нашел два предложения в начале параграфа два подраздела «Оценка сетевой архитектуры на основе записей двух ячеек одновременно встречающихся колосков» очень запутанными.Пришлось читать их несколько раз. Существует мало информации о том, как общие амплитуда и соотношение колосков используются для создания групп. Хотя я понимаю эту идею, детали мне все еще неясны. Критерии и полезность этих значений для группировки колосков должны быть явными и не позволять читателю гадать.
Мы перефразировали эти предложения в разделе «Результаты» и в разделах «Материалы и методы».
— Связь между нижними оливарными нейронами неоднородна, и одна клетка может быть дифференциально связана со своими соседями.Я не уверен, как это повлияет на анализ, проведенный авторами, в частности на соотношение обычных колосков. Была ли учтена неоднородность связи в компьютерном моделировании? Авторы должны решить этот вопрос.
Мы не уверены, относится ли комментарий к неоднородной силе связи или к неоднородному количеству нейронов, которые связаны с каждым нейроном. В модели количество подключенных нейронов было неоднородным (рис. 8C). Проводимость каждого щелевого соединения была идентична, но положение на дендритном дереве, а также морфология и свойства мембран клеток были разными, что создавало неоднородность в колосках, как показано на рисунке 8B и на рисунке в ответ на комментарий рецензента №3 5 .
— Я предлагаю сделать заголовок специфичным для нижней оливы, возможно, «Использование подпороговых событий для характеристики функциональной архитектуры нижней оливы». Хотя количество связанных клеток не было неожиданностью и согласуется с предыдущими отчетами с использованием другой методологии, анализ, проведенный авторами, дает дальнейшее понимание организации сетей (способствует существованию компартментов функционально связанных нейронов), и это должно быть отражено. в названии.Кроме того, хотя это теоретически возможно, неясно, можно ли использовать этот метод для анализа других, менее известных сетей. Предлагают ли авторы этот метод применять в других сетях? Если да, им следует отдельно обсудить этот момент в разделе «Обсуждение».
Благодарим рецензента за предложенное название. Мы изменили название, чтобы быть более конкретным для низшей оливы: «Использование подпороговых событий для характеристики функциональной архитектуры электрически связанной сети низшей оливы».
Кроме того, актуальность нашего подхода к другим сетям была добавлена в последний раздел раздела «Обсуждение».
Рецензент № 2:
У меня есть несколько предложений по улучшению рукописи:
— Частота всплесков IO-ячеек была приблизительной, и, таким образом, это возможный источник систематической ошибки. Хотя авторы действительно говорят, что они оценили частоту возникновения спонтанных медленных событий, а не скорость всплеска зарегистрированных клеток, поскольку на них влияет внутриклеточный раствор, мне интересно, в какой степени оценки изменчивости и частоты частота обстрела клеток определяется записью, прикрепленной к клеткам.
Комментарий нам непонятен. Чтобы избежать артефактов пиковой активности из-за процедуры записи, мы оценили скорость возбуждения по частоте общих колосков, которые отражают активность в отдельных клетках, не нарушенных записью. В наших экспериментах мы не использовали записи с привязкой к клеткам.
— Что касается источника быстрых спонтанных событий, или «внутренних регенеративных событий (IRE)», авторы находят, что эти IRE не зависят от щелевых соединений, а происходят из шипов на аксонном холмике клетки.Я нахожу эту гипотезу вполне правдоподобной и привлекательной, но мне интересно, можно ли провести дополнительные электрофизиологические эксперименты, либо с извлечением глутамата, либо с применением глутамата или AMPA с помощью вторичной пипетки с «выдуванием». Это могло бы подтвердить эффекты этих «перекрывающих» аксональных шипов на нейроны IO. В частности, было бы неплохо увидеть эксперимент с прямой и / или одиночной стимуляцией аксональных шипов.
См. Ответ на пункт № 3 редактора.
— Авторы предполагают, что амплитуды IRE в основном определяются длиной шеи. Однако мне интересно, почему наблюдаемые амплитуды не распределены нормально, а имеют форму синусоиды (см. Рисунок 1B). Я бы предположил, что длины шеи позвоночника распределятся нормально. Если предположить, что более высокие амплитуды являются результатом суммирования, тогда мне интересно, почему синусоида такая чистая (потому что переменная длина шейки снизит точность формы синусоиды, поскольку отдельные события, составляющие суммированное событие, будут больше также переменная).Кроме того, как упоминается в тексте, De Zeeuw et al., 1990 показали, что до 8 шипов отходят от начального сегмента аксона. Не ожидали бы авторы большего разброса амплитуд, если бы это было так?
Действительно, квантовоподобное распределение — интересное наблюдение, однако оно наблюдалось не во всех ячейках. Изображение ответа автора 2 (A) представляет собой другой пример, в котором амплитуды распределены неравномерно.
Мы не предполагаем, что более высокие амплитуды являются результатом суммирования, поскольку это влечет за собой, что оба шипа иннервируются одним и тем же аксоном.Мы действительно наблюдали очень мало IRE, где четко наблюдался перерыв во времени нарастания (стрелка в B). Мы предполагаем, что это результат суммирования активности двух шипов. Однако эти IRE не были включены в анализ.
Мы не ожидаем большего разброса амплитуд, поскольку согласно De Zeeuw et al., 1990, обнаруживается до 8 шипов, некоторые из них получают только ГАМКергическую иннервацию, что не приводит к IRE.
— Учитывая, что медленные колоски преимущественно возникают поверх STO, учитывая, что потенциалы действия в одной клетке часто приводят к колоскам в электронно связанных клетках, и учитывая, что IRE часто индуцируют потенциалы действия, разве авторы не ожидали бы большего количества колосков за пределами нормальное СТО-колосок-окошко? Или авторы предполагают, что афференты к аксональным шипам каким-то образом также находятся под влиянием STO? Или эффекты ИРЭ зависят от фазы подпороговых колебаний? Если бы это было так, разве не была бы в некоторой степени затруднена функциональность возникающего индуктора потенциала действия, который исключает участие дендритов?
Как показано на Рисунке 2H (бывший Рисунок 3H), в отличие от колосков, частота IRE зависит от мембранного потенциала, таким образом, IRE с меньшей вероятностью может быть вызвано на дне STO.Более того, менее вероятно, что IRE вызовет потенциал действия во впадине колебаний. Таким образом, хотя кажется, что «афференты к аксональным шипам каким-то образом также влияют на STO», вероятно, это результат чувствительности этих событий к напряжению.
— Существует зависимость между основными колебаниями и количеством медленных колосков. Однако колоски будет труднее обнаружить в фазах крутого подъема колебаний. Как авторы справились с этим потенциальным препятствием?
Колоски в осцилляторных трассах были обнаружены после вычитания трассы, отфильтрованной нижними частотами, из необработанной трассы.Таким образом можно было легко обнаружить даже небольшие колоски на переднем и заднем фронтах. Мы добавили эту разборку в раздел «Материалы и методы» «Анализ данных и статистика».
— Распределение данных об амплитудах и времени, вероятно, ненормально, поэтому может быть лучше сообщать медианы и квартили, а не средние и стандартные отклонения по всей статье.
Согласно тесту нормальности Шапиро-Уилка, некоторые распределения нормальны или близки к норме, а соотношение медиана / среднее в некоторых случаях> 0.95 и в других> 0,9. Расчет медианы и квартилей дал следующие результаты, которые не сильно отличаются от среднего + стандартное:
Время быстрого нарастания событий 1,4 ± 0,4 мс (медиана 1,333)
Продолжительность 4,2 ± 1,3 мс (медиана 3,86)
Время нарастания медленных событий 2,5 ± 0,6 мс (медиана 2,4)
Продолжительность 12,7 ± 3,9 мс (медиана 12,6)
Таким образом, мы предпочитаем использовать описание mean + std, поскольку оно более читабельно. Если все еще нужно, мы с радостью заменим все номера.
— Мне не хватает графика распределения амплитуд STO, особенно в контексте утверждения, что щелевые переходы необходимы для спонтанных колебаний. Наше текущее понимание клеточных механизмов, генерирующих колебания, предполагает, что STO могут генерироваться в нескольких ячейках с высокой амплитудой, при этом другие клетки либо сообщают об этих колебаниях, либо действительно участвуют в них. Возможно, в сети, в которой ячейки соединены щелевыми соединениями, существует комбинация этих двух явлений.Фактически, Leznik and Llinas, 2004, показали, что STO поддерживаются в отдельных ячейках после блокировки щелевого соединения, хотя синхронизация сети нарушается. Так что, на мой взгляд, это комбинация, в которой уровень связи будет способствовать уровню STO, в то время как связь не является абсолютно необходимой.
— В рукописи создается впечатление, что клетки либо колеблются, либо нет, в то время как в литературе сообщается о промежуточных клетках «условных осцилляторах». Это важный момент, поскольку анализ динамической системы показывает, что неколеблющуюся ячейку можно быстро заставить колебаться при возмущении (Schweighofer et al., 1999).
— Хотя вполне вероятно, что колебания потенцируются связью, «колебания сети» в пару мВ могут быть просто считыванием осцилляторов высокой амплитуды. Пожалуйста, обсудите этот момент более подробно.
Мы озадачены этими комментариями. Действительно, механизм генерации STO в оливковом ядре обсуждается, но это не является предметом настоящего исследования. В анализе, показанном на рисунке 6 (бывший рисунок 2), где использовались только стабильные колеблющиеся клетки, мы использовали появление колосков как показатель того, что в этой электрически связанной сети все связанные нейроны колеблются с одинаковой частотой, что согласуется с различные взгляды на источник колебаний IO и далее не обсуждаются.
— Было бы неплохо увидеть симуляцию с двумя альтернативными сценариями подключения. Один с меньшим количеством соединений, другой с большим — последнее отражает возможный сценарий в неповрежденном препарате.
Мы не уверены, полностью ли понимаем комментарий. В ответ на комментарий 6 рецензента №3 мы представляем больше сценариев подключения.
Рецензент № 3:
Эксперименты и моделирование были выполнены хорошо, анализ интересен, а выводы исследования потенциально важны.Если небольшие шиповидные события действительно происходят из шипообразных структур на начальном сегменте аксона, это было бы новым и уникальным наблюдением, насколько мне известно. Если электрические колоски, генерируемые синапсами, можно будет использовать для надежной оценки размера и структуры кластеров нейронов, связанных щелевыми соединениями, это станет важным вкладом в нейробиологию IO и, возможно, в более широком смысле. Однако, как здесь представлено, я не думаю, что ни один из выводов убедительно подтверждается доказательствами.
1) Это исследование полностью основано на свойствах небольших подпороговых событий, поэтому четкое описание их характеристик имеет решающее значение.Вопрос первого порядка заключается в том, представляет ли какое-либо из этих событий химический синаптический ввод. Они ВПСП? Они вызваны ВПСП? Ответы здесь расплывчаты. Авторы сообщают, что применение CNQX снизило частоту «унитарных униполярных событий» в среднем с 0,7 Гц до «ниже 0,02 Гц» (параграф 1 результатов), но никаких подробностей не приводится. Эта контрольная скорость, по-видимому, была взята из большой группы клеток, а не из 19 клеток, на которые было нанесено лекарство. Авторы должны правильно сообщить данные с измерениями одних и тех же нейронов до и после CNQX.Они также должны сообщать характеристики подпороговых событий, оставшихся после CNQX, включая амплитуды и кинетику, и сравнивать их с событиями из тех же клеток в отсутствие лекарственного средства. Позже авторы говорят, что быстрые события «не зависят от химической передачи», но они не приводят никаких доказательств этого. Все это связано с ключевым вопросом: какой вклад ВПСП вносят в подпороговые события и их свойства?
См. Развернутый ответ на комментарий редактора 1.
2) Все ли данные на Рисунке 1 были записаны в отсутствие CNQX? Если ответ положительный, и CNQX снизил частоту этих подпороговых событий на 97% (см. Предыдущий комментарий), то не следует ли ожидать, что большая часть подпороговых событий будет спонтанными ВПСП, а не колосками?
Да, CNQX не использовался ни в одном из нейронов, представленных на Рисунке 1. Уменьшение количества колосков под действием CNQX объясняется уменьшением возбуждения в сети.Мы добавили новый дополнительный рисунок (рисунок 2 — рисунок в приложении 1) и обсудили проблему далее в разделе «Результаты».
3) Запись и анализ подпороговых событий ограничены порогами обнаружения и отношениями сигнал / шум. Некоторые из событий (колоски и, возможно, IRE), по-видимому, слишком малы, чтобы их можно было увидеть. Это имеет значение для выводов, которые можно сделать из записей, включая оценки размера кластера связанных нейронов. Уровни шума здесь не сообщаются.Моделирование авторов может предоставить оценки пределов их обнаружения с учетом ограничений отношения сигнал-шум и, возможно, поддержать надежность их кластерных оценок. Авторы должны обсудить этот вопрос.
Мы согласны с тем, что анализ ограничен отношением сигнал / шум. Действительно, мы обсуждали, что наши результаты, вероятно, будут недооценкой количества колосков и общих групп. Однако это ограничение могло только существенно повлиять на наш вывод о том, что подключение организовано в кластеры.Чтобы проверить, будет ли это препятствовать нашей способности отвергать конфигурацию, зависящую от расстояния, мы уменьшили вдвое (6 пар) количество пар без общих групп и равномерно распределили эти 6 пар в общих группах (см. Изображение ответа автора 3). Эти измененные данные по-прежнему отвергают модель, зависящую от расстояния (значение A, p <0,002), и поддерживают организацию кластера (значение B, значение p> 0,05). Кроме того, следует отметить, что в случае 12 пар, не имеющих общих групп, мы тщательно проверили, что колоски в одной ячейке никогда не совпадают с каким-либо изменением напряжения в другой ячейке.
4) Как говорят авторы (дважды), происхождение IRE «остается загадкой». Доказательства того, что маленькие колоски являются постсинаптическим результатом пресинаптического потенциала действия, проходящего через электрические синапсы, весьма убедительны. Дымящееся ружье взято из записей парных клеток, которые были продемонстрированы в различных лабораториях много лет назад.Однако доказательства того, что более крупные и быстрые «IRE» возникают в шипах аксонов, прилегающих к соме, являются слабыми, предполагаемыми и косвенными.
Результаты моделирования IRE наводят на размышления, но я согласен с авторами в том, что «места и механизм генерации IRE полностью не выяснены». Фактически, без дополнительных экспериментальных данных я не считаю эту часть истории убедительной. Это, конечно, не соответствует преувеличению в абстракции: «быстрые события представляют собой регенеративную реакцию в уникальных возбудимых шипообразных структурах в бугорке аксона».Во-первых, неясно даже, существуют ли аксональные шипы в IO клетках мыши. Если да, то их следует проиллюстрировать. Авторы ссылаются на оригинальные данные об аксональных шипах от De Zeeuw, но эта работа была проведена на кошачьих нейронах. Во-вторых, некоторые более прямые доказательства местонахождения генерации IRE помогли бы сделать вывод убедительным, например прямые записи с начальных сегментов аксона или шипов с использованием электродов или фотоиндикаторов, или молекулярные / структурные демонстрации натриевых каналов высокой плотности в шипах аксонов.
Мы полностью согласны с тем, что источник и механизм IRE далеко не убедительны и потребуют тщательной работы, которой мы сейчас занимаемся. Однако, чтобы решить, следует ли включать или исключать IRE из анализа организации электрически связанной сети, мы должны были предоставить достаточно указаний на то, что они не представляют потенциалы электрической связи. Мы добавили предварительное наблюдение о существовании аксональных шипов в IO-клетках мыши и изменили формулировку в аннотации.См. Наш подробный ответ на комментарий редактора №2.
5) Анализ групп «общих колосков» в парных записях (рис. 7) как способ сделать вывод о размере электрически связанных кластеров клеток интересен и умен. Однако мне интересно узнать об ограничениях данных и их значении для оценок размера кластера. Проиллюстрирован один пример пары, из которой были заявлены четыре группы общих колосков (рис. 7E). Амплитуды колосков, конечно, небольшие и изменчивые, и выборки общих (и необычных) колосков в каждой ячейке также были небольшими (всего два в примере пары).В материалах и методах нет четкого определения статистической надежности этой формы кластерного подсчета. Например, два кластера колосков справа на рис. 7E для меня не сильно отличаются, особенно с учетом того, что образцы такие маленькие, а размеры / формы колосков очень разные. Распределение размеров колосков на Рисунке 1 также подразумевает большую и непрерывную дисперсию (а не пиковые распределения, показанные для IRE). Слабые критерии кластеризации колосков могут привести к завышению размеров электрически связанных кластеров.Показанная пара ячеек (рис. 7E) имеет одно из наименьших количеств предполагаемых соединений (рис. 7F). Кажется, проблема определения отдельных общих кластеров колосков (и оценки связей) также должна возрастать по мере увеличения количества кластеров в ячейке. Этот вопрос требует более строгого обоснования и обсуждения.
Мы согласны с рецензентом в том, что неправильная кластеризация колосков будет препятствовать нашей оценке связанных ячеек, и по этой причине мы приложили все усилия, чтобы разработать анализ, который будет правильно кластеризовать трассы.Верно, что, как показано на рисунке 1, распределение амплитуд колосков велико. Однако если взять 3 параметра: амплитуда общего колоска в первой ячейке; амплитуда общего колоска во второй ячейке; Разница между двумя амплитудами дает очень четкие скопления колосков.
И в экспериментальной, и в моделирующей части мы изначально использовали алгоритм кластеризации DBSCAN для кластеризации общих колосков в соответствии с их пиковыми амплитудами и различиями между амплитудами.Однако, хотя алгоритм хорошо работает для модели, он менее идеален для зашумленных экспериментальных трасс, хотя количество групп, обнаруженных алгоритмом, существенно не отличалось от ручного курирования. На изображении ответа автора 4 мы сравниваем ручное курирование с результатами DBSCAN в другой паре примеров. В то время как схожесть двух режимов анализа отчетливо видна, алгоритм предсказал 7 групп, тогда как ручной анализ привел к 6 группам. Во-первых, очевидно, что ручное курирование более точное.Во-вторых, как мы продемонстрировали выше (ответ на комментарий 3), небольшое изменение количества общих групп не повлияло на значимость нашего утверждения.
В примере, показанном на рисунке 7, два правых кластера действительно имеют похожие черные следы, но красные следы можно легко сгруппировать в два разных кластера в соответствии с их амплитудами. Мы выбрали этот пример, поскольку считаем, что это простой пример для объяснения нашего методологического подхода.
Мы добавили более четкое описание анализа в раздел «Материалы и методы».
Результаты моделирования на Рисунке 8 в некоторой степени проверяют анализ, показанный на Рисунке 7, путем изменения периода выборки и частоты срабатывания. Однако смоделированные данные выглядят гораздо менее изменчивыми (по амплитуде), чем реальные данные (например, см. Рисунок 8B с 7B и E). Из описаний я также делаю вывод, что в модельной сети не было шума.
Действительно, в примере, приведенном на Рисунке 8, есть небольшие различия, однако это не было общей чертой, как показано в другом примере на изображении ответа автора 5. Разница в амплитуде колосков возникла из-за разницы в расположении GJ и морфология клеток и свойства мембран. Также был шум, возникающий из-за разницы в средней скорости срабатывания ячеек, эффект, который мы количественно оценили на Рисунке 8 — добавлении к рисунку 1. Шум не добавлялся к графикам напряжения.
6) Моделирование на Рисунке 8 не обеспечивает убедительной поддержки гипотезы авторов о кластеризации связности щелевых соединений по сравнению с простой зависимой от расстояния моделью связности. Похоже, это сводится к еще одному сравнению оценок, полученных в результате анализа колосков на Рисунке 7, с результатами модели (Рисунок 8).В частности, авторы приходят к выводу, что модель расстояния «не смогла воспроизвести распределение общих групп, обнаруженное экспериментально (рис. 8F)», тогда как модель кластеризации «воспроизвела зависящую от расстояния вероятность соединения (рис. 8H)». На мой взгляд, посадки на рис. 8F и H кажутся примерно одинаково хорошими (или плохими). Интересно, что авторы приходят к выводу, что соответствие измерениям данных на рис. 8E и H примерно одинаково хорошо, но, на мой взгляд, соответствие на рис. 8H выглядит примерно так же хорошо (или плохо), как и на рис. 8F и H.Другими словами, результаты моделирования кажутся очень слабым доказательством выбора между этими сценариями кластерной зависимости или сценариями зависимости от расстояния.
В этом отношении может показаться, что экспериментальные исследования связывания индикаторов между IO-клетками (как у Devor and Yarom, 2004; Placantonakis et al., 2006; Hoge et al., 2011) на тех же срезах, что и анализируемые с помощью Общий метод колосков был бы более надежным способом определить, является ли электрическая связь случайной / зависимой от расстояния или определяется кластеризацией.
Спасибо за комментарий. Мы провели полный статистический анализ по этому вопросу, который обсуждается в новом разделе «Материалы и методы» и на дополнительном рисунке (Рисунок 8 — рисунок в приложении 2).
Короче говоря, мы добавили значение p для рисунков 8F и 8I (<0,002 для 8F и 0,907 и 0,125 для черного и серого 8I, соответственно). А также для рисунков 8E и H (все> 0,05).
Кроме того, в исходной рукописи мы протестировали общее групповое распределение только для одной конфигурации связи, зависящей от расстояния.Здесь мы подтвердили, что наши результаты не ограничиваются только этой конкретной дистанционно-зависимой конфигурацией, на что также указали два других рецензента. На изображении ответа автора 6 представлен анализ связности сетей с различной вероятностью связности, зависящей от расстояния (слева), в котором Σ и σ (см. Материалы и методы, матрицы связности) различны, а также соответствующее распределение общих соседей (справа). Для всех этих конфигураций подключения значения p для общего распределения соседей были <0.05.
7) Авторы обсуждают появление «обычных колосков», но не говорят много о IRE и электрическом взаимодействии. Что происходит в электрически связанной ячейке, когда IRE возникает в парной ячейке? Можно ли вообще обнаружить эти события? В Обсуждении просто говорится: «IRE в одном нейроне никогда не совпадало с колоском в другом нейроне», но я не верю, что в статье говорится, что действительно совпадает с IRE.
Мы не наблюдали никаких событий или изменений в мембранном потенциале в одной клетке, в то время как IRE был очевиден в другой клетке, а также мы не видели IRE в одной клетке из-за всплеска в другой клетке. Эти данные сильно подтверждают гипотезу о том, что IRE не вызываются в дендритах, так как они будут проходить через щелевые соединения к связанным нейронам. Это действительно была наша причина искать другую возможность происхождения IRE после того, как мы отклонили возможность прямых-косвенных колосков, которая представлена на рисунке 4 — рисунок в приложении 1.Этот момент был подробно рассмотрен в разделе «Результаты» и «Обсуждение».
[Примечание редакции: до принятия были предложены дальнейшие исправления, как описано ниже.]
Основное изменение текущего представления — полное удаление аксональных шипов и сосредоточение внимания на организации сети.
В текущей версии мы также ответили на комментарии рецензента:
a) Провел новую серию фармакологических экспериментов, показавших, что быстрые события блокируются синаптическими блокаторами, тогда как колоски все еще присутствуют и, следовательно, могут отражать электрическую связь.Как отмечали обозреватели, тот факт, что частота колосков значительно снижается под действием лекарств, может предполагать, что некоторые из медленных подпороговых событий могут быть химическими синаптическими событиями, что может поставить под сомнение использование спонтанных событий для изучения сетевой организации. . Чтобы решить эту проблему, мы сначала сравнили спонтанные колоски с вызванными (в парных записях) и показали их сходство. Во-вторых, мы проанализировали новую серию экспериментов с вызванными светом химическими синапсами у мышей Thy1, продемонстрировав, что их форма волны и амплитуда отличаются от спонтанных и вызванных медленных событий.Таким образом, мы заключаем, что большая часть, если не все, спонтанных медленных событий действительно происходит из-за электрической связи.
Кроме того, мы обсуждаем снижение активности колосков в условиях приема лекарств, утверждая, что спонтанная пиковая активность запускается быстрыми событиями и что блокирование быстрых событий приводит к снижению частоты колосков.
b) Мы изучаем роль шума в обнаружении общих групп колосков, предполагая либо отказ, либо чрезмерное обнаружение. Это показано на Рисунке 7.Мы также добавили моделирование ожидаемого распределения общих групп при различных предположениях. Во всех этих симуляциях довольно ясно, что сеть, в которой вероятность соединений зависит только от расстояния, не может учитывать экспериментально наблюдаемое распределение. Таким образом, главный вывод этого исследования о том, что ядро нижней оливы организовано в группу связанных клеток, полностью подтверждается экспериментальными наблюдениями.
На наш взгляд, убедительно то, что мы предлагаем новый экспериментальный и теоретический подход к изучению электрически связанных сетей, и надеемся, что он найдет ваше одобрение.
Рецензент № 3:
Авторы устранили некоторые недостатки оригинальной рукописи. Однако я все же считаю, что есть серьезные недостатки. На мой взгляд, аргумент о том, что IRE, вероятно, происходят из шипиков аксонов, неубедителен. Анализ и моделирование колосков — более сильная история, но моделирование не учитывает эффекты шума в клетках. Кроме того, фармакологические результаты все еще описываются неправильно.
1) Истоки IRE и отростки аксонов.Авторы несколько смягчили свой провокационный вывод о том, что IRE генерируются в аксональных шипах, хотя в аннотации все еще говорится: «Мы предполагаем, что быстрые события представляют собой регенеративную реакцию в уникальных возбудимых шипообразных структурах в аксонном бугорке». На мой взгляд, данные, подтверждающие это предположение, все еще очень слабы по следующим причинам:
A) Доказательства авторов даже о существовании аксональных шипов в нейронах мыши неубедительны. В ответе авторов на обзоры было представлено только одно изображение одного нейрона IO мыши (изображение ответа автора 1).Разрешение изображения низкое, и каждый предполагаемый аксональный отросток, кажется, представлен небольшим количеством пикселей. Увеличенное изображение на изображении ответа автора 1B включает пунктирный контур (нарисованный от руки?), Который представляет собой чрезмерно оптимистичную интерпретацию пикселей, а рисунок в изображении ответа автора 1C является просто карикатурной версией изображения ответа автора 1B. Я понимаю, что визуализация этих небольших структур технически сложна, но, не зная, распространены ли шипы, насколько они велики, где они расположены и насколько хорошо они коррелируют с IRE, вывод о том, что IRE генерируются аксонными шипами, просто не убедительно.
B) Авторы не предоставили никаких дополнительных морфологических, молекулярных или электрофизиологических доказательств, которые помогли бы связать происхождение IRE с шипами аксонов (как они ответили: «… мы работаем над этой проблемой, но в настоящее время мы не можем предоставить эту информацию. Мы надеюсь, что демонстрации их существования достаточно для настоящего отчета ».) Но их рукопись выходит далеко за рамки простой демонстрации существования или IRE. Размышления об экзотических механизмах должны подождать, пока не появятся подтверждающие доказательства.
C) Авторы смоделировали несколько возможных механизмов генерации IRE. Сначала они показали, что IRE-подобные события могут быть сгенерированы путем моделирования «горячих точек» возбудимости в дендритах или, альтернативно, путем моделирования сбоев спайков во все более удаленных аксональных узлах. Затем они отклонили эти возможности, заявив, что «трудно представить себе биологический механизм, который либо конкретно локализует каналы в ограниченной дендритной« горячей точке », либо одновременно блокирует два, три или более Узлов Ранвье» (четвертый абзац, подраздел «Моделирование Внутренние регенеративные события (IRE) »).Возможно, так, хотя две статьи, цитируемые авторами, демонстрируют доказательства относительно высокой плотности натриевых каналов в дендритных шипах (Araya et al., 2007; Bywalez et al., 2015). Мне так же трудно представить себе горячие точки ионных каналов в биологически уникальных аксональных шипах, которые не были четко продемонстрированы ни в изучаемых оливковых клетках, ни, действительно, в любом другом классе нейронов позвоночных.
D) Моделирование генерации спайков в предполагаемых аксональных шипах (Рисунок 5) исследовало очень ограниченное и биологически неоправданное пространство параметров.Авторы включили высокие плотности натриевых и калиевых каналов (идентичные таковым в начальном сегменте аксона) как в головках позвоночника, так и в бугорке аксона, в то время как возбудимость аксональных узлов Ранвье была фактически устранена (возбудимый бугорок и непоколебимый бугорок). аксоны упоминаются только в Материалах и методах, а не в основном тексте или легенде). Консенсус в этой области состоит в том, что плотности каналов в бугорках аксонов нейронов позвоночных довольно низки, особенно по сравнению с начальным сегментом.Возможно, оливковые клетки мыши не похожи на другие нейроны, но в отсутствие доказательств мы просто не знаем. Что послужило основанием для того, чтобы сделать остальную часть аксона совершенно неуправляемой при моделировании возбудимых шипов? В рукописи проиллюстрированы лишь некоторые результаты этого моделирования; насколько надежны эти результаты? Каковы последствия различной плотности и типов каналов, их распределения, пространственного рисунка, морфологии и количества шипов и т. Д.?
Как упоминалось выше, эта часть была удалена.
2) Сетевое моделирование и отсутствие шума. Самый новый и интересный вывод из сетевого моделирования состоит в том, что ячейки ввода-вывода могут быть организованы в электрически связанные кластеры ячеек (прогнозы вероятности соединения в значительной степени согласуются с широко варьируемым диапазоном, предложенным предыдущими исследованиями).
Спасибо. Мы, безусловно, согласны с этим описанием.
Заключение о кластеризации полностью основывается на совпадении распределений «общих колосков» в записанных парах ячеек с предсказаниями сетевой модели.Авторы отмечают, что точность смоделированных распределений соединений сильно зависит от частот срабатывания ячеек и длины записывающих образцов. Не должно ли это также зависеть от шума? Важной особенностью биологического препарата, отсутствующей в модели, является любой источник шума или изменчивости (кроме пуассоновского времени соматических токов, запускающих всплески), особенно в подпороговых мембранных напряжениях.
Мы изучаем роль шума в обнаружении общих групп колосков, предполагая либо отказ, либо чрезмерное обнаружение.Это суммировано на новом рисунке 7, показывающем, что даже если мы предположим, что обнаружение общей группы не удалось или неточность в определении количества общих групп, кластерная организация по-прежнему остается единственным возможным объяснением. Кроме того, мы исследовали несколько возможностей связи, зависящих от расстояния, и ни одна из них не дала распределения общих групп, которое соответствует экспериментальным результатам. Таким образом, мы можем заключить, что единственное объяснение наблюдаемого распределения — это кластерная организация.
Записи авторов подразумевают, что блокирование быстрых рецепторов глутамата резко снижает частоту спонтанных всплесков, поэтому можно сделать вывод, что обычно существует значительный химический синаптический шум в дополнение к другим потенциальным источникам.
Как упоминалось выше, снижение активности колосков сильно коррелирует со снижением быстрых событий, которые в IO запускают пиковую активность (рисунок 3 — приложение к рисунку 2). Таким образом, блокирование быстрых событий приведет к снижению пиковой активности с последующим уменьшением колосков.Синаптические потенциалы, запускающие быстрые события, не могут быть записаны в соматическом месте. Спонтанные синаптические события встречаются редко. Однако, чтобы изучить возможное участие химических синапсов в спонтанной активности, мы проанализировали синаптические события, которые запускались активацией ChR возбуждающих аксонов, проецируемых на нейроны IO у мышей Thy1. Используя минимальную продолжительность освещения, мы смогли активировать предположительно унитарные события. Затем мы сравнили формы сигналов синаптических потенциалов с формами сигналов медленных событий.Как показано на Рисунке 3 (приложение 1 к рисунку), форма волны и амплитуда синаптических потенциалов явно отличаются от спонтанного медленного события (n = 17 ячеек). Кроме того, мы демонстрируем (рис. 2), что колоски нечувствительны к мембранному потенциалу. Это, которое было исследовано на большой популяции нейронов, дополнительно подтверждает наш вывод о том, что большинство, если не все, медленных событий являются отражением электрического взаимодействия. Можно утверждать, что нечувствительность к мембранному потенциалу может быть связана с дистальным расположением синапса, однако форма колосков не поддерживает такую возможность.
3) Фармакология. Фармакологические данные (синаптические блокаторы) все еще описываются запутанно и не очень полезны. Из результатов, подраздел «Спонтанные единичные события, зарегистрированные в нейронах нижней оливы»: «Однако применение синаптических блокаторов (см. Материалы и методы) полностью исключило наличие быстрых событий (n = 19 нейронов; в 4 из этих нейронов. где CNQX был добавлен во время записи, частота изменилась с 0,017 ± 0,005 Гц до 0 Гц), тогда как частота спонтанных медленных событий значительно снизилась (с 0.92 ± 0,73 Гц от 163 до 0,26 ± 0,16 Гц, p = 0,019, парный t-критерий, n = 9 нейронов) «Мои вопросы:
А) Какие были блокираторы? В этом предложении результатов указано только CNQX, в легенде к рисунку 2 — в дополнении к рисунку 1 указано APV плюс CNQX или DNQX, а в материалах и методах просто перечислены все препараты.
B) Размеры протестированных выборок все еще неоднозначны. Во фразе о быстрых событиях говорится, что n = 19, но затем говорится о n = 4, «где был добавлен CNQX». Не добавляли ли препарат в остальные 15 клеток? Во фразе о медленных событиях приводится n = 9.Были ли протестированы одни и те же клетки до и после добавления блокаторов? Являются ли 4 и 9 ячеек подмножествами 19 или разными выборками? Просьба уточнить.
C) Авторы говорят, что они провели «тщательный анализ» эффектов блокаторов на формы волны медленных событий, но на самом деле они сообщают данные только из двух примеров ячеек на Рисунке 2 — приложение к рисунку 1. Эти данные показали время нарастания и половинное время. длительности, но не амплитуды.
Мы добавили новую когорту данных и упростили описание эффекта препарата.Как упоминалось выше, мы показываем, что препарат полностью устраняет быстрые события, сопровождающиеся значительным снижением активности колосков. Это было добавлено к тексту и к рисунку 3 — добавление к рисунку 1, и мы надеемся, что теперь это понятно.
https://doi.org/10.7554/eLife.43560.sa2
(PDF) Подпороговый кольцевой осциллятор со сверхнизким энергопотреблением
14992 Abhishek Kumar
В современных цифровых КМОП интегральных схемах преобладает динамическая мощность
.Для снижения энергопотребления лучше всего снизить напряжение питания
. Для проведения МОП напряжение затвора должно быть больше Vth.
GS th
GS th
V V NMOS OFF
V NMOS ON
VGS , известному как подпороговый ток. В подпороговых схемах напряжение питания понижается до ниже порогового напряжения транзистора.Из-за квадратичного уменьшения мощности по отношению к питающему напряжению подпороговые цепи классифицируются как сверхмалые цепи . Подпороговая схема КМОП аналогична стандартной схеме . Это привлекает внимание к конструктору cmos малой мощности. Подход с подпороговым значением жертвует скоростью ради мощности, используя токи утечки для управления логическими вентилями. Пороговое значение Volateg Engineering Пороговое напряжение — один из наиболее важных параметров полевого МОП-транзистора, минимальное напряжение , необходимое для создания инверсии канала.Если напряжения смещения затвора недостаточно для инвертирования поверхности VGS , называемый допороговым током. Поддерживаемое напряжение питания подпороговой цепи на ниже порогового напряжения. В данной исследовательской работе напряжение питания находится в диапазоне от 0,1 до 0,5 В. требуется, чтобы Vth был намного ниже. Пороговое напряжение можно регулировать путем селективной имплантации легирующих ионов в область канала.Корпус влияет на пороговое напряжение , когда он не привязан к источнику, его можно рассматривать как задний затвор. Порог NMOS увеличивается за счет добавления дополнительных примесей p-типа через оптовую часть терминала. Положительное напряжение тела увеличивает порог NMOS, в то время как отрицательное напряжение тела приводит к снижению порога. Порог MOS задается хорошо известным уравнением смещения [6] 0 F SB | 2 В | | 2 | th t F VV Vt0 — пороговое напряжение для порога нулевой подложки, γ — эффект смещения подложки. Положительное напряжение тела имеет тенденцию к увеличению порогового напряжения PMOS в то же время , когда оно снижает пороговое напряжение NMOS. Аналогичным образом отрицательное напряжение тела имеет тенденцию к снижению порогового напряжения PMOS и увеличению порогового напряжения NMOS. Прямое смещение корпуса снижает пороговое напряжение, а обратное смещение увеличивает пороговое напряжение [4]. Метод смещения тела — лучший выбор для подпороговой конструкции.Он обеспечивает быстрое переключение и эффективное отключение устройств на холостом ходу в пороговом режиме ниже . Существует 3 способа подачи напряжения корпуса на инвертор , а именно: обычная КМОП, КМОП с динамическим порогом и смещение переставленного тела КМОП, показанное на рисунке 1 [3] В стандартной конфигурации большая часть NMOS и PMOS привязана к клемму заземления и напряжение питания (VDD) для преобразователя CMOS соответственно.