
Повторно-кратковременный режим — Энциклопедия по машиностроению XXL
Повторно-кратковременный режим. Этот режим характеризуется наличием периодически повторяемых процессов торможения и пауз в работе тормоза. Период охлаждения (пауза) относительно невелик, и температура поверхности трения не успевает снизиться до температуры окружающей среды, поэтому каждое последующее торможение начинается при температуре более высокой, чем начальная температура предыдущего торможения. По мере увеличения температурного перепада между температурой элементов тормоза и температурой окружающей среды увеличивается количество тепла, отдаваемого в окружающую среду, и рост температуры поверхности трения замедляется. После некоторого числа торможений количество тепла, отводимого в окружающую среду, становится равным количеству тепла, образующемуся при торможении, и создается некоторое условное
[c.591]
ПКР означает повторно-кратковременный режим работы, выражающийся в процентах как отношение времени горения дуги к времени работы аппарата при 5-минутном цикле. [c.345]
[c.345]
Пример 2. Рассмотрим повторно-кратковременный режим работы тормоза. Рассчитаем объемную н поверхностную температуры после пяти торможений. [c.198]
Повторно-кратковременный режим работы [c.283]
Повторно-кратковременный режим, при котором кратковременные рабочие периоды чередуются с кратковременными паузами. Этот режим характеризуется относительной продолжительностью включения ПВ, представляющей собой отношение времени работы tp ко времени цикла t . [c.228]
Агрегаты вспомогательного привода должны рассчитываться на повторно-кратковременный режим работы. Продолжительность включения определяется по типовым нагрузочным режимам. [c.131]
Плазменная резка 311 Плазменная сварка 8, 233 Плазмообразующие сопла 230 Плазмообразующий газ 223, 225 Плазмотрон 223 Пластические деформации 37 Пневматические испытания 358 Поверхностный эффект 264 Повторно-кратковременный режим источника питания дуги 94 Подогреватель газа 161 Покрытия электродов для ручной дуговой сварки 113, 115 Полуавтомат сварочный 141, 164 Полярность сварочной дуги 85 Порошковое копьё 310 Поры 338
[c. 393]
393]
Редукторы рассчитаны на два режима работы повторно-кратковременный режим ПВ = 25 /о и непрерывный режим ПВ = 100 /о для положения червяк под колесом . Для верхнего или бокового положения червяка расчетная мощность должна быть снижена на 20—25 /о. [c.520]
Повторно-кратковременный режим Режим работы, при котором периоды включения регулярно чередуются с периодами выключения ПК [c.87]
Повторно-кратковременный режим представляет собой длительно-повторяющиеся циклы. В каждом цикле последовательно чередуются включение — работа, выключение — пауза. Характеризуется этот режим продолжительностью включения ПВ, выражаемой в процентах [c.341]
Повторно-кратковременный режим работы отличается частыми пусками и остановками, при которых за время работы двигателя tj его температура не успевает достигнуть установившегося значения, а за время остановки tr=o не успевает опуститься до температуры окружающего воздуха. Продолжительность одного периода tj + ir=o Продолжительность включения двигателя (%) характеризуется фактической величиной
[c.384]
Продолжительность одного периода tj + ir=o Продолжительность включения двигателя (%) характеризуется фактической величиной
[c.384]
Существует два типовых режима сварки. При ручной электродуговой сварке (а также автоматической и полуавтоматической на постоянном токе) используется повторно-кратковременный режим, при котором время сварки чередуется с временем работы источника питания на холостом ходу. Такой режим оценивается по относительной продолжительности работы [c.51]
Относительная продолжительность работы (ПР) и относительная продолжительность включения (ПВ) в прерывистом режиме характеризуют повторно-кратковременный режим работы источника питания.
[c.136]
Повторно-кратковременный режим работы характеризуется чередованием работы двигателя и паузами, когда двигатель отключен от сети, причем за время работы двигатель не нагревается до установившейся температуры, а за время паузы не охлаждается до температуры окружающей среды. Основной характеристикой повторно-кратковременного режима работы является относительная продолжительность включения ПВ%. Для двигателей повторно-кратковременного режима установлены стандартные значения ПВ, равные 15, 25, 40 и 60%. Причем время одного цикла не должно превышать 10 мин. Если время
[c.64]
Основной характеристикой повторно-кратковременного режима работы является относительная продолжительность включения ПВ%. Для двигателей повторно-кратковременного режима установлены стандартные значения ПВ, равные 15, 25, 40 и 60%. Причем время одного цикла не должно превышать 10 мин. Если время
[c.64]
В связи с этим рекомендуется за критерий принять произведение эквивалентных величин q Va, которое учитывает переменность нагрузки и повторно-кратковременный режим работы подшипников. Ниже дается определение эквивалентных величин дэ и Уэ для различных цапф механизмов. [c.45]
Повторно-кратковременный режим [c.28]
Повторно-кратковременный режим крановых электродвигателей характеризуется относительной продолжительностью включения, выраженной в процентах, а именно [c.354]
Основным номинальным режимом является повторно-кратковременный режим с относительной продолжительностью включения ПВ=25%. В табл. 69—76 приведены также технические данные для режимов работы ПВ=15, 40, 60%, для длительного режима работы (ПВ=100 /о) и для кратковременного режима работы продолжительностью 60 мин.
[c.354]
В табл. 69—76 приведены также технические данные для режимов работы ПВ=15, 40, 60%, для длительного режима работы (ПВ=100 /о) и для кратковременного режима работы продолжительностью 60 мин.
[c.354]
Под режимом работы понимается соотношение между временем /ев сварки и временем холостого хода (перемежающийся режим) или временем паузы в случае, если источник питания отключается от сети (повторно-кратковременный режим). Во время холостого хода или паузы выполняют смену электродов, сборку заготовок, очистку шва от шлака и других загрязнений, происходит также охлаждение источника питания. [c.111]
Повторно-кратковременный режим характеризуется продолжительностью включения ПВ (%), для определения которой используют выражение [c.111]
Повторно-кратковременный режим — режим работы электродвигателя, при котором в период включенного состояния температура частей двигателя не успевает достигнуть установившихся значений, а в период отключенного состояния части дви,-гателя не успевают остыть до температуры окружающей среды. [c.176]
[c.176]
Повторно-кратковременный режим используют при работе сварочными полуавтоматами. Постоянный рабочий режим (ПВ = 1бО %) используют для установок автоматизированной сварки или для автоматов. [c.45]
Для снижения искрения параллельно контактам 6 прерывателя подключен конденсатор. Сирена рассчитана на повторно-кратковременный режим работы. Продолжительность непрерывного сигнала свыше 8 мин по температурному режиму недопустима. Номинальное напряжение сирены 110 В, но она включена на тепловозе в цепь напряжением 75 В. Катушки намотаны проводом ПЭЛ 0,15 400 витков, сопротивление 240+12 Ом, напряжение ПО В, потребляемая мощность 60 Вт, громкость звукового сигнала (при ПО В) 105 дБ. [c.196]
Конструкция электросверлилки рассчитана на повторно-кратковременный режим работы. Иными словами, сверлилка не может работать длительное время без выключения, так как это может вызвать перегрев обмоток двигателя и выход его из строя.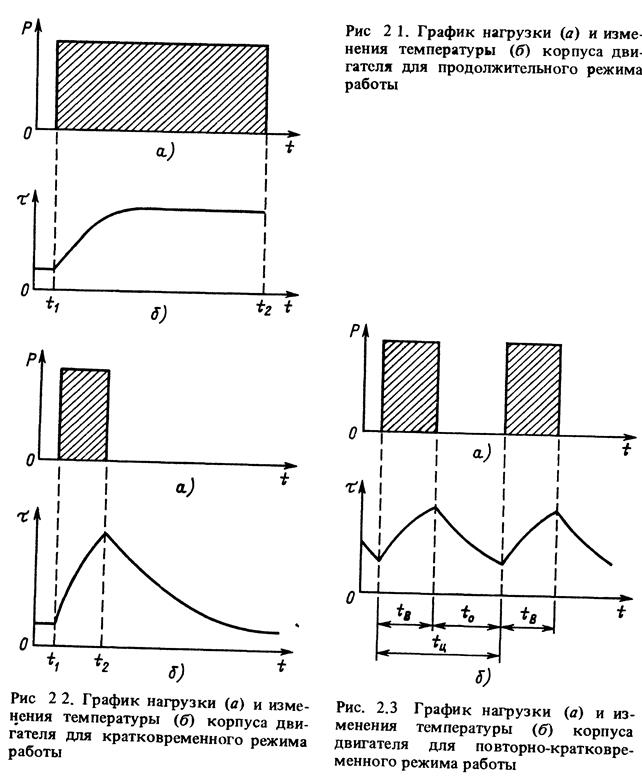 Цикл работы сверлилки состоит из рабочего периода, когда производится сверление, и перерыва, в течение которого сменяют инструмент или рабочий отдыхает. Период работы и перерыв составляют вместе цикл. Отношение рабочего периода к циклу, выраженное в процентах, характеризует собой повторно-кратковременный режим работы (ПКР). Для сверлилки И-38 нормальный ПКР равен 60%. Для ряда других сверлилок ПКР равняется 40%. Эта характеристика указывается в паспорте инструмента.
[c.78]
Цикл работы сверлилки состоит из рабочего периода, когда производится сверление, и перерыва, в течение которого сменяют инструмент или рабочий отдыхает. Период работы и перерыв составляют вместе цикл. Отношение рабочего периода к циклу, выраженное в процентах, характеризует собой повторно-кратковременный режим работы (ПКР). Для сверлилки И-38 нормальный ПКР равен 60%. Для ряда других сверлилок ПКР равняется 40%. Эта характеристика указывается в паспорте инструмента.
[c.78]
Электроинструменты рассчитаны на повторно-кратковременный режим работы. Во избежание выхода инструмента из строя не следует допускать перегрева его двигателя. Обмотки электродвигателя не должны нагреваться выше 95°. [c.213]
Включив сверлилку, необходимо помнить, что она рассчитана на повторно-кратковременный режим работы или длите ль-ный с переменной нагрузкой, и поэтому необходимо следить за нагревом, прекращая работу, если нельзя длительно держать ладонь на ее корпусе. [c.224]
[c.224]
Повторно-кратковременным называется такой режим, при котором работа электродвигателя в течение одного цикла чередуется с остановками продолжительностью не более 10 мин. Повторно-кратковременный режим характеризуется отношением продолжительности рабочего периода к продолжительности всего рабочего цикла. [c.46]
Номинальная мощность, ква. . Первичное напряжение, в. .. Повторно-кратковременный режим
[c.58]
Повторно-кратковременный режим работы при цикле 10 мин с отключением первичной обмотки силового трансформатора выпрямителя от сети во время паузы. [c.132]
ОТНОСИТЕЛЬНАЯ ПРОДОЛЖИТЕЛЬНОСТЬ РАБОТЫ — см. Повторно-кратковременный режим. [c.98]
ПОВТОРНО-КРАТКОВРЕМЕННЫЙ РЕЖИМ (для электрической машины или аппарата) — номинальный режим, при котором кратковременные рабочие периоды чередуются с паузами, т. е. с кратковременными периодами остановки без работы или без нагрузки, причем температура частей машины или аппарата в период включения не достигает установившегося значения, а в период паузы не успевает достигнуть
[c. 106]
106]
Определение мощности электродвигателя при повторно-кратковременной нагрузке. Повторно-кратковременный режим работы электродвигателя характеризуется короткими периодами нагрузки, за которые температура электродвигателя не успевает достичь установившегося значения, а за время кратковременных отключений электродвигателя от сети он не успевает охладиться до температуры окружающей среды. При таком режиме перегрев электродвигателя изменяется по пилообразной ломаной линии, состоящей из чередующихся отрезков кривых нагревания и охлаждения. Этот режим наиболее характерен для приводов большинства металлорежущих станков. Время одного цикла не должно превышать 10 мин. Мощность электродвигателя, работающего в повторно-кратковременном режиме, наиболее удобно определять по методу средних потерь. [c.78]
Другой характеристикой источника питания является п р одолжи-тельность рабо-т ы (ПР) или продолжительность включения (ПВ). Обе эти величины определяют повторно-кратковременный режим работы, на который рассчитан данный источник питания, и представляет собой отношение времени сварки ко времени всего цикла, т. е. к общему времени сварки и паузы величины ПР и ПВ выражают в процентах.
[c.227]
е. к общему времени сварки и паузы величины ПР и ПВ выражают в процентах.
[c.227]
Повторно — кратковременный режим, при котором рабочие периоды чередуются с периодами отключения силовой цепи источника от сети. Режим характеризуется относительной продолжительностью включения ПВ = Гр / Гц, или в процентах ПВ = (Гр / Гц) 100 %. [c.220]
Эффективность работы жидкостного нейтрализатора зависит от режимов работы двигателя. Постоянная работа на режимах, близких к номинальным, приводит к перегреву неДтрализуюшего раствора, резко снижая его поглотительную способность и одновременно увеличивая его расход. Наиболее оптимален для работы жидкостного нейтрализатора повторно-кратковременный режим работы двигателя с умеренными средними температурами и расходами ОГ. Такие режимы имеют место, например,при работе автосамосвалов в условиях горных выработок. [c.79]
Повторно-кратковременный режим харлктеризуется относительной продолжительностью включения, которая определяется отношением времени работы электродвигателя к полному времени цикла (ПВ, %). [c.17]
[c.17]
Г1КР означает повторно-кратковременный режим работы, выражающийся как отношение времени горения дуги ко времени работы генератора при 5-минутном цикле, в процентах. [c.278]
Работа электрооборудования грузоподъемных кранов отличается рядом специфических особенностей. К ним относятся повторно-кратковременный режим работы, частые изменения направления вращения (реверс), необходимость регулирования частоты вращения привода, значительные перегрузки, вибрация, затрудненный доступ для обслу-жив1ания й ремонта, а для башенных кранов также работа в условиях загрязненности, влажности, значительного перепада температур. Поэтому элёктрооборудование должно обладать повышенной прочностью, высококачественной изоляцией и надежной защитой от окружающей среды. Этому требованию отвечают машины и аппараты специального кранового исполнения. Однако на некоторых кранах применяют также электрические машины и аппараты общего (не кранового) исполнения. [c.337]
[c.337]
Работа электрооборудования грузоподъемных кранов отличается рядом специфических особенностей. К ним относятся повторно-кратковременный режим работы, частые изменения направления вращения (реверс), необходимость регулирования скорости привода, значительные перегрузки, вибрация, затрудненный доступ для обслуживания и ремонта, а для башенных кранов также работа в условиях загрязненности, влажности, значительного перепада температур. Поэтому электрооборудование должно обладать повышенной прочностью, высо- [c.99]
Катушка нормального исполнения допускает продолжительное вклю-ченне (ПВ 100%), усиленная (с увеличенным тяговым усилием) — только повторни-кратковременный режим работы при ПВ не более 40%. [c.168]
Сирена рассчитана на повторно-кратковременный режим работы. Продолжительность непрерывного сигнала более 8 мин по температурному режиму недопустима. Обмоточные данные напряжение ПО В, число витков — 400, марка провода ПЭЛ0Д5, сопротивление 240 5% Ом.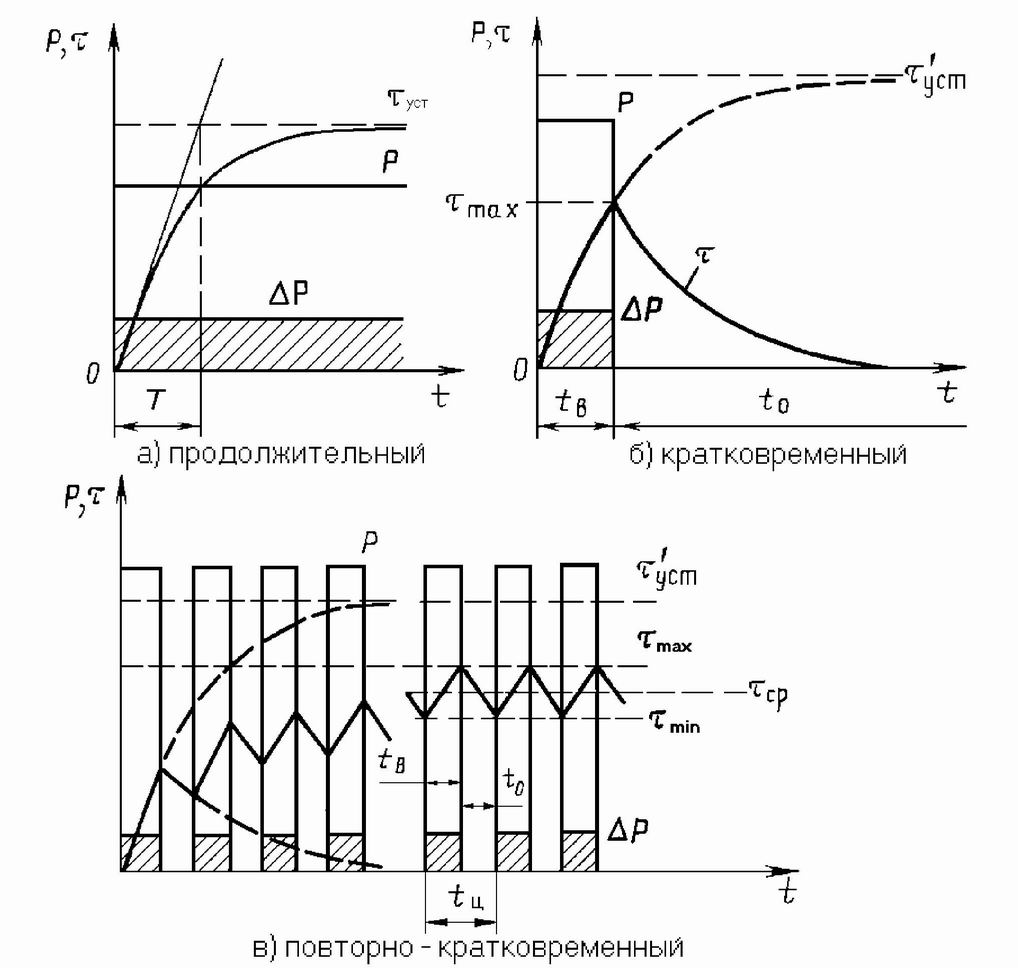 [c.205]
[c.205]
Режимы электродвигателей
Режимы работы асинхронного электродвигателя
При выборе электродвигателя нужно учитывать довольно много параметров, таких как: номинальная мощность, число оборотов в минуту, способ монтажного крепления, габаритные размеры, климатическое исполнение, степень защиты и так далее.
Немаловажным параметром при выборе эл двигателя является номинальный режим работы электродвигателя. В этой статье мы рассмотрим режимы работы электродвигателей и объясним, почему так важно учитывать этот фактор.
• S1 – Продолжительный
В режиме работы электродвигателя S1 агрегат продолжительное время работает от сети напряжения с неизменной нагрузкой. Постепенно разогревается до рабочей температуры, а параметры работы остаются неизменными. Большинство общепромышленных электродвигателей имеют именно такой режим работы. Характеризуется определенной относительной продолжительностью включений ПВ -100%.
• S2 – Кратковременный
При включении в сеть агрегат достигает постоянной нагрузки в течение десяти, тридцати, 60 или 90 минут. Времени недостаточно для достижения максимального нагрева, а когда не работает, электромотор остывает до внешних температур. Применяется в устройствах подающих рабочие вещества, например нефть, газ или воду. Применяется, к примеру, в запорных устройствах.
• S3 – Повторно-кратковременный
Электродвигатель так же не достигает максимального нагрева, но в отличие от S2 при остановке не остывает до внешней температуры. Применяется для привода в башенных кранах, в оборудовании для работы лифтов и эскалаторов. Агрегаты работают при ПВ 15, 25, 40, 60%.
• S4 – режим S3 с частыми пусками
Продолжительность работы и число пусков примерно одинаковы, включений в час: 30, 60, 120 и 240. Режим циклический, пуск-работа-остановка и так далее по кругу.
• S5 – Режим повторно –кратковременный с добавлением электрического торможения
В конце каждого цикла агрегат принудительно останавливается.
• S6 – Периодический режим (нагрузка на электродвигатель кратковременная)
Повторяющиеся циклы работы, однако, агрегат не успевает нагреться до постоянной температуры, но и не успевает остыть. Чередование холостого хода с нагрузкой на агрегат.
• S7 – Периодический (нагрузка кратковременна и торможение в конце)
Аналогично S6, но добавлено торможение электродвигателя в конце каждого цикла.
• S8 – Периодический режим (нагрузка кратковременна и изменяется с изменениями скорости вращения механизма)
Происходят взаимосвязанные циклические изменения нагрузки электродвигателя и скорости вращения его вала.
• S9 – Особый (периодические изменения нагрузки агрегата и частоты вращения)
Изменения происходят произвольным образом. Допускается работа с перегрузкой.
Допускается работа с перегрузкой.
Как Вы видите, каждый режим работы предназначен для достижения определенных целей. Использовать электродвигатель с режимом несоответствующим условиям эксплуатации крайне не рекомендуется. Агрегат проработает непродолжительное время или вообще прекратит свою работу сразу после первого цикла включения. Конструктивная особенность эл двигателя каждого режима работы индивидуальна. Если Вы сомневаетесь и нужна консультация, обратитесь к нашим специалистам.
Электродвигатель АИР характеристики
| Тип двигателя | Р, кВт | Номинальная частота вращения, об/мин | кпд,* | COS ф | 1п/1н | Мп/Мн | Мmах/Мн | 1н, А | Масса, кг |
| АИР56А2 | 0,18 | 2840 | 68,0 | 0,78 | 5,0 | 2,2 | 2,2 | 0,52 | 3,4 |
| АИР56В2 | 0,25 | 2840 | 68,0 | 0,698 | 5,0 | 2,2 | 2,2 | 0,52 | 3,9 |
| АИР56А4 | 0,12 | 1390 | 63,0 | 0,66 | 5,0 | 2,1 | 2,2 | 0,44 | 3,4 |
| АИР56В4 | 0,18 | 1390 | 64,0 | 0,68 | 5,0 | 2,1 | 2,2 | 0,65 | 3,9 |
| АИР63А2 | 0,37 | 2840 | 72,0 | 0,86 | 5,0 | 2,2 | 2,2 | 0,91 | 4,7 |
| АИР63В2 | 0,55 | 2840 | 75,0 | 0,85 | 5,0 | 2,2 | 2,3 | 1,31 | 5,5 |
| АИР63А4 | 0,25 | 1390 | 68,0 | 0,67 | 5,0 | 2,1 | 2,2 | 0,83 | 4,7 |
| АИР63В4 | 0,37 | 1390 | 68,0 | 0,7 | 5,0 | 2,1 | 2,2 | 1,18 | 5,6 |
| АИР63А6 | 0,18 | 880 | 56,0 | 0,62 | 4,0 | 1,9 | 2 | 0,79 | 4,6 |
| АИР63В6 | 0,25 | 880 | 59,0 | 0,62 | 4,0 | 1,9 | 2 | 1,04 | 5,4 |
| АИР71А2 | 0,75 | 2840 | 75,0 | 0,83 | 6,1 | 2,2 | 2,3 | 1,77 | 8,7 |
| АИР71В2 | 1,1 | 2840 | 76,2 | 0,84 | 6,9 | 2,2 | 2,3 | 2,6 | 10,5 |
| АИР71А4 | 0,55 | 1390 | 71,0 | 0,75 | 5,2 | 2,4 | 2,3 | 1,57 | 8,4 |
| АИР71В4 | 0,75 | 1390 | 73,0 | 0,76 | 6,0 | 2,3 | 2,3 | 2,05 | 10 |
| АИР71А6 | 0,37 | 880 | 62,0 | 0,70 | 4,7 | 1,9 | 2,0 | 1,3 | 8,4 |
| АИР71В6 | 0,55 | 880 | 65,0 | 0,72 | 4,7 | 1,9 | 2,1 | 1,8 | 10 |
| АИР71А8 | 0,25 | 645 | 54,0 | 0,61 | 4,7 | 1,8 | 1,9 | 1,1 | 9 |
| АИР71В8 | 0,25 | 645 | 54,0 | 0,61 | 4,7 | 1,8 | 1,9 | 1,1 | 9 |
| АИР80А2 | 1,5 | 2850 | 78,5 | 0,84 | 7,0 | 2,2 | 2,3 | 3,46 | 13 |
| АИР80А2ЖУ2 | 1,5 | 2850 | 78,5 | 0,84 | 7,0 | 2,2 | 2,3 | 3,46 | 13 |
| АИР80В2 | 2,2 | 2855 | 81,0 | 0,85 | 7,0 | 2,2 | 2,3 | 4,85 | 15 |
| АИР80В2ЖУ2 | 2,2 | 2855 | 81,0 | 0,85 | 7,0 | 2,2 | 2,3 | 4,85 | 15 |
| АИР80А4 | 1,1 | 1390 | 76,2 | 0,77 | 6,0 | 2,3 | 2,3 | 2,85 | 14 |
| АИР80В4 | 1,5 | 1400 | 78,5 | 0,78 | 6,0 | 2,3 | 2,3 | 3,72 | 16 |
| АИР80А6 | 0,75 | 905 | 69,0 | 0,72 | 5,3 | 2,0 | 2,1 | 2,3 | 14 |
| АИР80В6 | 1,1 | 905 | 72,0 | 0,73 | 5,5 | 2,0 | 2,1 | 3,2 | 16 |
| АИР80А8 | 0,37 | 675 | 62,0 | 0,61 | 4,0 | 1,8 | 1,9 | 1,49 | 15 |
| АИР80В8 | 0,55 | 680 | 63,0 | 0,61 | 4,0 | 1,8 | 2,0 | 2,17 | 18 |
| АИР90L2 | 3,0 | 2860 | 82,6 | 0,87 | 7,5 | 2,2 | 2,3 | 6,34 | 17 |
| АИР90L2ЖУ2 | 3,0 | 2860 | 82,6 | 0,87 | 7,5 | 2,2 | 2,3 | 6,34 | 17 |
| АИР90L4 | 2,2 | 1410 | 80,0 | 0,81 | 7,0 | 2,3 | 2,3 | 5,1 | 17 |
| АИР90L6 | 1,5 | 920 | 76,0 | 0,75 | 5,5 | 2,0 | 2,1 | 4,0 | 18 |
| АИР90LA8 | 0,75 | 680 | 70,0 | 0,67 | 4,0 | 1,8 | 2,0 | 2,43 | 23 |
| АИР90LB8 | 1,1 | 680 | 72,0 | 0,69 | 5,0 | 1,8 | 2,0 | 3,36 | 28 |
| АИР100S2 | 4,0 | 2880 | 84,2 | 0,88 | 7,5 | 2,2 | 2,3 | 8,2 | 20,5 |
| АИР100S2ЖУ2 | 4,0 | 2880 | 84,2 | 0,88 | 7,5 | 2,2 | 2,3 | 8,2 | 20,5 |
| АИР100L2 | 5,5 | 2900 | 85,7 | 0,88 | 7,5 | 2,2 | 2,3 | 11,1 | 28 |
| АИР100L2ЖУ2 | 5,5 | 2900 | 85,7 | 0,88 | 7,5 | 2,2 | 2,3 | 11,1 | 28 |
| АИР100S4 | 3,0 | 1410 | 82,6 | 0,82 | 7,0 | 2,3 | 2,3 | 6,8 | 21 |
| АИР100L4 | 4,0 | 1435 | 84,2 | 0,82 | 7,0 | 2,3 | 2,3 | 8,8 | 37 |
| АИР100L6 | 2,2 | 935 | 79,0 | 0,76 | 6,5 | 2,0 | 2,1 | 5,6 | 33,5 |
| АИР100L8 | 1,5 | 690 | 74,0 | 0,70 | 5,0 | 1,8 | 2,0 | 4,4 | 33,5 |
| АИР112M2 | 7,5 | 2895 | 87,0 | 0,88 | 7,5 | 2,2 | 2,3 | 14,9 | 49 |
| АИР112М2ЖУ2 | 7,5 | 2895 | 87,0 | 0,88 | 7,5 | 2,2 | 2,3 | 14,9 | 49 |
| АИР112М4 | 5,5 | 1440 | 85,7 | 0,83 | 7,0 | 2,3 | 2,3 | 11,7 | 45 |
| АИР112MA6 | 3,0 | 960 | 81,0 | 0,73 | 6,5 | 2,1 | 2,1 | 7,4 | 41 |
| АИР112MB6 | 4,0 | 860 | 82,0 | 0,76 | 6,5 | 2,1 | 2,1 | 9,75 | 50 |
| АИР112MA8 | 2,2 | 710 | 79,0 | 0,71 | 6,0 | 1,8 | 2,0 | 6,0 | 46 |
| АИР112MB8 | 3,0 | 710 | 80,0 | 0,73 | 6,0 | 1,8 | 2,0 | 7,8 | 53 |
| АИР132M2 | 11 | 2900 | 88,4 | 0,89 | 7,5 | 2,2 | 2,3 | 21,2 | 54 |
| АИР132М2ЖУ2 | 11 | 2900 | 88,4 | 0,89 | 7,5 | 2,2 | 2,3 | 21,2 | 54 |
| АИР132S4 | 7,5 | 1460 | 87,0 | 0,84 | 7,0 | 2,3 | 2,3 | 15,6 | 52 |
| АИР132M4 | 11 | 1450 | 88,4 | 0,84 | 7,0 | 2,2 | 2,3 | 22,5 | 60 |
| АИР132S6 | 5,5 | 960 | 84,0 | 0,77 | 6,5 | 2,1 | 2,1 | 12,9 | 56 |
| АИР132M6 | 7,5 | 970 | 86,0 | 0,77 | 6,5 | 2,0 | 2,1 | 17,2 | 61 |
| АИР132S8 | 4,0 | 720 | 81,0 | 0,73 | 6,0 | 1,9 | 2,0 | 10,3 | 70 |
| АИР132M8 | 5,5 | 720 | 83,0 | 0,74 | 6,0 | 1,9 | 2,0 | 13,6 | 86 |
| АИР160S2 | 15 | 2930 | 89,4 | 0,89 | 7,5 | 2,2 | 2,3 | 28,6 | 116 |
| АИР160S2ЖУ2 | 15 | 2930 | 89,4 | 0,89 | 7,5 | 2,2 | 2,3 | 28,6 | 116 |
| АИР160M2 | 18,5 | 2930 | 90,0 | 0,90 | 7,5 | 2,0 | 2,3 | 34,7 | 130 |
| АИР160М2ЖУ2 | 18,5 | 2930 | 90,0 | 0,90 | 7,5 | 2,0 | 2,3 | 34,7 | 130 |
| АИР160S4 | 15 | 1460 | 89,4 | 0,85 | 7,5 | 2,2 | 2,3 | 30,0 | 125 |
| АИР160S4ЖУ2 | 15 | 1460 | 89,4 | 0,85 | 7,5 | 2,2 | 2,3 | 30,0 | 125 |
| АИР160M4 | 18,5 | 1470 | 90,0 | 0,86 | 7,5 | 2,2 | 2,3 | 36,3 | 142 |
| АИР160S6 | 11 | 970 | 87,5 | 0,78 | 6,5 | 2,0 | 2,1 | 24,5 | 125 |
| АИР160M6 | 15 | 970 | 89,0 | 0,81 | 7,0 | 2,0 | 2,1 | 31,6 | 155 |
| АИР160S8 | 7,5 | 720 | 85,5 | 0,75 | 6,0 | 1,9 | 2,0 | 17,8 | 125 |
| АИР160M8 | 11 | 730 | 87,5 | 0,75 | 6,5 | 2,0 | 2,0 | 25,5 | 150 |
| АИР180S2 | 22 | 2940 | 90,5 | 0,90 | 7,5 | 2,0 | 2,3 | 41,0 | 150 |
| АИР180S2ЖУ2 | 22 | 2940 | 90,5 | 0,90 | 7,5 | 2,0 | 2,3 | 41,0 | 150 |
| АИР180M2 | 30 | 2950 | 91,4 | 0,90 | 7,5 | 2,0 | 2,3 | 55,4 | 170 |
| АИР180М2ЖУ2 | 30 | 2950 | 91,4 | 0,90 | 7,5 | 2,0 | 2,3 | 55,4 | 170 |
| АИР180S4 | 22 | 1470 | 90,5 | 0,86 | 7,5 | 2,2 | 2,3 | 43,2 | 160 |
| АИР180S4ЖУ2 | 22 | 1470 | 90,5 | 0,86 | 7,5 | 2,2 | 2,3 | 43,2 | 160 |
| АИР180M4 | 30 | 1470 | 91,4 | 0,86 | 7,2 | 2,2 | 2,3 | 57,6 | 190 |
| АИР180М4ЖУ2 | 30 | 1470 | 91,4 | 0,86 | 7,2 | 2,2 | 2,3 | 57,6 | 190 |
| АИР180M6 | 18,5 | 980 | 90,0 | 0,81 | 7,0 | 2,1 | 2,1 | 38,6 | 160 |
| АИР180M8 | 15 | 730 | 88,0 | 0,76 | 6,6 | 2,0 | 2,0 | 34,1 | 172 |
| АИР200M2 | 37 | 2950 | 92,0 | 0,88 | 7,5 | 2,0 | 2,3 | 67,9 | 230 |
| АИР200М2ЖУ2 | 37 | 2950 | 92,0 | 0,88 | 7,5 | 2,0 | 2,3 | 67,9 | 230 |
| АИР200L2 | 45 | 2960 | 92,5 | 0,90 | 7,5 | 2,0 | 2,3 | 82,1 | 255 |
| АИР200L2ЖУ2 | 45 | 2960 | 92,5 | 0,90 | 7,5 | 2,0 | 2,3 | 82,1 | 255 |
| АИР200M4 | 37 | 1475 | 92,0 | 0,87 | 7,2 | 2,2 | 2,3 | 70,2 | 230 |
| АИР200L4 | 45 | 1475 | 92,5 | 0,87 | 7,2 | 2,2 | 2,3 | 84,9 | 260 |
| АИР200M6 | 22 | 980 | 90,0 | 0,83 | 7,0 | 2,0 | 2,1 | 44,7 | 195 |
| АИР200L6 | 30 | 980 | 91,5 | 0,84 | 7,0 | 2,0 | 2,1 | 59,3 | 225 |
| АИР200M8 | 18,5 | 730 | 90,0 | 0,76 | 6,6 | 1,9 | 2,0 | 41,1 | 210 |
| АИР200L8 | 22 | 730 | 90,5 | 0,78 | 6,6 | 1,9 | 2,0 | 48,9 | 225 |
| АИР225M2 | 55 | 2970 | 93,0 | 0,90 | 7,5 | 2,0 | 2,3 | 100 | 320 |
| АИР225M4 | 55 | 1480 | 93,0 | 0,87 | 7,2 | 2,2 | 2,3 | 103 | 325 |
| АИР225M6 | 37 | 980 | 92,0 | 0,86 | 7,0 | 2,1 | 2,1 | 71,0 | 360 |
| АИР225M8 | 30 | 735 | 91,0 | 0,79 | 6,5 | 1,9 | 2,0 | 63 | 360 |
| АИР250S2 | 75 | 2975 | 93,6 | 0,90 | 7,0 | 2,0 | 2,3 | 135 | 450 |
| АИР250M2 | 90 | 2975 | 93,9 | 0,91 | 7,1 | 2,0 | 2,3 | 160 | 530 |
| АИР250S4 | 75 | 1480 | 93,6 | 0,88 | 6,8 | 2,2 | 2,3 | 138,3 | 450 |
| АИР250M4 | 90 | 1480 | 93,9 | 0,88 | 6,8 | 2,2 | 2,3 | 165,5 | 495 |
| АИР250S6 | 45 | 980 | 92,5 | 0,86 | 7,0 | 2,1 | 2,0 | 86,0 | 465 |
| АИР250M6 | 55 | 980 | 92,8 | 0,86 | 7,0 | 2,1 | 2,0 | 104 | 520 |
| АИР250S8 | 37 | 740 | 91,5 | 0,79 | 6,6 | 1,9 | 2,0 | 78 | 465 |
| АИР250M8 | 45 | 740 | 92,0 | 0,79 | 6,6 | 1,9 | 2,0 | 94 | 520 |
| АИР280S2 | 110 | 2975 | 94,0 | 0,91 | 7,1 | 1,8 | 2,2 | 195 | 650 |
| АИР280M2 | 132 | 2975 | 94,5 | 0,91 | 7,1 | 1,8 | 2,2 | 233 | 700 |
| АИР280S4 | 110 | 1480 | 94,5 | 0,88 | 6,9 | 2,1 | 2,2 | 201 | 650 |
| АИР280M4 | 132 | 1480 | 94,8 | 0,88 | 6,9 | 2,1 | 2,2 | 240 | 700 |
| АИР280S6 | 75 | 985 | 93,5 | 0,86 | 6,7 | 2,0 | 2,0 | 142 | 690 |
| АИР280M6 | 90 | 985 | 93,8 | 0,86 | 6,7 | 2,0 | 2,0 | 169 | 800 |
| АИР280S8 | 55 | 740 | 92,8 | 0,81 | 6,6 | 1,8 | 2,0 | 111 | 690 |
| АИР280M8 | 75 | 740 | 93,5 | 0,81 | 6,2 | 1,8 | 2,0 | 150 | 800 |
| АИР315S2 | 160 | 2975 | 94,6 | 0,92 | 7,1 | 1,8 | 2,2 | 279 | 1170 |
| АИР315M2 | 200 | 2975 | 94,8 | 0,92 | 7,1 | 1,8 | 2,2 | 248 | 1460 |
| АИР315МВ2 | 250 | 2975 | 94,8 | 0,92 | 7,1 | 1,8 | 2,2 | 248 | 1460 |
| АИР315S4 | 160 | 1480 | 94,9 | 0,89 | 6,9 | 2,1 | 2,2 | 288 | 1000 |
| АИР315M4 | 200 | 1480 | 94,9 | 0,89 | 6,9 | 2,1 | 2,2 | 360 | 1200 |
| АИР315S6 | 110 | 985 | 94,0 | 0,86 | 6,7 | 2,0 | 2,0 | 207 | 880 |
| АИР315М(А)6 | 132 | 985 | 94,2 | 0,87 | 6,7 | 2,0 | 2,0 | 245 | 1050 |
| АИР315MВ6 | 160 | 985 | 94,2 | 0,87 | 6,7 | 2,0 | 2,0 | 300 | 1200 |
| АИР315S8 | 90 | 740 | 93,8 | 0,82 | 6,4 | 1,8 | 2,0 | 178 | 880 |
| АИР315М(А)8 | 110 | 740 | 94,0 | 0,82 | 6,4 | 1,8 | 2,0 | 217 | 1050 |
| АИР315MВ8 | 132 | 740 | 94,0 | 0,82 | 6,4 | 1,8 | 2,0 | 260 | 1200 |
| АИР355S2 | 250 | 2980 | 95,5 | 0,92 | 6,5 | 1. 6 6 | 2,3 | 432,3 | 1700 |
| АИР355M2 | 315 | 2980 | 95,6 | 0,92 | 7,1 | 1,6 | 2,2 | 544 | 1790 |
| АИР355S4 | 250 | 1490 | 95,6 | 0,90 | 6,2 | 1,9 | 2,9 | 441 | 1700 |
| АИР355M4 | 315 | 1480 | 95,6 | 0,90 | 6,9 | 2,1 | 2,2 | 556 | 1860 |
| АИР355MА6 | 200 | 990 | 94,5 | 0,88 | 6,7 | 1,9 | 2,0 | 292 | 1550 |
| АИР355S6 | 160 | 990 | 95,1 | 0,88 | 6,3 | 1,6 | 2,8 | 291 | 1550 |
| АИР355МВ6 | 250 | 990 | 94,9 | 0,88 | 6,7 | 1,9 | 2,0 | 454,8 | 1934 |
| АИР355L6 | 315 | 990 | 94,5 | 0,88 | 6,7 | 1,9 | 2,0 | 457 | 1700 |
| АИР355S8 | 132 | 740 | 94,3 | 0,82 | 6,4 | 1,9 | 2,7 | 259,4 | 1800 |
| АИР355MА8 | 160 | 740 | 93,7 | 0,82 | 6,4 | 1,8 | 2,0 | 261 | 2000 |
| АИР355MВ8 | 200 | 740 | 94,2 | 0,82 | 6,4 | 1,8 | 2,0 | 315 | 2150 |
| АИР355L8 | 132 | 740 | 94,5 | 0,82 | 6,4 | 1,8 | 2,0 | 387 | 2250 |
Обозначение режимов работы электродвигателя по ГОСТ Р 52776-2007
В станках используют самые разные электродвигатели: они устанавливаются на шпиндель, приводят в движение оси, конвейеры удаления стружки или выгрузки деталей. Эти двигатели работают в разных режимах — продолжительно или кратковременно, под нагрузкой или же вхолостую. В зависимости от параметров работы различают 10 номинальных режимов работы электродвигателя. Они описаны в ГОСТ Р 52776-2007 и обозначаются от S1 до S10.
Эти двигатели работают в разных режимах — продолжительно или кратковременно, под нагрузкой или же вхолостую. В зависимости от параметров работы различают 10 номинальных режимов работы электродвигателя. Они описаны в ГОСТ Р 52776-2007 и обозначаются от S1 до S10.
Разберем каждый из этих режимов подробнее.
Номинальные режимы работы двигателей в станках
S1 – продолжительный режим работы двигателя
Он характеризуется постоянной нагрузкой в течение длительного времени, при этом электродвигатель прогревается до неизменной рабочей температуры. Ниже приведены три графика, которые показывают, как нагружается двигатель, как меняются потери и температура.
Р – нагрузка. На графике видно, что двигатель испытывает постоянную нагрузку с самого начала цикла.
Рэ – потери энергии, которые неизбежно возникают при преобразовании электрической энергии в механическую. В случае с электродвигателем это электрические потери, которые можно рассчитать по формуле:
Рэ=Ii2riv
Где riv – сопротивление обмотки двигателя, а Ii – это сила тока, протекающего по этой обмотке.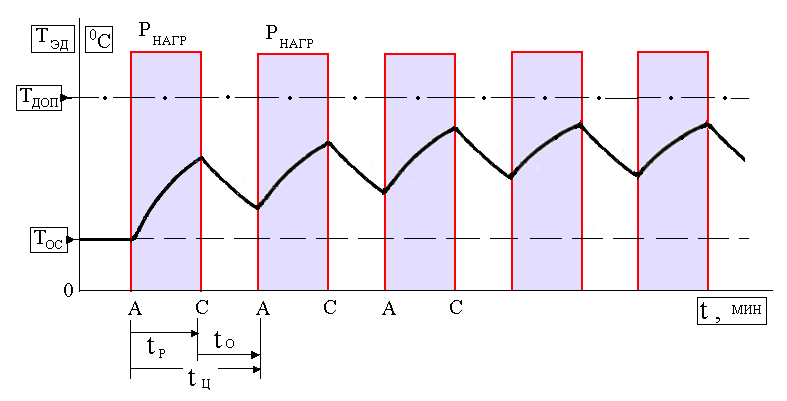 Значения этих переменных получают путём измерений.
Значения этих переменных получают путём измерений.
Потери рассчитывают для каждой обмотки двигателя отдельно, а общие потери двигателя определяют как сумму потерь всех его обмоток. На графике видно, что потери не изменяются при постоянной нагрузке, так как сопротивление на обмотках электродвигателя тоже не изменяется.
Θmax – это максимальная температура электродвигателя. Этот параметр можно получить путём измерений. На графике видно, что после определенного времени работы электродвигателя с постоянной нагрузкой температура в дальнейшем не меняется — охлаждение двигателя компенсирует тепловыделение.
S2 – кратковременный режим работы двигателя
В таком режиме электродвигатель работает определенное время (Δtp), после чего его останавливают, чтобы он остыл до температуры окружающей среды.
Как и в предыдущем режиме, нагрузка и потери остаются неизменными на всем протяжении времени Δtp. За это время двигатель нагревается до температуры Θmax, после чего останавливается. Величина Δtp нормируется — она может быть равна 15, 30, 60 и 90 минутам. При указании режима работы S2 необходимо указывать Δtp, например, S2 60 мин.
За это время двигатель нагревается до температуры Θmax, после чего останавливается. Величина Δtp нормируется — она может быть равна 15, 30, 60 и 90 минутам. При указании режима работы S2 необходимо указывать Δtp, например, S2 60 мин.
S3 – повторно кратковременный режим работы
Этот режим работы характеризуется повторяющимися рабочими циклами с постоянной нагрузкой, прерывающимися на определенное время (Δtотк), за которое двигатель не успевает охладиться до температуры окружающей среды.
Величина Т – это цикл включения от пуска до пуска, который также включает время остановки двигателя.
Этот режим характеризуется продолжительностью включения в процентном соотношении и рассчитывается по формуле
ПВ=∆tp(∆tp+∆tотк)×100%
Продолжительность включения также является нормируемой величиной и может принимать значения 15, 25, 40 и 60%.
S6 – перемежающийся режим работы электродвигателя
Этот режим работы характеризуется попеременной работой на вхолостую и под нагрузкой. В перемежающемся режиме двигатель не останавливается.
В перемежающемся режиме двигатель не останавливается.
На графике отмечено время работы с постоянной нагрузкой Δtp, время работы в холостом режиме Δtо и время цикла Т.
Также можно заметить, что электрические потери при отсутствии нагрузки уменьшаются, что связано с уменьшением сопротивления катушек электродвигателя.
Так же как и S3, перемежающийся режим характеризуется продолжительностью включения в процентном соотношении, которое рассчитывается по формуле:
ПВ=∆tp(∆tp+∆tо)×100%
Как было сказано выше, этот параметр нормированный и всегда указывается с продолжительностью включения, например S6 60%.
Перечисленные режимы являются основными, которые используются для двигателей станков. Существуют также другие режимы.
Другие номинальные режимы работы электродвигателей
S4 – повторно-кратковременный режим работы электродвигателя с влиянием пусковых процессов. В таком режиме пуск двигателя происходит достаточно продолжительное время и влияет на изменение его температуры.
В таком режиме пуск двигателя происходит достаточно продолжительное время и влияет на изменение его температуры.
S5 – повторно-кратковременный режим работы электродвигателя с влиянием пусковых процессов и торможением. Этот режим включает в себя достаточно длительное время пуска, которое влияет на его температуру, время работы с постоянной нагрузкой, время торможения и время полной остановки двигателя.
S7 – перемежающийся режим работы двигателя с влиянием пусковых процессов и торможения. Данный режим работы двигателя характерен отсутствием остановки двигателя, режимом работы с постоянной нагрузкой, режимом работы без нагрузки, длительным процессом пуска, влияющим на его температуру, а так же торможением.
S8 – перемежающийся режим работы двигателя с периодически изменяющейся частотой вращения. Так же, как и предыдущий режим не содержит остановок двигателя, но содержит цикл работы с постоянной нагрузкой на заданной частоте вращения, за которым следует один или более циклов при других постоянных нагрузках и другой частоте вращения.
S9 – режим работы электродвигателя с непериодическими изменениями нагрузки и частоты вращения. В отличие от режима S8, в данном режиме смена нагрузки и частоты вращения происходит непериодически. Для такого режима работы характерны перегрузки.
S10 – режим работы электродвигателя с дискретными постоянными нагрузками и частотами вращения. Этот режим работы с непостоянными нагрузками и частотой вращения, которые сохраняются достаточно длительное время, чтобы температура достигла неизменной величины. При этом нагрузка может достигать нулевого значения (холостой ход или режим покоя).
Соответствие режима работы двигателя его применению в станке
Режимы работы S1, S2, S3 и S6 подходят для двигателей шпинделя, поэтому в характеристиках станка часто указывают параметр мощности и момента для этих режимов.
В режиме S1 двигатель может длительное время работать с допустимой нагрузкой, его температура будет стабильна и перегрева двигателя не произойдет.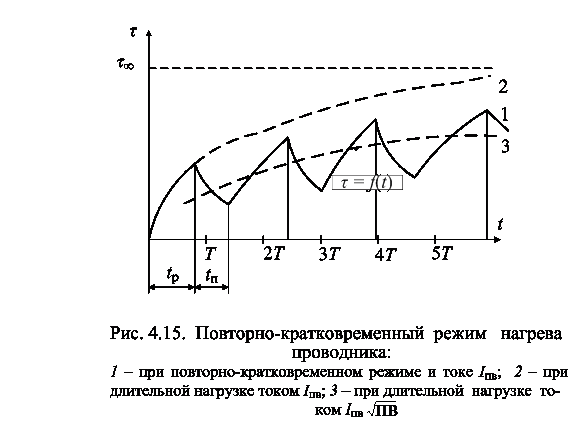 Современные фрезерные станки с ЧПУ часто используют для обработки сложных фасонных поверхностей. Такая обработка может длиться очень долго, поэтому двигатель шпинделя должен быть приспособлен для работы в этом режиме.
Современные фрезерные станки с ЧПУ часто используют для обработки сложных фасонных поверхностей. Такая обработка может длиться очень долго, поэтому двигатель шпинделя должен быть приспособлен для работы в этом режиме.
При операциях сверления листового материала шпиндель станка работает в режиме S2 с постоянными остановка, так как после завершения операции требуется замена готовой детали на заготовку. За это время температура двигателя шпинделя успевает снизиться до температуры окружающей среды. Таким образом, двигатель шпинделя сверлильного станка должен иметь номинальный режим работы S2.
Во время выполнения операции торцевого фрезерования электродвигатель шпинделя работает в режиме S3, так как при движении инструмента в материале двигатель будет работать с нагрузкой, а во время перемещений инструмента в точку резания двигатель работает в холостом режиме, но при этом не успевает остыть до температуры окружающей среды.
Бывают ситуации, при которых толщина удаляемого материала неравномерна. Это приводит к дискретным нагрузкам на двигатель шпинделя. Использование функции постоянной скорости резания на токарных станках приводит к изменению частоты вращения в зависимости от диаметра обработки, что также приводит к неравномерной нагрузке и изменению частоты вращения. Эти ситуации полностью соответствуют режиму S6.
Это приводит к дискретным нагрузкам на двигатель шпинделя. Использование функции постоянной скорости резания на токарных станках приводит к изменению частоты вращения в зависимости от диаметра обработки, что также приводит к неравномерной нагрузке и изменению частоты вращения. Эти ситуации полностью соответствуют режиму S6.
Указанные режимы работы двигателя должны быть предусмотрены производителями станков. Неправильно подобранные электродвигатели не позволят использовать станок максимально эффективно.
Если у вас появились вопросы, специалисты Центра инженерно-технологической поддержки портала СТАНКОТЕКА на них ответят.
Выбор кабеля при повторно-кратковременном режиме работы
В данной статье будет рассматриваться выбор кабеля (провода) по нагреву при повторно-кратковременном режиме работы.
Согласно ПУЭ п.1.3.3, 1.3.4:
Как мы видим ПУЭ допускает увеличении нагрузки для проводников линии с повторно-кратковременным режимом работы, с учетом поправочного коэффициента Кп, определяемый по формуле:
где:
- ПВ = tр/tц – продолжительность включенного состояния (рабочего периода) в относительных единицах;
- tр – время, на которое включается установка (рабочий период) не более 4 мин;
- tп – длительность перерыва в работе;
- tц = tр + tп – общая длительность цикла до 10 мин.


Если условия работы электропроводки требуют применения поправочных коэффициентов учитывающие изменения условий прокладки (по температуре окружающей среды, по числу одновременно проложенных проводников), то тогда общий поправочный коэффициент определяется путем перемножения всех коэффициентов.
Пример 1
Требуется выбрать сечение кабеля марки АВВГнг для электродвигателя станка с повторно-кратковременным режимом работы. Максимальная мощность, потребляемая электродвигателем от сети Uном. = 380 В, составляет Р = 37 кВт при cosφ = 0,88 и КПД = 0,915.
Кабель прокладывается в цехе, с двумя другими кабелями; расстояние в свету между ними 100 мм, температура воздуха составляет +25°С. Длительность работы двигателя tр = 3 мин и длительность перерыва в работе tп = 6 мин.
Решение
1. Определяем общую длительность цикла работы двигателя:
tц = tр + tп = 3 + 6 = 9 мин. < 10 мин. (условие выполняется)
2. Определяем продолжительность включенного состояния в относительных единицах:
ПВ = tр/tц = 3/9 = 0,33 отн. ед.
ед.
3. Определяем значение поправочного коэффициента Кп по ПУЭ п.1.3.3:
4. Определяем значение поправочного коэффициента по температуре К1, учитывая, что температура воздуха в цехе составляет +25°С. Согласно ПУЭ п.1.3.10 допустимые длительные токи для кабелей с поливинхлоридной изоляцией приняты для температур: жил +65, окружающего воздуха +25 и земли + 15°С. По ПУЭ таблица 1.3.3 принимаем значение поправочного коэффициента по температуре К1 = 1,0.
5. Определяем значение поправочного коэффициента K2 по ПУЭ таблица 1.3.26 учитывающий снижение токовой нагрузки при числе работающих кабелей рядом. В моем случае, кабель прокладывается с двумя другими кабелями, расстояние между кабелями составляет 100 мм. Принимаем K2 = 0,85.
6. Определяем расчетный длительно допустимый ток:
7. Определяем ток нагрузки, приведенный к длительному режиму по формуле 5-1 [Л2, с.97]:
8. По таблице 21 ГОСТ 31996-2012 выбираем кабель марки АВВГнг 3х10+1х6, имеющий допустимый ток 50 А при прокладке на воздухе.
Если же Вам нужно проверить уже существующий кабель (провод) на допустимую нагрузку для повторно-кратковременного режима, с учетом поправочных коэффициентов, можно определить по формуле 4 [Л1, с.15].
где:
- К1 – поправочный коэффициент по температуре;
- К2 – поправочный коэффициент по числу одновременно проложенных проводников;
- Кп – поправочный коэффициент для проводников линии с повторно-кратковременным режимом работы;
- Iд.т – длительно допустимый ток кабеля (провода).
Пример 2
На примере определим допустимую нагрузку для кабеля марки АВВГнг 3х25+1х16, имеющий допустимый ток Iд.т.=87 А (таблица 21 ГОСТ 31996-2012) при прокладке на воздухе. Значения поправочных коэффициентов принимаем из примера 1.
1. Определяем допустимую нагрузку для повторно-кратковременного режима, с учетом поправочных коэффициентов по формуле 4 [Л1, с.15]:
где:
- К1 = 1,0 – поправочный коэффициент по температуре;
- К2 = 0,85 – поправочный коэффициент по числу одновременно проложенных проводников;
- Кп = 1,54 – поправочный коэффициент для проводников линии с повторно-кратковременным режимом работы;
- Iд. т.=87 А – длительно допустимый ток кабеля марки АВВГнг 3х25+1х16.
 т.=87 А – длительно допустимый ток кабеля марки АВВГнг 3х25+1х16.
т.=87 А – длительно допустимый ток кабеля марки АВВГнг 3х25+1х16.Литература:
- Как выбрать сечение проводов и кабелей. Ф.Ф. Карпов. 1973 г.
- Электрические сети энергетических систем. В.А. Боровиков. 1977 г.
Всего наилучшего! До новых встреч на сайте Raschet.info.
Поделиться в социальных сетях
Что такое S1, S2, S* и ПВ у электродвигателей? | ЭТМ для профессионалов
На шильдиках электродвигателей кроме механической мощности, номинальных напряжений и токов указывается и режим работы. В зависимости от производителя и года производства информация может несколько отличаться, но обычно режим указывается как S1, S2, S3 и так далее. Кроме обозначения режима может быть и дополнительная информация, обозначенная буквами ПВ и числом в процентах. Ниже прилагаю подборку табличек разных двигателей, собранную на просторах интернета.
Давайте разберёмся что такое S1, S2, S3 и ПВ у электродвигателей!
Режимы работы
Электродвигатели нашли широкое применение в быту и на производстве.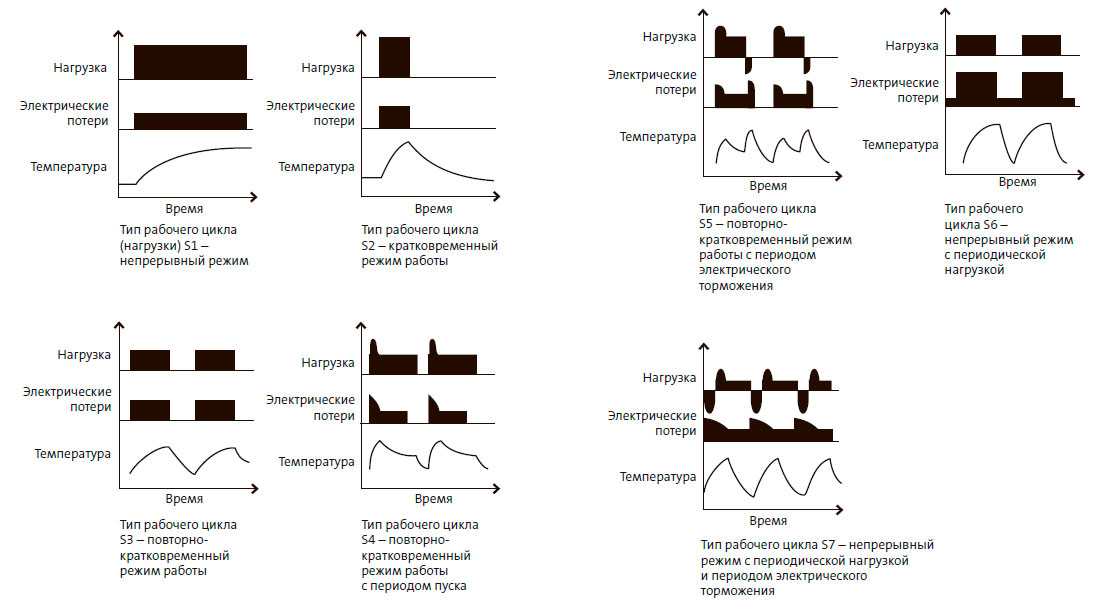 Они используются для привода в движение всевозможных механизмов, например: конвейеров, метало- и деревообрабатывающих станков, задвижек на трубопроводах, компрессоров, лифтов, грузоподъёмных механизмах и прочих.
Они используются для привода в движение всевозможных механизмов, например: конвейеров, метало- и деревообрабатывающих станков, задвижек на трубопроводах, компрессоров, лифтов, грузоподъёмных механизмах и прочих.
Но в каждом применении на двигатель действуют нагрузки разной продолжительности, требуются разные варианты использования, например, длительная работа на протяжении дней, недель и месяцев или, наоборот, кратковременные включения несколько раз в сутки или периодически повторяющиеся включения, остановки и реверс. Сама же нагрузка может быть постоянной, как на конвейере или же изменяться в ходе работы. Особенности работы в каждом конкретном случае описывает режим работы электродвигателя. И каждый двигатель должен быть рассчитан для работы в конкретном режиме.
Режимы работы электродвигателей описаны в ГОСТ IEC 60034-1-2014, определение приведено в п. 3.9, а в разделе 4.2 приведено 10 типовых режимов, каждый из которых обозначается буквой S и цифрой от 1 до 10, например, S1.
Различают 3 основных режима (от S1 до S3), и 7 дополнительных (от S4 до S10), которые более точно описывают особенности работы и изменения нагрузки у двигателя. При рассмотрении режимов будем руководствоваться основной частью информации из ГОСТ IEC 60034-1, некоторые особенности в нём не описаны и за ними обратимся к советскому (ныне не действующему) ГОСТ 183-74 или на справочные данные. В необходимых местах будут приведены ссылки на пункты, из которых взята информация или иллюстрации.
Примечание — ГОСТ различает 10 типовых режимов, и фактического деления на «основные» и «дополнительные» нет, но в разных источниках такое деление присутствует. Я думаю, связано это с тем, что в старом ГОСТе режима было всего 3 и в последующих редакциях они дополнялись.
S1 — продолжительный режим
В режиме S1 — электрические машины работают с постоянной нагрузкой, при этом времени работы достаточно для достижения установившегося теплового состояния.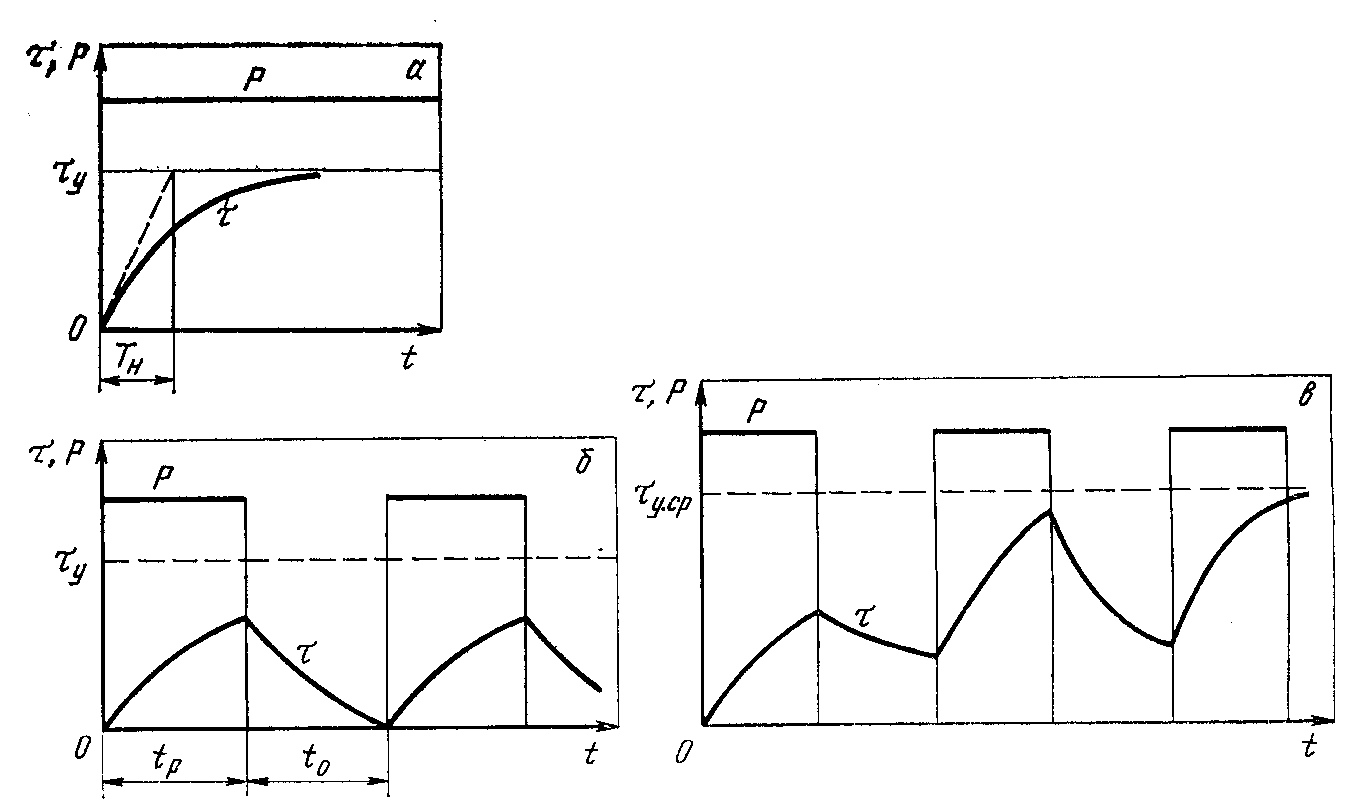 То есть за время своей работы машина нагреется до определённой температуры, и при дальнейшей работе температура изменяться не будет. По времени работы ограничений нет.
То есть за время своей работы машина нагреется до определённой температуры, и при дальнейшей работе температура изменяться не будет. По времени работы ограничений нет.
В таком режиме работают электродвигатели насосов, конвейеров, вентиляторов.
S2 —кратковременный режим
В режиме S2 двигатель работает при постоянной нагрузке в течение определённого времени, за которое не наступает установившееся тепловое состояние. После окончания работы двигатель останавливается на время, достаточное для того, чтобы он остыл до температуры окружающей среды или охлаждающего агента с точностью до 2 К (п. 4.2.2. ГОСТ IEC 60034-1).
Условное обозначение содержит длительность периода нагрузки, например, S2 60 минут. Согласно п. 1.4. ГОСТ 183-74, если не оговорено иное, то периоды нагрузки выбираются из стандартного ряда 10, 30, 60 и 90 минут.
В таком режиме работают электродвигатели приводящие в движение заслонки или другие запорные устройства на трубопроводах.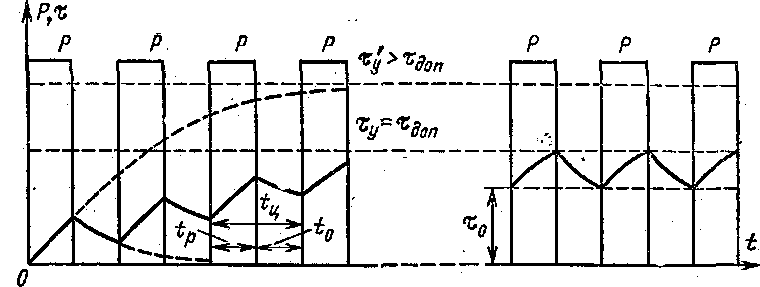
S3 — повторно-кратковременный периодический режим
В режиме S3 двигатель выполняет последовательность повторяющихся одинаковых рабочих циклов. Каждый цикл состоит из периода работы с постоянной нагрузкой и периода покоя (п. 4.2.3. ГОСТ IEC 60034-1), при этом пусковой ток не оказывает существенного влияния на превышение температуры. Цикл работы двигателя в повторно-кратковременном режиме можно записать в виде формулы:
T=Δtр+Δtотк,
где T — время цикла, Δtр — период работы, а Δtотк — период покоя.
Двигатель в периодических режимах за время работы не успевает нагреться до установившейся температуры, а за время паузы не успевает охладиться до температуры окружающей среды.
В условном обозначении режима указывается коэффициент циклической продолжительности включения, например, S3 25%.
В этом режиме работают электродвигатели приводящие в движение грузоподъёмные механизмы, лифты и другие механизмы, работающие циклично.
Что такое продолжительность включения
Продолжительность включения или ПВ — это основная характеристика электродвигателя, работающего в повторно-кратковременном режиме (S3—S8). Характеристика отражает в процентном отношении времени, которое работает двигатель к длительности цикла, и вычисляется по формуле:
ПВ% = (Δtр/T)×100%
Согласно п. 5.2.3. ГОСТ IEC 60034-1 продолжительность включения должна быть равна одному из следующих значений —15, 25, 40, 60%, если не оговорено иное, а длительность цикла не должна превышать 10 минут.
Если режим работы предполагает длительный пуск и/или электрическое торможение, то в расчётах продолжительности включения учитывается время пуска и торможения (согласно п.3.1.1. ГОСТ IEC 60034-1).
Режимы S4 и S5 — повторно-кратковременные
Режимы S4 и S5 по определению похожи на S3, но описывают особенности работы машины подробнее, поэтому предлагаю объединить их одним подзаголовком.
Типовой режим S4 — повторно-кратковременный периодический с пусками. Для этого режима характерны относительно длинные пуски, которые влияют на нагрев машины. Обратите внимание в S4 используется другая формула для определения коэффициента циклической продолжительности включения, учитывающая время пуска (п. 4.2.4. ГОСТ IEC 60034-1).
Цикл работы T состоит из 3 периодов: Δtп — время пуска, Δtр — время работы, Δtотк — время покоя (остановки или отключения питания):
T= Δtп + Δtр + Δtотк
В обозначении режима кроме коэффициента циклической продолжительности указывается момент инерции двигателя Jд и момент инерции нагрузки Jнагр. Пример — S4 25%, Jд=0,15 кг·м², Jнагр= 0,7 кг·м².
Типовой режим S5 — повторно-кратковременный периодический с длительными пусками и электрическим торможением. В этом случае учитывается влияние на температуру машины и пусковых токов, и токов, протекающих в обмотках машины при торможении (п. 4.2.5. ГОСТ IEC 60034-1). Таким образом, цикл работы T состоит из: Δtп — времени пуска, Δtр — времени работы, Δtт — времени электрического торможения, Δtотк — времени покоя (остановки или отключения питания):
4.2.5. ГОСТ IEC 60034-1). Таким образом, цикл работы T состоит из: Δtп — времени пуска, Δtр — времени работы, Δtт — времени электрического торможения, Δtотк — времени покоя (остановки или отключения питания):
T= Δtп + Δtр + Δtт + Δtотк
Условное обозначение режима аналогично предыдущему — Пример — S5 25%; Jд=0,15 кг·м²; Jнагр= 0,7 кг·м².
Действующий ГОСТ не определяет количество включений в час для режимов S4 и S5, однако в ГОСТ 183-74 оно нормировалось стандартным рядом: 30, 60, 120 и 240 включений в час.
Режим S6 — непрерывный периодический режим с кратковременной нагрузкой
Этот режим похож на S3, но при работе двигателя чередуется работа под нагрузкой Δtр и работа на холостом ходу Δt0 без остановок, то есть время покоя отсутствует (п. 4.2.6. ГОСТ IEC 60034-1). Условное обозначение — S6 40%
Режим S7 — непрерывный периодический с электрическим торможением
В этом режиме двигатель работает постоянно, но при этом выполняются одинаковые рабочие циклы, каждый из которых состоит из: Δtп — времени пуска, Δt времени работы при постоянной нагрузке и Δtт времени электрического торможения (п. 4.2.7. ГОСТ IEC 60034-1). В процессе работы двигатель остаётся включённым и время покоя отсутствует.
4.2.7. ГОСТ IEC 60034-1). В процессе работы двигатель остаётся включённым и время покоя отсутствует.
В условном обозначении указываются моменты инерции двигателя Jд и момент инерции нагрузки Jнагр — S7; 0,4 кг·м²; 7,5 кг·м² .
Режим S8 — непрерывный периодический режим со взаимозависимыми изменениями нагрузки и частоты вращения
В этом режиме двигатель работает с разными нагрузками при разных частотах вращения. Изменение частоты вращения может осуществляться переключением числа полюсов в асинхронных двигателях, использованием частотного привода и другими способами (п. 4.2.8. ГОСТ IEC 60034-1).
В условном обозначении указывают моменты инерции двигателя Jд и момент инерции нагрузки Jнагр для каждой нагрузки и частоте вращения, а также коэффициенты циклической продолжительности включения для каждой нагрузки и частоты. Пример обозначения — S8; 0,5 кг·м²; 6 кг·м²; 16 кВт 740 1/мин — 30%; 40 кВт 1460 1/мин — 30%; 25 кВт 980 1/мин — 40%.
Режим S9 — с непериодическими изменениями нагрузки и частоты вращения
В этом режиме нагрузка и частота вращения изменяются не периодически, а в случаях, когда это необходимо в технологическом процессе. Возможны перегрузки двигателя, значительно превышающие базовую нагрузку (п. 4.2.9. ГОСТ IEC 60034-1).
Режим S10 — с дискретными постоянными нагрузками и частотами вращения
В этом режиме число нагрузок и, по возможности, частот вращения ограничено, а каждая комбинация нагрузки и частоты длится достаточно долго, чтобы машина достигла практически установившегося теплового состояния (п. 4.2.10. ГОСТ IEC 60034-1). При этом возможны периоды работы машины на холостом ходу или отключение двигателя.
Условное обозначение содержит информацию о продолжительности нагрузки в виде P/Δt, где P — соответствующая нагрузка в долях базовой нагрузки и Δt — её продолжительность в долях продолжительности полного цикла нагрузки, а также относительная величина ожидаемого термического срока службы (ТСС) изоляционной системы).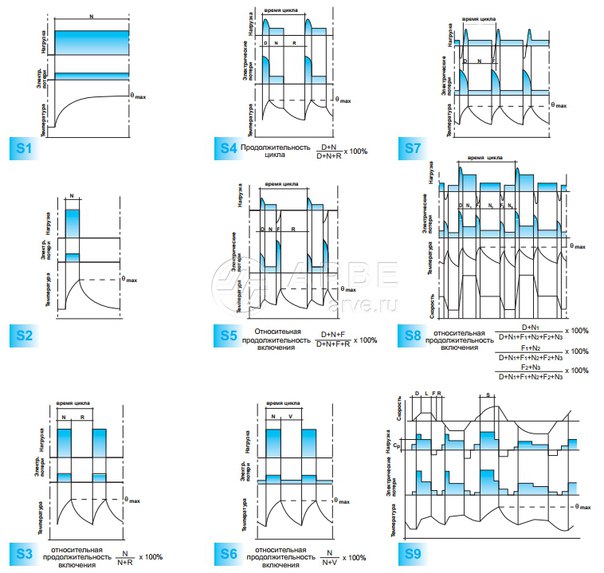 Нагрузка для времени холостого хода и обесточенного состояния машины обозначается буквой О.
Нагрузка для времени холостого хода и обесточенного состояния машины обозначается буквой О.
Пример —S10; P/Δt 1,1/0,4; 1/0,3; 0,9/0,2; O/0,1; ТСС=0,6.
Примечание — нормативной базовой величиной для оценки ожидаемого ТСС изоляции является ожидаемый термический срок службы при номинальной мощности и допускаемом пределе превышения температуры, соответствующих продолжительному типовому режиму S1.
Заключение
Двигатель, предназначенный для работы в длительном режиме S1 (или при ПВ = 100%) может работать в повторно-кратковременном и кратковременном режимах без потери мощности, и возможно использование бри нагрузке больше номинальной.
Однако двигатели, предназначенные для работы в повторно-кратковременных режимах, не могут длительно выдавать номинальную мощность, они перегреются. Длительная работа возможна либо на пониженной мощности, либо при принудительной вентиляции (обдуве).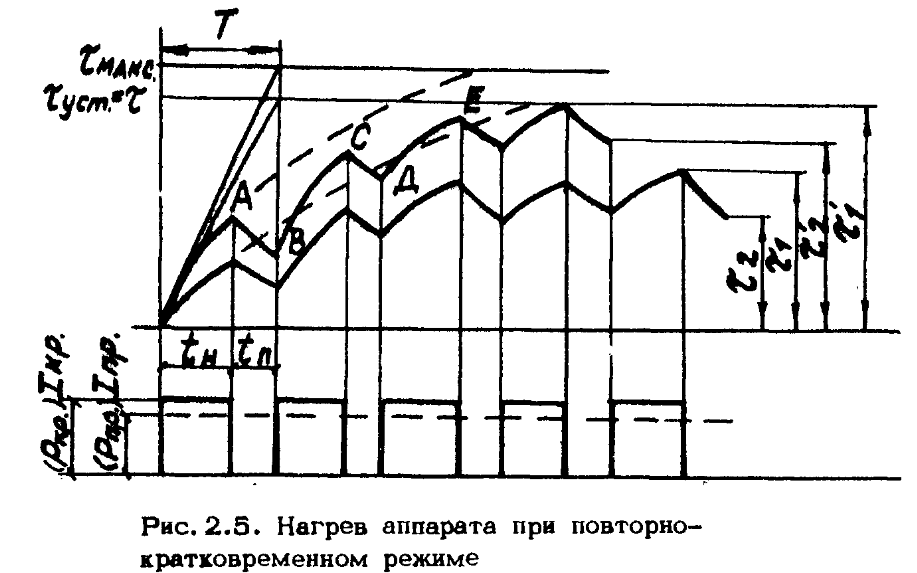
Хотелось добавить, что в старых двигателях (60-х, 70-х годов) повторно-кратковременный режим мог указываться иначе не в виде S3 40%, а в виде ПВ 40%, в чём вы можете убедиться, пересмотрев подборку фото в начале статьи. Как отмечалось выше, в то время в ГОСТе не было такого обилия режимов, их было всего 3.
1.2.2 Повторно-кратковременный режим работы. Применение тарифов на электрическую энергию и мощность для потребителей
Похожие главы из других работ:
Автоматизация электроводонагревателя ЭВ-Ф-15
1.4 Автоматический режим работы
Режим работы (ручной или автоматический) устанавливается переключателем SA1. Основной режим автоматический. Работа в ручном режиме допускается лишь при выходе из строя системы автоматики поддержания температуры…
Анализ деятельности ПАО «Газпром»
Режим работы участка
В Филиале ОАО «Газпром трансгаз Санкт- Петербург» Управление аварийно-восстановительных работ Смоленский участок службы аварийно-восстановительного поезда. установлен 8-ми часовой рабочий день.
установлен 8-ми часовой рабочий день.
* Начало рабочего дня : 8.00ч…
Изучение статической и динамической устойчивости простейшей регулируемой системы
6.1 Исходный режим работы
Исходная схема и схема замещения приведены ниже на рисунке 12.
Рисунок 12. Исходная схема и схема замещения
Результирующее сопротивление схемы , а также величину ЭДС примем согласно пункту 2.1…
Изучение статической и динамической устойчивости простейшей регулируемой системы
6.2 Аварийный режим работы
В качестве возмущающего фактора рассмотрим КЗ вначале одной из линий с последующим отключением одной из них. В схему параллельно емкостной проводимости включается дополнительный шунт…
Изучение статической и динамической устойчивости простейшей регулируемой системы
6.3 Послеаварийный режим работы
Схема послеаварийного режима работы и схема замещения приведены ниже на рисунке 16.
Рисунок 16. Схема послеаварийного режима работы и схема замещения
Сопротивление связи , определяющее амплитуду угловой характеристики. ..
..
Конденсационная паровая турбина типа К-6-4
2.1 Расчётный режим работы турбины
Расчётный режим работы турбины имеет максимальным к.п.д. преобразования энергии и определяет размеры проточной части турбины. В качестве расчётного принимается наиболее вероятный режим эксплуатации турбины в составе ПТУ, то есть такой режим…
Применение тарифов на электрическую энергию и мощность для потребителей
1.2.1 Продолжительный режим работы
В этом режиме электрические машины и аппараты могут работать длительное время без превышения температуры отдельных частей машины или аппарата выше допустимой; при этом условии обеспечивается безаварийная работа электроустановок…
Применение тарифов на электрическую энергию и мощность для потребителей
1.2.3 Кратковременный режим работы
Он характеризуется небольшими по времени периодами работы и длительными паузами с отключением электроприемника от сети. Иначе говоря, период работы имеет столь ограниченную продолжительность. ..
..
Проектирование районной электрической сети
12.1 Максимальный режим работы сети
Потери мощности в меди и стали для п- параллельно работающих двухобмоточных трансформаторов определяются по следующим выражениям:
;…
Проектирование районной электрической сети
12.2 Минимальный режим работы сети
Скорректируем нагрузки подстанций, т.е. уменьшим их на 40%:
;
;
;
;
.
Данные заносим в таблицу 12.4
Таблица 12.4 — Потери мощности в трансформаторах
п/ст
¦Sпст¦, МВА
Кол-во тр-ров
Sном тр, МВА
?Pкз, МВт
Uкз, %
?Sмтр, МВА
?Pхх, МВт
Iхх,%
?Sст тр…
Проектирование районной электрической сети
12.3 Послеаварийный режим работы сети
Точный баланс активной мощности:
Определим суммарную активную мощность нагрузок, питающихся от сети:
Для одноцепной ВЛЭП параметры ее схемы замещения: активное Rуч и индуктивное Хуч сопротивления ВЛЭП участка электрической сети…
Проектирование силовой сети производственного цеха ОАО ХБК «Шуйские ситцы» (залы №8 и №10 ткацкой фабрики №1)
1.
 2 Режим работы цеха
2 Режим работы цеха
Проектируемый участок работает по, так называемому Ивановскому графику с скользящими выходными днями. Общее число рабочих дней в году — 257, в том числе в две смены — 290; в три — 67. Средний коэффициент сменности (2 · 290 + 3 · 67)/257 = 3,03…
Расчет управляемого выпрямителя
5.1 Рабочий режим работы преобразователя
Рис. 5.1. Структурная схема проектируемой установки
Рабочий режим работы установки при номинальных (с учетом допустимых отклонений) параметрах питающей сети и нагрузки.
Действующее значение тока цепи определяется по выражению:
I1ф = Кi1КТId…
Расчет управляемого выпрямителя
5.2 Аварийный режим работы преобразователя
Аварийный режим вызывается аварийными условиями работы питающей сети, преобразователя или нагрузки и возникает ограниченное число раз за срок службы…
Технико-экономическое обоснование выбора устройств компенсации реактивной мощности и напряжения питающей линии ГПП инструментального завода
1.
 6 Экономический режим работы трансформаторов
6 Экономический режим работы трансформаторов
При эксплуатации и проектировании необходимо предусматривать экономически целесообразный режим работы трансформаторов, который определяется их параметрами и нагрузкой подстанции. Нагрузка подстанции изменяется в течение суток…
Режимы работы асинхронного двигателя | Мир инженера
Приветствую тебя, дорогой и уважаемый читатель сайта “world-engineer.ru”. В одной из статей посвященной шаровым кранам с электроприводом я затронул вопрос о работе асинхронного двигателя и пообещал рассказать о нём в другой статье. Так что в этой статье вы узнаете принцип работы асинхронного двигателя.
Асинхронный двигатель предназначен для работы в продолжительном режиме работы (условное обозначение S1), при котором нагрузка, приложенная к валу и условия охлаждения двигателя практически постоянны в промежутке времени, достаточном для нагрева двигателя до установившейся температуры.
Правильный расчет усредненных величин КПД и коэффициента мощности особенно важен при выборе геометрических размеров, расчета потерь, нагрева и охлаждения, и выборе установленной мощности стандартных двигателей, применяемых для работы в кратковременных и повторно-кратковременных режимах.
Для асинхронных двигателей совокупность кратковременных, повторно-кратковременных и перемежающихся режимов работы подразделяется на 7 групп с условными обозначениями S2…S8.
Кратковременный режим (S2), режим при, котором двигатель не успевает достигнуть номинальной температуры нагрева частей за время работы при постоянной нагрузке, и успевает охладиться до температуры окружающей среды за время пауз.
Повторно-кратковременный режим (S3) работы асинхронного двигателя, характерен для условий работы, при котором продолжительность цикла (10 мин) намного больше времени пуска (т.е. переходной процесс при пуске не влияет на нагрев двигателя и не учитываются). Продолжительность рабочего времени в течении цикла, задается в процентах от времени цикла 5%, 25%, 40% и 60%.
В режимах (S4), в отличие от (S3), продолжительность цикла настолько мала, что процессы, происходящие во время пуска (увеличение потерь из за многократного превышения величин пусковых токов над номинальным), оказывают непосредственное влияние на нагрев машины. Длительность цикла, в этом случае, определяется продолжительностью рабочего времени в процентах от времени цикла и числом включений в час. Поскольку характер переходного процесса во многом определяется динамической нагрузкой на валу, дополнительно задается допускаемый коэффициент инерции (отношение суммы моментов инерции ротора и приведенного к скорости вращения ротора, момента инерции приводного механизма к моменту инерции ротора), на который рассчитан двигатель из условий нормального нагрева.
Длительность цикла, в этом случае, определяется продолжительностью рабочего времени в процентах от времени цикла и числом включений в час. Поскольку характер переходного процесса во многом определяется динамической нагрузкой на валу, дополнительно задается допускаемый коэффициент инерции (отношение суммы моментов инерции ротора и приведенного к скорости вращения ротора, момента инерции приводного механизма к моменту инерции ротора), на который рассчитан двигатель из условий нормального нагрева.
Режимы работы асинхронного двигателя, при которых в конце каждого цикла предусмотрено электрическое торможение двигателя обозначаются (S5). Продолжительности включения в этом случае, рассчитывается с учетом времени электрического торможения.
Перемежающийся режим (S6) повторяет условия работы режима (S3) с учетом того что, в этом режиме допускается реверс с электрическим торможением или переход на другую скорость вращения. Учет пусковых потерь на нагрев двигателя не производится.
Перемежающийся режим (S7) повторяет условия работы режима (S4) с учетом того что, в этом режиме допускается реверс с электрическим торможением или переход на другую скорость вращения.
Перемежающийся режим (S8) повторяет условия работы режима (S7) с учетом того что, в этом режиме допускается работа двигателя на нескольких разных скоростях вращения с разными величинами нагрузки на валу.
Особенности определения установленной мощности в кратковременном (S2) и повторно-кратковременном режиме эксплуатации (S3, S6)
В кратковременном (S2) и повторно-кратковременных режимах эксплуатации (S3, S6) электродвигатели могут работать с большей установленной мощностью, чем в длительном режиме работы (S1).
Возможное значение величины превышения мощности рекомендуется определять из условия сохранения перегрузочной способности асинхронного двигателя по максимальному моменту в пределах:
МКР / (МN*KP) >= 1,6
При больших числах включения и больших маховых массах определение мощности рекомендуется производить исходя из:
- относительной продолжительности включения;
- частоты включения;
- величины внешнего момента инерции;
- нагрузочной диаграммы привода;
- типа торможения.

Рекомендуемые коэффициенты превышения установленной мощности
| S2 | Коэффициент превышения | S3 | Коэффициент превышения | S6 | Коэффициент превышения |
| 10 мин | 1,4 | 25% | 1,33 | 25% | 1,45 |
| 30 мин | 1,15 | 40% | 1,18 | 40% | 1,35 |
| 60% | 1,08 | 60% | 1,15 |
В настоящее время отменен ГОСТ 183-74 Машины электрические вращающиеся. Общие технические условия. И теперь взамен этого ГОСТ, действует ГОСТ Р 52776-2007 (МЭК 60034-1-2004) Машины электрические вращающиеся. Номинальные данные и характеристики.
Согласно, актуальному ГОСТ режим работы асинхронного двигателя (электрических машин) выбирается из типовых режимов от S1 до S10:
S1 – продолжительный режим. Работа асинхронного двигателя с постоянной нагрузкой и продолжительностью;
S2 – кратковременный режим. Работа асинхронного двигателя с постоянной нагрузкой в течении определенного времени. Вариант обозначения S2 60 мин;
Работа асинхронного двигателя с постоянной нагрузкой в течении определенного времени. Вариант обозначения S2 60 мин;
S3 – повторно-кратковременный периодический режим. Работа асинхронного двигателя при последовательных одинаковых рабочих циклах. Вариант обозначения S3 25%;
S4 – повторно-кратковременный периодический режим с пусками;
S5 — повторно-кратковременный периодический режим с электрическим торможением;
S6 — непрерывный периодический режим с кратковременной нагрузкой;
S7 — непрерывный периодический режим с электрическим торможением;
S8 — непрерывный периодический режим с взаимозависимыми изменениями нагрузки и частоты вращения;
S9 — режим с непериодическими изменениями нагрузки и частоты вращения;
S10 — режиме дискретными постоянными нагрузками и частотами вращения.
Более подробную информацию можете изучить в ГОСТ Р 52776-2007 (МЭК 60034-1-2004) Машины электрические вращающиеся. Номинальные данные и характеристики.
Поделиться ссылкой:
В чем разница между долговременной, кратковременной и рабочей памятью?
Прог Мозг Рес. Авторская рукопись; Доступно в PMC 2009 MAR 18.
Опубликовано в окончательной редактированной форме AS:
PMCID: PMC2657600
NIHMSID: NIHMS84208
Nelson Cowan
Департамент психологических наук Nelson, Университет Миссури, 18 Макалестер Зал, Колумбия, Мо 65211, США
Нельсон Коуэн, кафедра психологических наук, Университет Миссури, 18 Макалестер Холл, Колумбия, Миссури 65211, США;
* Автор корреспонденции. Тел.: +1 573-882-4232; Факс: +1 573-882-7710; Электронная почта: ude.iruossim@NnawoCОкончательная отредактированная версия этой статьи доступна по адресу Prog Brain Res См. другие статьи в PMC, в которых цитируется опубликованная статья.
Тел.: +1 573-882-4232; Факс: +1 573-882-7710; Электронная почта: ude.iruossim@NnawoCОкончательная отредактированная версия этой статьи доступна по адресу Prog Brain Res См. другие статьи в PMC, в которых цитируется опубликованная статья.
Abstract
В современной литературе существует значительная путаница в отношении трех типов памяти: долговременной, кратковременной и рабочей памяти. В этой главе мы попытаемся уменьшить эту путаницу и дадим современные оценки этих типов памяти. Долговременная и кратковременная память могут различаться двумя фундаментальными способами, и только кратковременная память демонстрирует (1) временное затухание и (2) ограничения емкости фрагментов.Оба свойства кратковременной памяти до сих пор вызывают споры, но современная литература довольно обнадеживает в отношении существования как распада, так и пределов емкости. Рабочая память была задумана и определена тремя разными, слегка несовместимыми способами: как кратковременная память, применяемая к когнитивным задачам, как многокомпонентная система, которая удерживает и обрабатывает информацию в кратковременной памяти, и как использование внимания для управления. краткосрочная память. Независимо от определения, есть некоторые краткосрочные показатели памяти, которые кажутся рутинными и плохо коррелируют с когнитивными способностями и другими показателями (обычно обозначаемыми термином «рабочая память»), которые кажутся более требовательными к вниманию и хорошо коррелируют. с этими способностями.Доказательства оцениваются и помещаются в теоретические рамки, изображенные на рис.
краткосрочная память. Независимо от определения, есть некоторые краткосрочные показатели памяти, которые кажутся рутинными и плохо коррелируют с когнитивными способностями и другими показателями (обычно обозначаемыми термином «рабочая память»), которые кажутся более требовательными к вниманию и хорошо коррелируют. с этими способностями.Доказательства оцениваются и помещаются в теоретические рамки, изображенные на рис.
Ключевые слова: внимание, объем оперативной памяти, контроль внимания, распад кратковременной памяти, фокус внимания, долговременная память, кратковременная память, рабочая память
Исторические корни основного научного вопроса
Сколько фаз памяти? С наивной точки зрения память можно было бы сделать из одной ткани. Некоторые люди обладают хорошей способностью запоминать факты и события, в то время как другие обладают меньшей такой способностью.Тем не менее, задолго до того, как появились настоящие психологические лаборатории, более тщательное наблюдение должно было показать, что существуют отдельные аспекты памяти. Можно увидеть, как пожилой учитель рассказывает старые уроки так же ярко, как и всегда, и все же может быть очевидно, что его способность улавливать имена новых учеников или вспоминать, какой ученик сделал какой комментарий в продолжающемся разговоре, уменьшилась за последнее время. годы.
Можно увидеть, как пожилой учитель рассказывает старые уроки так же ярко, как и всегда, и все же может быть очевидно, что его способность улавливать имена новых учеников или вспоминать, какой ученик сделал какой комментарий в продолжающемся разговоре, уменьшилась за последнее время. годы.
Научное изучение памяти обычно восходит к Герману Эббингаузу (перевод 1885/1913), который исследовал собственное усвоение и забывание новой информации в виде серии бессмысленных слогов, проверенных в разные периоды до 31 дня.Среди многих важных наблюдений Эббингауз заметил, что у него часто было «первое мимолетное схватывание… серии в моменты особой концентрации» (стр. 33), но эта непосредственная память не гарантировала, что серия была запомнена таким образом, чтобы разрешить его отзыв позже. Устойчивое запоминание иногда требовало повторения серии. Вскоре после этого Джеймс (1890) предложил различать первичную память, небольшой объем информации, хранящийся как задний край сознательного настоящего, и вторичную память, обширный объем знаний, хранящихся на протяжении всей жизни. Первичное воспоминание о Джеймсе подобно первому мимолетному постижению Эббингауза.
Первичное воспоминание о Джеймсе подобно первому мимолетному постижению Эббингауза.
Промышленная революция предъявила некоторые новые требования к тому, что Джеймс (1890) назвал первичной памятью. В 1850-х годах телеграфистам приходилось запоминать и интерпретировать быстрые серии точек и тире, передаваемые акустически. В 1876 году был изобретен телефон. Три года спустя операторы в Лоуэлле, штат Массачусетс, начали использовать телефонные номера для более чем 200 абонентов, чтобы было легче обучить подменных операторов, если четыре обычных оператора города погибнут от бушующей эпидемии кори.Такое использование телефонных номеров, дополненных префиксом слова, конечно же, распространилось. (Номер телефона автора в 1957 г. был Уайтхолл 2–6742; этот номер все еще присвоен, хотя и в виде семизначного числа.) Еще до книги Эббингауза Нифер (1878 г.) сообщил о кривой последовательного положения, полученной среди цифр в логарифмы, которые он пытался вспомнить. Можно увидеть, что бессмысленные слоги, которые Эббингауз изобрел в качестве инструмента, приобрели большую экологическую значимость в индустриальную эпоху с растущими информационными потребностями, что, возможно, подчеркивает практическую важность первичной памяти в повседневной жизни.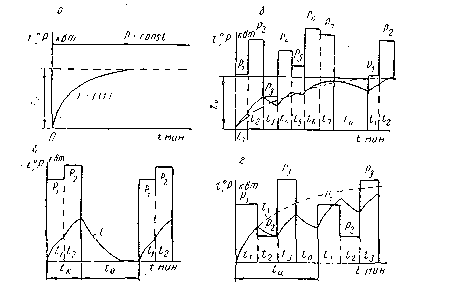 Первичная память кажется перегруженной, поскольку человека просят помнить аспекты незнакомой ситуации, такие как имена, места, вещи и идеи, с которыми он раньше не сталкивался.
Первичная память кажется перегруженной, поскольку человека просят помнить аспекты незнакомой ситуации, такие как имена, места, вещи и идеи, с которыми он раньше не сталкивался.
Тем не менее, субъективный опыт различия между первичной и вторичной памятью не гарантирует автоматически, что эти типы памяти по отдельности вносят свой вклад в науку о запоминании. Исследователи с другой точки зрения давно надеялись, что им удастся написать одно уравнение или, по крайней мере, один набор принципов, которые охватят всю память, от самых непосредственных до очень долгосрочных.McGeoch (1932) продемонстрировал, что забывание с течением времени было не просто вопросом неизбежного угасания памяти, но скорее вмешательством во время интервала удержания; можно найти ситуации, в которых память улучшалась, а не ухудшалась со временем. С этой точки зрения можно рассматривать то, что казалось забыванием из первичной памяти, как глубокое влияние вмешательства других элементов в память любого одного элемента, при этом эффекты вмешательства продолжаются вечно, но не уничтожают полностью данное воспоминание. Эта точка зрения поддерживалась и развивалась на протяжении многих лет рядом исследователей, верящих в единство памяти, включая, среди прочих, Мелтона (1963), Бьорка и Уиттена (1974), Викельгрена (1974), Краудера (1982, 1993). ), Гленберг и Суонсон (1986), Браун и др. (2000), Nairne (2002), Neath and Surprenant (2003) и Lewandowsky et al. (2004).
Эта точка зрения поддерживалась и развивалась на протяжении многих лет рядом исследователей, верящих в единство памяти, включая, среди прочих, Мелтона (1963), Бьорка и Уиттена (1974), Викельгрена (1974), Краудера (1982, 1993). ), Гленберг и Суонсон (1986), Браун и др. (2000), Nairne (2002), Neath and Surprenant (2003) и Lewandowsky et al. (2004).
Описание трех видов памяти
В этой главе я оценю силу доказательств для трех типов памяти: долговременной памяти, кратковременной памяти и рабочей памяти. Долговременная память — это огромный запас знаний и запись предшествующих событий, и она существует согласно всем теоретическим воззрениям; было бы трудно отрицать, что каждый нормальный человек имеет в своем распоряжении богатый, хотя и не безупречный и не полный набор долговременных воспоминаний.
Кратковременная память связана с первичной памятью Джеймса (1890) и представляет собой термин, который Бродбент (1958) и Аткинсон и Шиффрин (1968) использовали несколько по-разному. Подобно Аткинсону и Шиффрин, я считаю, что это отражает способности человеческого разума, которые могут временно удерживать ограниченный объем информации в очень доступном состоянии.Одно из различий между термином «кратковременная память» и термином «первичная память» заключается в том, что последний может считаться более ограниченным. Возможно, что не каждая временно доступная идея находится или даже находилась в сознании. Например, в соответствии с этой концепцией, если вы разговариваете с человеком с иностранным акцентом и непреднамеренно меняете свою речь, чтобы она соответствовала акценту иностранного говорящего, на вас влияет то, что до этого момента было бессознательным (и, следовательно, неконтролируемым) аспектом вашей короткой речи. -срочная память.Кратковременную память можно связать с паттерном возбуждения нейронов, представляющим конкретную идею, и можно считать, что эта идея находится в кратковременной памяти только тогда, когда активен паттерн возбуждения или сборка клеток (Hebb, 1949).
Подобно Аткинсону и Шиффрин, я считаю, что это отражает способности человеческого разума, которые могут временно удерживать ограниченный объем информации в очень доступном состоянии.Одно из различий между термином «кратковременная память» и термином «первичная память» заключается в том, что последний может считаться более ограниченным. Возможно, что не каждая временно доступная идея находится или даже находилась в сознании. Например, в соответствии с этой концепцией, если вы разговариваете с человеком с иностранным акцентом и непреднамеренно меняете свою речь, чтобы она соответствовала акценту иностранного говорящего, на вас влияет то, что до этого момента было бессознательным (и, следовательно, неконтролируемым) аспектом вашей короткой речи. -срочная память.Кратковременную память можно связать с паттерном возбуждения нейронов, представляющим конкретную идею, и можно считать, что эта идея находится в кратковременной памяти только тогда, когда активен паттерн возбуждения или сборка клеток (Hebb, 1949).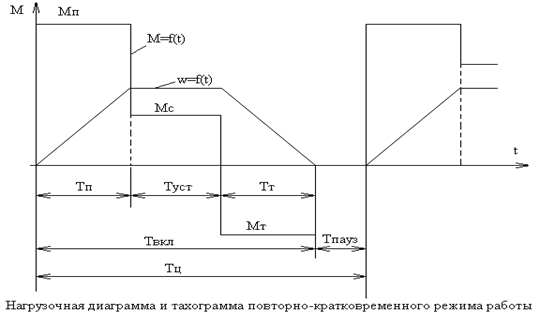 Человек может осознавать или не осознавать эту идею в течение этого периода активации.
Человек может осознавать или не осознавать эту идею в течение этого периода активации.
Рабочая память не полностью отличается от кратковременной памяти. Этот термин использовали Миллер и др. (1960) для обозначения памяти, поскольку она используется для планирования и осуществления поведения.Человек полагается на рабочую память, чтобы запомнить частичные результаты при решении арифметической задачи без бумаги, объединить предпосылки в длинном риторическом споре или испечь пирог, не совершив досадной ошибки, дважды добавив один и тот же ингредиент. (Ваша рабочая память подверглась бы более сильному напряжению при чтении предыдущего предложения, если бы я сохранил фразу «полагается на рабочую память» до конца предложения, что я и сделал в своем первом черновике этого предложения; таким образом, рабочая память влияет на хорошее письмо.) Термин «рабочая память» стал намного более доминирующим в этой области после того, как Баддели и Хитч (1974) продемонстрировали, что один модуль не может объяснить все виды временной памяти. Их мышление привело к влиятельной модели (Baddeley, 1986), в которой вербально-фонологические и визуально-пространственные репрезентации удерживались отдельно и управлялись и манипулировались с помощью связанных с вниманием процессов, называемых центральным исполнительным органом. В статье 1974 года этот центральный исполнительный орган, возможно, имел собственную память, которая пересекала области репрезентации.К 1986 году эта общая память была исключена из модели, но снова была добавлена Баддели (Baddeley, 2000) в виде эпизодического буфера . Это казалось необходимым для объяснения кратковременной памяти признаков, которые не соответствовали другим хранилищам (особенно семантической информации в памяти), и для объяснения междоменных ассоциаций в рабочей памяти, таких как сохранение связей между именами и лицами. Благодаря работе Baddeley et al. (1975), рабочая память обычно рассматривается как комбинация нескольких компонентов, работающих вместе.Некоторые даже включают в этот пакет значительный вклад долговременной памяти, которая снижает нагрузку на рабочую память за счет организации и группировки информации в рабочей памяти в меньшее количество единиц (Miller, 1956; Ericsson and Kintsch, 1995).
Их мышление привело к влиятельной модели (Baddeley, 1986), в которой вербально-фонологические и визуально-пространственные репрезентации удерживались отдельно и управлялись и манипулировались с помощью связанных с вниманием процессов, называемых центральным исполнительным органом. В статье 1974 года этот центральный исполнительный орган, возможно, имел собственную память, которая пересекала области репрезентации.К 1986 году эта общая память была исключена из модели, но снова была добавлена Баддели (Baddeley, 2000) в виде эпизодического буфера . Это казалось необходимым для объяснения кратковременной памяти признаков, которые не соответствовали другим хранилищам (особенно семантической информации в памяти), и для объяснения междоменных ассоциаций в рабочей памяти, таких как сохранение связей между именами и лицами. Благодаря работе Baddeley et al. (1975), рабочая память обычно рассматривается как комбинация нескольких компонентов, работающих вместе.Некоторые даже включают в этот пакет значительный вклад долговременной памяти, которая снижает нагрузку на рабочую память за счет организации и группировки информации в рабочей памяти в меньшее количество единиц (Miller, 1956; Ericsson and Kintsch, 1995). Например, серию букв IRSCIAFBI можно гораздо легче запомнить как серию аббревиатур трех федеральных агентств Соединенных Штатов Америки: Службы внутренних доходов (IRS), Центрального разведывательного управления (ЦРУ) и Федерального бюро Расследование (ФБР).Однако этот фактор не был подчеркнут в известной модели Baddeley (1986).
Например, серию букв IRSCIAFBI можно гораздо легче запомнить как серию аббревиатур трех федеральных агентств Соединенных Штатов Америки: Службы внутренних доходов (IRS), Центрального разведывательного управления (ЦРУ) и Федерального бюро Расследование (ФБР).Однако этот фактор не был подчеркнут в известной модели Baddeley (1986).
Из моего определения ясно, что рабочая память включает в себя кратковременную память и другие механизмы обработки, которые помогают использовать кратковременную память. Это определение отличается от того, которое используют некоторые другие исследователи (например, Engle, 2002), которые хотели бы зарезервировать термин «рабочая память» для обозначения только связанных с вниманием аспектов кратковременной памяти. Это, однако, не столько спор о содержании, сколько немного сбивающее с толку несоответствие в использовании терминов.
Одной из причин использования термина «рабочая память» является то, что было обнаружено, что показатели рабочей памяти коррелируют с интеллектуальными способностями (и особенно подвижным интеллектом) лучше, чем показатели кратковременной памяти, и фактически, возможно, лучше, чем показатели любых других конкретного психологического процесса (например, Daneman and Carpenter, 1980; Kyllonen and Christal, 1990; Daneman and Merikle, 1996; Engle et al. , 1999; Conway et al., 2005). Считалось, что это отражает использование мер, которые включают не только хранение, но и обработку, причем идея состоит в том, что и хранение, и обработка должны быть задействованы одновременно для оценки емкости рабочей памяти таким образом, чтобы это было связано с когнитивными способностями.Совсем недавно Engle et al. (1999) ввели понятие о том, что способности и рабочая память зависят от способности контролировать внимание или применять контроль внимания к управлению как первичной, так и вторичной памятью (Unsworth and Engle, 2007). Однако необходимы дополнительные исследования того, что именно мы узнаем из высокой корреляции между рабочей памятью и интеллектуальными способностями, и этот вопрос будет обсуждаться далее после того, как будет рассмотрен более фундаментальный вопрос о различиях между кратковременной и долговременной памятью.
, 1999; Conway et al., 2005). Считалось, что это отражает использование мер, которые включают не только хранение, но и обработку, причем идея состоит в том, что и хранение, и обработка должны быть задействованы одновременно для оценки емкости рабочей памяти таким образом, чтобы это было связано с когнитивными способностями.Совсем недавно Engle et al. (1999) ввели понятие о том, что способности и рабочая память зависят от способности контролировать внимание или применять контроль внимания к управлению как первичной, так и вторичной памятью (Unsworth and Engle, 2007). Однако необходимы дополнительные исследования того, что именно мы узнаем из высокой корреляции между рабочей памятью и интеллектуальными способностями, и этот вопрос будет обсуждаться далее после того, как будет рассмотрен более фундаментальный вопрос о различиях между кратковременной и долговременной памятью.
Тем временем может оказаться полезным обобщить теоретическую основу (Cowan, 1988, 1995, 1999, 2001, 2005), основанную на прошлых исследованиях. Эта схема, показанная на рисунке, помогает объяснить связь между механизмами долговременной, кратковременной и рабочей памяти и объясняет то, что я вижу как связь между ними. В этой структуре кратковременная память является производной от временно активированного подмножества информации в долговременной памяти. Это активированное подмножество может распасться со временем, если оно не будет обновлено, хотя доказательства распада все еще в лучшем случае предварительные.Подмножество активированной информации находится в центре внимания, которое, по-видимому, ограничено вместимостью фрагмента (сколько отдельных элементов может быть включено одновременно). Новые ассоциации между активированными элементами могут оказаться в центре внимания. Теперь будут обсуждаться доказательства, связанные с этой структурой моделирования.
Эта схема, показанная на рисунке, помогает объяснить связь между механизмами долговременной, кратковременной и рабочей памяти и объясняет то, что я вижу как связь между ними. В этой структуре кратковременная память является производной от временно активированного подмножества информации в долговременной памяти. Это активированное подмножество может распасться со временем, если оно не будет обновлено, хотя доказательства распада все еще в лучшем случае предварительные.Подмножество активированной информации находится в центре внимания, которое, по-видимому, ограничено вместимостью фрагмента (сколько отдельных элементов может быть включено одновременно). Новые ассоциации между активированными элементами могут оказаться в центре внимания. Теперь будут обсуждаться доказательства, связанные с этой структурой моделирования.
Различие между краткосрочной и долговременной памятью
Если есть разница между кратковременной и долговременной памятью, то есть два возможных способа, которыми эти хранилища могут различаться: в продолжительность и в емкость . Разница в длительности означает, что элементы в краткосрочном хранении разрушаются от такого типа хранения в зависимости от времени. Разница вместимости означает, что существует ограничение на количество предметов, которые могут храниться в краткосрочном хранилище. Если имеется только ограничение по вместимости, то количество предметов, меньшее предела вместимости, может оставаться на краткосрочном хранении до тех пор, пока они не будут заменены другими предметами. Оба типа ограничений являются спорными. Таким образом, чтобы оценить полезность концепции краткосрочного хранения, в свою очередь будут оцениваться пределы продолжительности и вместимости.
Разница в длительности означает, что элементы в краткосрочном хранении разрушаются от такого типа хранения в зависимости от времени. Разница вместимости означает, что существует ограничение на количество предметов, которые могут храниться в краткосрочном хранилище. Если имеется только ограничение по вместимости, то количество предметов, меньшее предела вместимости, может оставаться на краткосрочном хранении до тех пор, пока они не будут заменены другими предметами. Оба типа ограничений являются спорными. Таким образом, чтобы оценить полезность концепции краткосрочного хранения, в свою очередь будут оцениваться пределы продолжительности и вместимости.
Пределы длительности
Понятие кратковременной памяти, ограниченной затуханием во времени, присутствовало еще на заре когнитивной психологии, например в работе Бродбента (1958). Если бы затухание было единственным принципом, влияющим на производительность в эксперименте с непосредственной памятью, возможно, было бы легко обнаружить это затухание.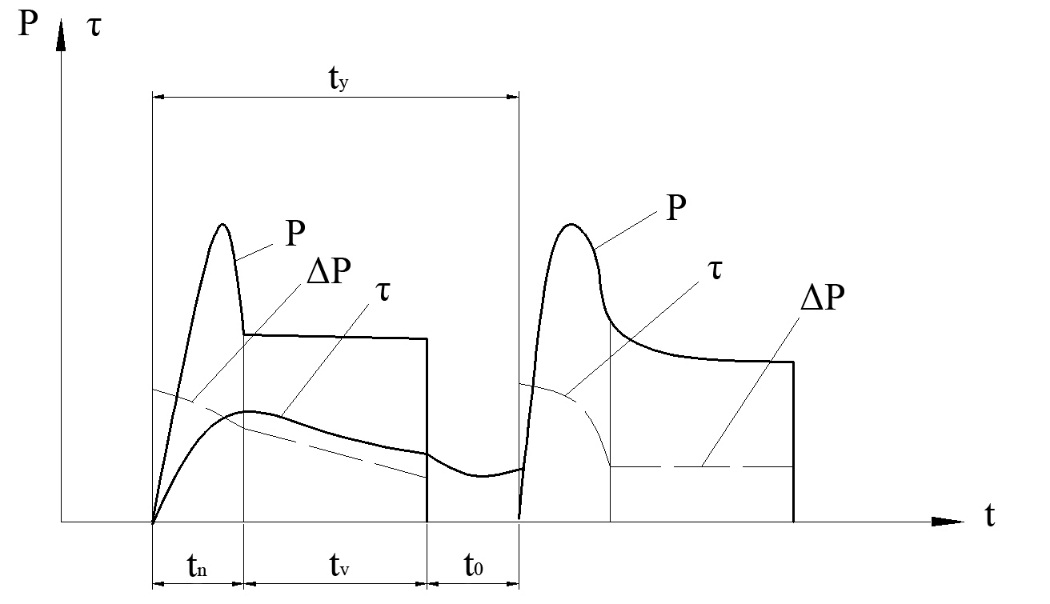 Однако даже в работе Бродбента были обнаружены контаминирующие переменные. Чтобы оценить распад, нужно принять во внимание или преодолеть загрязняющие эффекты репетиции, долговременного поиска и временной различимости, которые будут обсуждаться по одному в связи с доказательствами за и против распада.
Однако даже в работе Бродбента были обнаружены контаминирующие переменные. Чтобы оценить распад, нужно принять во внимание или преодолеть загрязняющие эффекты репетиции, долговременного поиска и временной различимости, которые будут обсуждаться по одному в связи с доказательствами за и против распада.
Преодоление контаминации от репетиции
По мнению различных исследователей, существует процесс, посредством которого человек представляет себе, как произносятся слова из списка, не произнося их вслух, процесс, называемый скрытой вербальной репетицией. С практикой этот процесс начинает происходить с минимумом внимания. Guttentag (1984) использовал второстепенное задание, чтобы показать, что повторение списка, который нужно вспомнить, требовало усилий у маленьких детей, но не у взрослых. Если в какой-то конкретной экспериментальной процедуре не наблюдается потери кратковременной памяти, то такой паттерн реакции можно отнести к репетиции.Поэтому были предприняты шаги для устранения репетиции с помощью процесса, называемого артикуляционным подавлением, при котором простое высказывание, такое как слово «the», неоднократно произносится участником в течение части или всего задания на кратковременную память (например, Baddeley et al. др., 1975). Все еще возможно возражение, что любое высказывание, используемое для подавления репетиции, к сожалению, вызывает помехи, которые могут быть истинной причиной потери памяти с течением времени, а не распада.
др., 1975). Все еще возможно возражение, что любое высказывание, используемое для подавления репетиции, к сожалению, вызывает помехи, которые могут быть истинной причиной потери памяти с течением времени, а не распада.
Эта проблема интерференции может показаться спорной в свете открытий Левандовски и др.(2004). Они представляли списки букв, которые нужно было вспомнить, и варьировали время, которое участник должен был потратить на то, чтобы вспомнить каждый элемент списка. В некоторых случаях они добавляли артикуляционное подавление, чтобы предотвратить репетицию. Несмотря на это подавление, они не наблюдали разницы в производительности со временем между элементами в ответе, которое варьировалось от 400 до 1600 мс (или между условиями, в которых слово «супер» произносилось один, два или три раза между последовательными элементами в ответе). ).Они не нашли никаких признаков ухудшения памяти.
Ограничение этого вывода, однако, заключается в том, что скрытая словесная репетиция может быть не единственным типом репетиции, которую могут использовать участники. Возможно, есть типы, которым не препятствует артикуляционное подавление. В частности, Коуэн (1992) предположил, что процесс мысленного внимания к словам или поиска в списке, требующий внимания, может служить для повторной активации элементов, которые необходимо вспомнить, аналогично скрытой вербальной репетиции.Ключевое отличие состоит в том, что нельзя ожидать, что артикуляционное подавление помешает этому типу репетиций. Вместо этого, чтобы предотвратить такую репетицию, необходимо было бы использовать задание, требующее внимания.
Возможно, есть типы, которым не препятствует артикуляционное подавление. В частности, Коуэн (1992) предположил, что процесс мысленного внимания к словам или поиска в списке, требующий внимания, может служить для повторной активации элементов, которые необходимо вспомнить, аналогично скрытой вербальной репетиции.Ключевое отличие состоит в том, что нельзя ожидать, что артикуляционное подавление помешает этому типу репетиций. Вместо этого, чтобы предотвратить такую репетицию, необходимо было бы использовать задание, требующее внимания.
Барруйе и др. (2004, 2007) имеют результаты, которые, кажется, предполагают, что существует другой, более требующий внимания тип репетиции. Между элементами, требующими выбора, они вставили материалы, которые нужно вспомнить; это могут быть числа для чтения вслух или время реакции с несколькими вариантами ответов.Обнаружено, что они мешают удержанию в памяти в той степени, в какой доля межпредметного интервала используется для внимания к отвлекающим предметам. По мере увеличения количества отвлекающих предметов вспоминается меньше предметов, которые нужно вспомнить. Идея состоит в том, что когда отвлекающая задача не требует внимания, высвобожденное внимание позволяет репетировать элементы, которые нужно вспомнить, на основе внимания. Когда вставленная задача более автоматизирована и не требует особого внимания (например,g., задание на артикуляционное подавление) влияние скорости этих вставок гораздо меньше.
По мере увеличения количества отвлекающих предметов вспоминается меньше предметов, которые нужно вспомнить. Идея состоит в том, что когда отвлекающая задача не требует внимания, высвобожденное внимание позволяет репетировать элементы, которые нужно вспомнить, на основе внимания. Когда вставленная задача более автоматизирована и не требует особого внимания (например,g., задание на артикуляционное подавление) влияние скорости этих вставок гораздо меньше.
Исходя из этой логики, можно представить вариант задачи Левандовского, в которой между элементами ответа помещаются не артикуляционное подавление, а требующие внимания вербальные стимулы, и в котором продолжительность этого заполненного времени между элементами ответа варьируется от суд за судом. Вербальные стимулы, требующие внимания, должны препятствовать как репетиции, основанной на внимании, так и репетиции, основанной на артикуляции.Если есть затухание, то производительность должна снижаться по последовательным позициям более серьезно, когда между элементами в ответе помещаются более длинные заполненные интервалы. Однако, к сожалению, такие результаты могут быть объяснены как результат интерференции отвлекающих стимулов без необходимости вызывать распад.
Однако, к сожалению, такие результаты могут быть объяснены как результат интерференции отвлекающих стимулов без необходимости вызывать распад.
То, что кажется необходимым, — это процедура, предотвращающая репетицию как на основе артикуляции, так и на основе внимания, без создания помех. Коуэн и Обюшон (в печати) опробовали один тип процедуры, с помощью которой это можно сделать.Они представили списки из семи печатных цифр, в которых время между элементами варьировалось в пределах списка. В дополнение к некоторым случайным спискам-заполнителям, было четыре критических типа испытаний, в которых все шесть пустых интервалов между цифрами были короткими (0,5 с после каждого элемента) или все длинными (2 с после каждого элемента), или состояли из трех коротких интервалов. а затем три длинных интервала или три длинных, а затем три коротких интервала. Кроме того, было два сигнала ответа после списка. Согласно одной реплике, участник должен был вспомнить список с элементами в представленном порядке, но во всяком случае они хотели. В соответствии с другой реакцией список должен был быть вызван в то же время, в которое он был представлен. Ожидалось, что необходимость помнить время в последнем состоянии реакции предотвратит репетицию любого типа. Как следствие, производительность должна быть снижена в испытаниях, в которых первые три интервала ответа длинные, потому что в этих испытаниях есть больше времени для того, чтобы забыть большинство элементов списка. Как и предсказывалось, существовала значительная взаимосвязь между сигналом реакции и продолжительностью первой половины интервалов реакции.Когда участники могли свободно вспоминать элементы в своем собственном темпе, результаты были не лучше при короткой первой половине (90 005 M 90 006 = 0,71), чем при длинной первой половине (90 005 M = 0,74). Небольшое преимущество длинной первой половины в этой ситуации может быть связано с тем, что это позволяет отрепетировать список в самом начале ответа. Напротив, когда время припоминания должно было совпадать со временем представления списка, производительность была лучше с короткой первой половиной ( M = 0,70), чем с длинной первой половиной ( M = 0,70).
В соответствии с другой реакцией список должен был быть вызван в то же время, в которое он был представлен. Ожидалось, что необходимость помнить время в последнем состоянии реакции предотвратит репетицию любого типа. Как следствие, производительность должна быть снижена в испытаниях, в которых первые три интервала ответа длинные, потому что в этих испытаниях есть больше времени для того, чтобы забыть большинство элементов списка. Как и предсказывалось, существовала значительная взаимосвязь между сигналом реакции и продолжительностью первой половины интервалов реакции.Когда участники могли свободно вспоминать элементы в своем собственном темпе, результаты были не лучше при короткой первой половине (90 005 M 90 006 = 0,71), чем при длинной первой половине (90 005 M = 0,74). Небольшое преимущество длинной первой половины в этой ситуации может быть связано с тем, что это позволяет отрепетировать список в самом начале ответа. Напротив, когда время припоминания должно было совпадать со временем представления списка, производительность была лучше с короткой первой половиной ( M = 0,70), чем с длинной первой половиной ( M = 0,70). 67). Таким образом, это предполагает, что в кратковременной памяти может быть распад.
67). Таким образом, это предполагает, что в кратковременной памяти может быть распад.
Преодоление загрязнения из-за долговременного поиска
Если имеется более одного типа памяти, то все равно возникает проблема, какое хранилище предоставило информацию, лежащую в основе ответа. Нет никакой гарантии, что долговременное хранилище не будет использовано только потому, что процедура считается проверкой краткосрочного хранения. Например, в простом задании на диапазон цифр представляется ряд цифр, который следует сразу же повторить по памяти.Если эта серия оказывалась лишь незначительно отличающейся от номера телефона участника, участник мог быстро запомнить новый номер и повторить его из долговременной памяти. Теории памяти с двойным хранилищем допускают это. Хотя Бродбент (1958) и Аткинсон и Шиффрин (1968) нарисовали свои модели обработки информации в виде ряда блоков, представляющих различные хранилища памяти, причем долговременная память следует за кратковременной памятью, эти блоки не подразумевают, что память находится исключительно в одном месте. коробка или другая; их лучше интерпретировать как относительное время первого поступления информации от стимула в одно хранилище, а затем в следующее.Остается вопрос, как можно определить, исходит ли ответ из кратковременной памяти.
коробка или другая; их лучше интерпретировать как относительное время первого поступления информации от стимула в одно хранилище, а затем в следующее.Остается вопрос, как можно определить, исходит ли ответ из кратковременной памяти.
Во и Норман (1965) разработали математическую модель для достижения этой цели. Модель работала с предположением, что долговременная память имеет место для всего списка, включая плато в середине списка. Напротив, говорят, что к моменту припоминания кратковременная память остается только в конце списка. Эта модель предполагает, что для любой конкретной последовательной позиции в списке вероятность успешного краткосрочного хранения (S) и долгосрочного хранения (L) независимы, так что вероятность отзыва элемента равна S+L-SL. .
Несколько иное предположение состоит в том, что краткосрочные и долгосрочные запасы не являются независимыми, а используются взаимодополняющим образом. Наличие кратковременной памяти элемента может позволить переместить ресурсы, необходимые для долговременного запоминания, в другое место в списке. Данные кажутся более соответствующими этому предположению. В нескольких исследованиях пациентам с корсаковской амнезией и участникам нормальной контрольной группы были представлены списки, которые следует вызвать (Baddeley and Warrington, 1970; Carlesimo et al., 1995). Эти исследования показывают, что при немедленном воспоминании у лиц с амнезией работоспособность сохраняется на последних нескольких позициях в списке. Создается впечатление, что производительность в этих последовательных позициях в основном или полностью основана на кратковременном хранении, и что у пациентов с амнезией этот тип хранения не уменьшается. При отсроченном воспоминании пациенты с амнезией обнаруживают дефицит всех серийных позиций, как и следовало ожидать, если кратковременная память на конец списка утрачивается в зависимости от заполненного периода задержки (Glanzer and Cunitz, 1966).
Данные кажутся более соответствующими этому предположению. В нескольких исследованиях пациентам с корсаковской амнезией и участникам нормальной контрольной группы были представлены списки, которые следует вызвать (Baddeley and Warrington, 1970; Carlesimo et al., 1995). Эти исследования показывают, что при немедленном воспоминании у лиц с амнезией работоспособность сохраняется на последних нескольких позициях в списке. Создается впечатление, что производительность в этих последовательных позициях в основном или полностью основана на кратковременном хранении, и что у пациентов с амнезией этот тип хранения не уменьшается. При отсроченном воспоминании пациенты с амнезией обнаруживают дефицит всех серийных позиций, как и следовало ожидать, если кратковременная память на конец списка утрачивается в зависимости от заполненного периода задержки (Glanzer and Cunitz, 1966).
Преодоление загрязнения временной различимостью
Наконец, утверждалось, что потеря памяти с течением времени не обязательно является результатом распада. Вместо этого это может быть вызвано временной различимостью в поиске. Этот тип теории предполагает, что временной контекст элемента служит сигналом для извлечения этого элемента даже при свободном воспоминании. Элемент, отделенный во времени от всех других элементов, относительно отличим и легко вспоминается, в то время как элемент, который относительно близок к другим элементам, сложнее вспомнить, потому что он использует их временные сигналы для извлечения.Вскоре после того, как представлен список, самые последние элементы становятся наиболее различимыми во времени (во многом подобно различимости телефонного столба, к которому вы практически прикасаетесь, по сравнению с столбами, простирающимися дальше по дороге). В течение интервала удержания относительная различимость самых последних элементов уменьшается (во многом это похоже на то, как если бы вы находились далеко даже от последнего полюса в серии).
Вместо этого это может быть вызвано временной различимостью в поиске. Этот тип теории предполагает, что временной контекст элемента служит сигналом для извлечения этого элемента даже при свободном воспоминании. Элемент, отделенный во времени от всех других элементов, относительно отличим и легко вспоминается, в то время как элемент, который относительно близок к другим элементам, сложнее вспомнить, потому что он использует их временные сигналы для извлечения.Вскоре после того, как представлен список, самые последние элементы становятся наиболее различимыми во времени (во многом подобно различимости телефонного столба, к которому вы практически прикасаетесь, по сравнению с столбами, простирающимися дальше по дороге). В течение интервала удержания относительная различимость самых последних элементов уменьшается (во многом это похоже на то, как если бы вы находились далеко даже от последнего полюса в серии).
Хотя есть данные, которые можно интерпретировать в соответствии с различительностью, есть и то, что выглядит как диссоциация между эффектами различимости и подлинным эффектом кратковременной памяти. Это можно видеть, например, в классической процедуре Петерсона и Петерсона (1959), в которой буквенные триграммы должны вызываться сразу или только после отвлекающего задания, считая в обратном порядке от начального числа на три, в течение периода продолжительностью до 18 с. Петерсон и Петерсон обнаружили серьезную потерю памяти для буквенной триграммы по мере увеличения задержки заполнения. Однако впоследствии скептики утверждали, что потеря памяти произошла из-за того, что временная различимость текущей буквенной триграммы уменьшилась по мере увеличения заполненной задержки.В частности, было сказано, что этот эффект задержки возникает из-за увеличения задержек между тестами из-за упреждающих помех от предыдущих испытаний. В первых нескольких испытаниях задержка не имеет значения (Кеппель и Андервуд, 1962), и не наблюдается отрицательного эффекта задержки, если задержки в 5, 10, 15 и 20 с тестируются в отдельных пробных блоках (Турви и др., 1970; Грин, 1996).
Это можно видеть, например, в классической процедуре Петерсона и Петерсона (1959), в которой буквенные триграммы должны вызываться сразу или только после отвлекающего задания, считая в обратном порядке от начального числа на три, в течение периода продолжительностью до 18 с. Петерсон и Петерсон обнаружили серьезную потерю памяти для буквенной триграммы по мере увеличения задержки заполнения. Однако впоследствии скептики утверждали, что потеря памяти произошла из-за того, что временная различимость текущей буквенной триграммы уменьшилась по мере увеличения заполненной задержки.В частности, было сказано, что этот эффект задержки возникает из-за увеличения задержек между тестами из-за упреждающих помех от предыдущих испытаний. В первых нескольких испытаниях задержка не имеет значения (Кеппель и Андервуд, 1962), и не наблюдается отрицательного эффекта задержки, если задержки в 5, 10, 15 и 20 с тестируются в отдельных пробных блоках (Турви и др., 1970; Грин, 1996).
Тем не менее, при более коротких интервалах испытаний может наблюдаться настоящий эффект затухания. Баддели и Скотт (Baddeley and Scott, 1971) установили трейлер в торговом центре, чтобы протестировать большое количество участников по одному испытанию каждый, чтобы избежать упреждающего вмешательства.Они обнаружили эффект задержки теста в течение первых 5 с, но не при более длительных задержках. Тем не менее, кажется, что концепция распада еще не очень прочная и требует дальнейшего изучения. Возможно, распад на самом деле отражает не постепенное ухудшение качества следа кратковременной памяти, а внезапный коллапс в точке, которая меняется от попытки к попытке. С контролем временной различимости Cowan et al. (1997a) обнаружили, что может быть внезапный коллапс в репрезентации памяти для тона с задержкой от 5 до 10 с.
Баддели и Скотт (Baddeley and Scott, 1971) установили трейлер в торговом центре, чтобы протестировать большое количество участников по одному испытанию каждый, чтобы избежать упреждающего вмешательства.Они обнаружили эффект задержки теста в течение первых 5 с, но не при более длительных задержках. Тем не менее, кажется, что концепция распада еще не очень прочная и требует дальнейшего изучения. Возможно, распад на самом деле отражает не постепенное ухудшение качества следа кратковременной памяти, а внезапный коллапс в точке, которая меняется от попытки к попытке. С контролем временной различимости Cowan et al. (1997a) обнаружили, что может быть внезапный коллапс в репрезентации памяти для тона с задержкой от 5 до 10 с.
Пределы емкости фрагментов
Концепция пределов емкости поднималась несколько раз в истории когнитивной психологии. Миллер (1956) классно обсуждал «магическое число семь плюс-минус два» как константу в краткосрочной обработке, включая припоминание списка, абсолютное суждение и эксперименты по численной оценке. Однако его автобиографическое эссе (Miller, 1989) указывает на то, что он никогда не относился серьезно к числу семь; это был риторический прием, который он использовал, чтобы связать воедино не связанные друг с другом направления своего исследования для выступления.Хотя объем памяти у взрослых действительно составляет примерно семь элементов, нет никакой гарантии, что каждый элемент представляет собой отдельный объект. Возможно, самым важным моментом статьи Миллера (1956) было то, что несколько элементов могут быть объединены в более крупную и осмысленную единицу. Более поздние исследования показали, что ограничение вместимости обычно составляет всего три или четыре единицы (Broadbent, 1975; Cowan, 2001). Этот вывод был основан на попытке учесть стратегии, которые часто повышают эффективность использования ограниченной емкости или позволяют хранить дополнительную информацию отдельно от этой ограниченной емкости.Чтобы понять эти методы обсуждения пределов производительности, я снова упомяну три типа загрязнения.
Однако его автобиографическое эссе (Miller, 1989) указывает на то, что он никогда не относился серьезно к числу семь; это был риторический прием, который он использовал, чтобы связать воедино не связанные друг с другом направления своего исследования для выступления.Хотя объем памяти у взрослых действительно составляет примерно семь элементов, нет никакой гарантии, что каждый элемент представляет собой отдельный объект. Возможно, самым важным моментом статьи Миллера (1956) было то, что несколько элементов могут быть объединены в более крупную и осмысленную единицу. Более поздние исследования показали, что ограничение вместимости обычно составляет всего три или четыре единицы (Broadbent, 1975; Cowan, 2001). Этот вывод был основан на попытке учесть стратегии, которые часто повышают эффективность использования ограниченной емкости или позволяют хранить дополнительную информацию отдельно от этой ограниченной емкости.Чтобы понять эти методы обсуждения пределов производительности, я снова упомяну три типа загрязнения.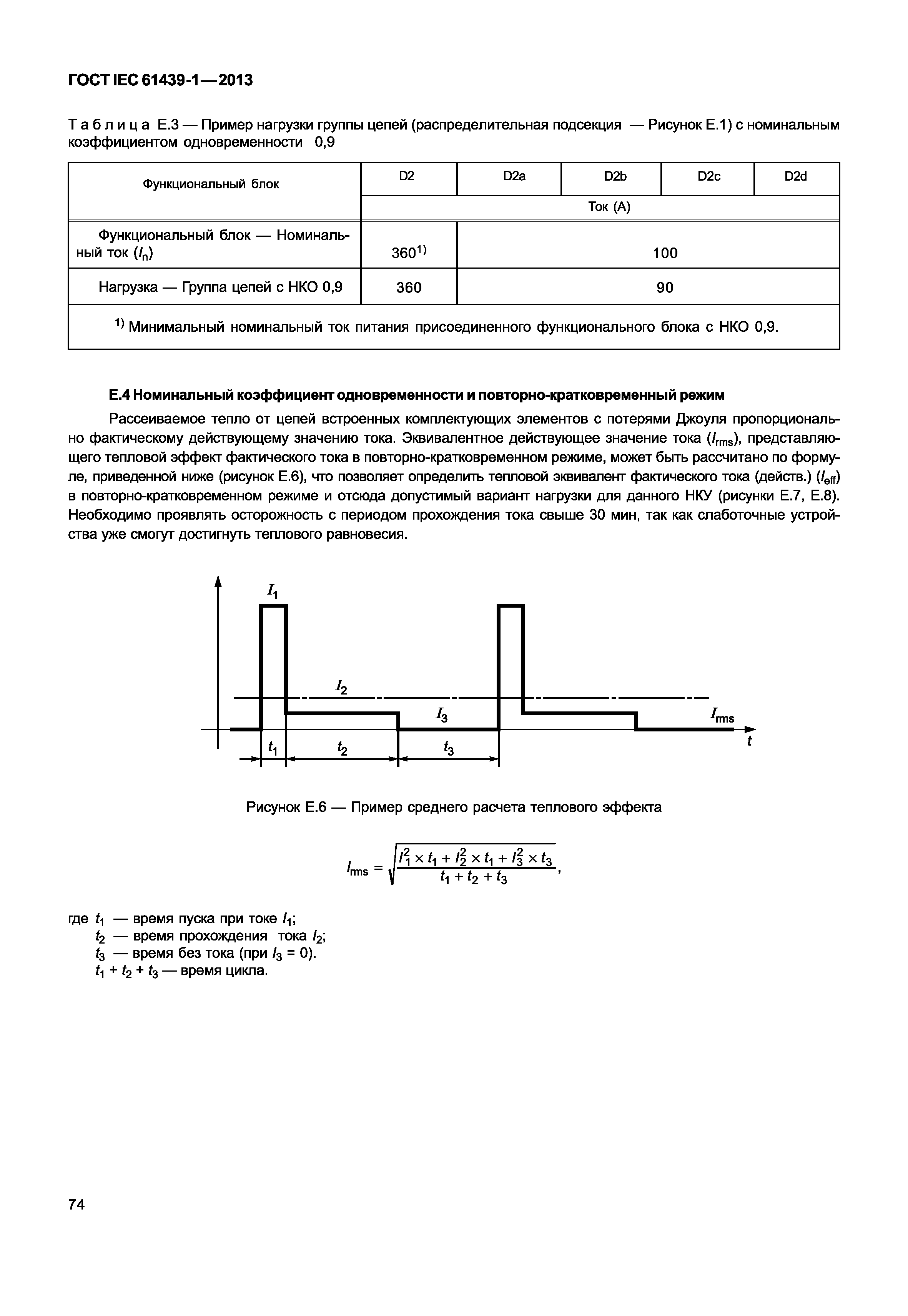 Они возникают из-за фрагментации и использования долговременной памяти, из репетиций и из типов хранения, не ограниченных по емкости.
Они возникают из-за фрагментации и использования долговременной памяти, из репетиций и из типов хранения, не ограниченных по емкости.
Преодоление загрязнения из-за фрагментации и использования долговременной памяти
Реакция участника в задании на немедленную память зависит от того, как информация, которую нужно вызвать, сгруппирована для формирования фрагментов, состоящих из нескольких элементов (Miller, 1956). Поскольку обычно неясно, какие фрагменты использовались при воспроизведении, неясно, сколько фрагментов можно сохранить и действительно ли это число фиксировано.Бродбент (1975) предложил несколько ситуаций, в которых формирование блоков из нескольких элементов не было фактором, и на основе результатов таких процедур предположил, что истинный предел емкости составляет три элемента (каждый из которых служит в качестве блока из одного элемента). Например, хотя объем памяти часто составляет около семи элементов, ошибки делаются со списками из семи элементов, а безошибочное ограничение обычно составляет три элемента. Когда люди должны вспомнить элементы из категории в долговременной памяти, например штаты США, они делают это в среднем примерно по три элемента.Это как если бы ведро кратковременной памяти наполнялось из колодца долговременной памяти и должно быть опорожнено, прежде чем оно снова наполнится. Коуэн (2001) отметил другие подобные ситуации, в которых невозможно сформировать фрагменты из нескольких элементов. Например, в рабочей области памяти длинный список элементов представлен с непредсказуемой конечной точкой, что делает невозможной группировку. Когда список заканчивается, участник должен вспомнить определенное количество предметов из конца списка. Как правило, люди могут вспомнить три или четыре элемента из конца списка, хотя точное число зависит от требований задачи (Bunting et al., 2006). Люди различаются по способностям, которые колеблются от двух до шести пунктов у взрослых (и меньше у детей), а предел индивидуальных способностей сильно коррелирует с когнитивными способностями.
Когда люди должны вспомнить элементы из категории в долговременной памяти, например штаты США, они делают это в среднем примерно по три элемента.Это как если бы ведро кратковременной памяти наполнялось из колодца долговременной памяти и должно быть опорожнено, прежде чем оно снова наполнится. Коуэн (2001) отметил другие подобные ситуации, в которых невозможно сформировать фрагменты из нескольких элементов. Например, в рабочей области памяти длинный список элементов представлен с непредсказуемой конечной точкой, что делает невозможной группировку. Когда список заканчивается, участник должен вспомнить определенное количество предметов из конца списка. Как правило, люди могут вспомнить три или четыре элемента из конца списка, хотя точное число зависит от требований задачи (Bunting et al., 2006). Люди различаются по способностям, которые колеблются от двух до шести пунктов у взрослых (и меньше у детей), а предел индивидуальных способностей сильно коррелирует с когнитивными способностями.
Еще один способ учесть роль формирования фрагментов из нескольких элементов — настроить задачу таким образом, чтобы можно было наблюдать за фрагментами. Тульвинг и Паткау (1962) изучали свободное припоминание списков слов с различными уровнями структуры, от случайных слов до правильно построенных английских предложений, с несколькими промежуточными уровнями связности.Фрагмент был определен как серия слов, воспроизведенных участником в том же порядке, в котором слова были представлены. Было подсчитано, что во всех условиях участники вспоминали в среднем от четырех до шести фрагментов. Коуэн и др. (2004) попытались усовершенствовать этот метод, проверив последовательное запоминание списков из восьми слов, которые состояли из четырех пар слов, которые ранее были связаны с различными уровнями обучения (0, 1, 2 или 4 предшествующих пары слово-слово). ). Каждое слово, использованное в списке, было представлено равное количество раз (четыре, за исключением неизученного контрольного условия), но варьировалось то, сколько из этих представлений были одиночными и сколько были последовательными парами.Количество парных предыдущих экспозиций оставалось постоянным для всех четырех пар в списке.
Тульвинг и Паткау (1962) изучали свободное припоминание списков слов с различными уровнями структуры, от случайных слов до правильно построенных английских предложений, с несколькими промежуточными уровнями связности.Фрагмент был определен как серия слов, воспроизведенных участником в том же порядке, в котором слова были представлены. Было подсчитано, что во всех условиях участники вспоминали в среднем от четырех до шести фрагментов. Коуэн и др. (2004) попытались усовершенствовать этот метод, проверив последовательное запоминание списков из восьми слов, которые состояли из четырех пар слов, которые ранее были связаны с различными уровнями обучения (0, 1, 2 или 4 предшествующих пары слово-слово). ). Каждое слово, использованное в списке, было представлено равное количество раз (четыре, за исключением неизученного контрольного условия), но варьировалось то, сколько из этих представлений были одиночными и сколько были последовательными парами.Количество парных предыдущих экспозиций оставалось постоянным для всех четырех пар в списке. Математическая модель использовалась для оценки доли припоминаемых пар, которые можно отнести к усвоенной ассоциации (т. е. фрагменту из двух слов), а не к раздельному воспоминанию двух слов в паре. Эта модель предполагала, что ограничение емкости составляло около 3,5 фрагментов в каждом условии обучения, но соотношение фрагментов из двух слов к фрагментам из одного слова увеличивалось в зависимости от количества предыдущих показов пар в списке.
Математическая модель использовалась для оценки доли припоминаемых пар, которые можно отнести к усвоенной ассоциации (т. е. фрагменту из двух слов), а не к раздельному воспоминанию двух слов в паре. Эта модель предполагала, что ограничение емкости составляло около 3,5 фрагментов в каждом условии обучения, но соотношение фрагментов из двух слов к фрагментам из одного слова увеличивалось в зависимости от количества предыдущих показов пар в списке.
Преодоление загрязнения от репетиции
Проблема репетиции не полностью отделена от проблемы формирования кусков. В традиционной концепции репетиции (например, Baddeley, 1986) предполагается, что элементы тайно артикулируются в представленном порядке в равномерном темпе. Однако другая возможность заключается в том, что репетиция включает использование артикуляционных процессов для объединения предметов в группы. Фактически, Коуэн и соавт. (2006a) спросили участников эксперимента с диапазоном цифр, как они справились с заданием, и, безусловно, наиболее распространенным ответом среди взрослых было то, что они сгруппировали элементы; участники редко упоминали о том, что говорили эти предметы про себя. Тем не менее, ясно, что подавление репетиций влияет на производительность.
Тем не менее, ясно, что подавление репетиций влияет на производительность.
Предположительно, ситуации, в которых элементы не могут быть отрепетированы, по большей части такие же, как и ситуации, в которых элементы не могут быть сгруппированы. Например, Коуэн и др. (2005) опирались на процедуру непрерывной памяти, в которой элементы предъявлялись с высокой скоростью 4 в секунду. В таком случае невозможно репетировать предметы в том виде, в каком они представлены. Вместо этого задача, вероятно, выполняется за счет сохранения пассивного хранилища (сенсорной или фонологической памяти) и последующего переноса последних нескольких элементов из этого хранилища в хранилище, более связанное с вниманием, во время припоминания.На самом деле, при высокой скорости презентации в беговом диапазоне инструкции по репетиции элементов вредны, а не полезны для производительности (Hockey, 1973). Другой пример — запоминание списков, которые были проигнорированы во время их представления (Cowan et al. , 1999). В этих случаях предел мощности близок к трем или четырем пунктам, предложенным Бродбентом (1975) и Коуэном (2001).
, 1999). В этих случаях предел мощности близок к трем или четырем пунктам, предложенным Бродбентом (1975) и Коуэном (2001).
Вполне возможно, что существует механизм краткосрочного хранения на основе речи, который в целом не зависит от механизма на основе фрагментов.В терминах популярной модели Баддели (2000) первое — это фонологическая петля, а второе — эпизодический буфер. С точки зрения Коуэна (1988, 1995, 1999, 2005), первое является частью активированной памяти, которая может иметь ограничение по времени из-за распада, а второе находится в центре внимания, которое, как предполагается, имеет предел емкости фрагмента. .
Чен и Коуэн (2005) показали, что ограничение по времени и ограничение на емкость фрагмента в кратковременной памяти различны. Они повторили процедуру Cowan et al.(2004), в которых пары слов иногда предъявлялись на тренировочном занятии, предшествующем тесту на припоминание списка. Они объединили списки, состоящие из пар, как в том исследовании. Однако теперь использовались как бесплатные, так и серийные задания на припоминание, и длина списка варьировалась.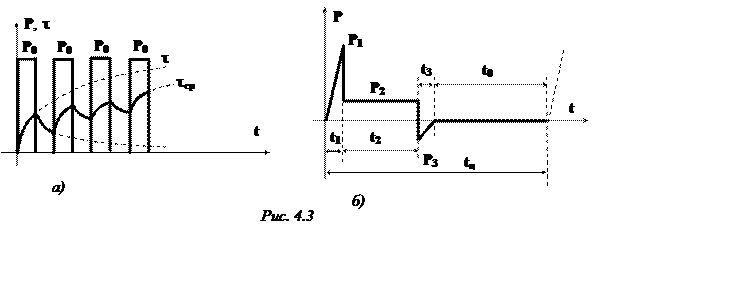 Для длинных списков и свободного отзыва ограничение на количество фрагментов регулировало отзыв. Например, вспоминались списки из шести хорошо выученных пар, а также списки из шести непарных одноэлементных слов (т. е. вспоминались с одинаковым соотношением правильных слов). Для более коротких списков и последовательного отзыва подсчитывались строгие оценки, вместо этого отзыв регулировался ограничением по времени.Например, списки из четырех хорошо заученных пар запоминались не так хорошо, как списки из четырех непарных одиночек, а лишь так же хорошо, как списки из восьми непарных одиночек. Для промежуточных условий оказалось, что ограничения емкости фрагмента и ограничения времени работают вместе, чтобы управлять отзывом. Возможно, механизм с ограниченной емкостью хранит элементы, а механизм репетиции сохраняет некоторую память последовательного порядка для этих удерживаемых элементов. Как именно эти ограничения работают вместе, пока неясно.
Для длинных списков и свободного отзыва ограничение на количество фрагментов регулировало отзыв. Например, вспоминались списки из шести хорошо выученных пар, а также списки из шести непарных одноэлементных слов (т. е. вспоминались с одинаковым соотношением правильных слов). Для более коротких списков и последовательного отзыва подсчитывались строгие оценки, вместо этого отзыв регулировался ограничением по времени.Например, списки из четырех хорошо заученных пар запоминались не так хорошо, как списки из четырех непарных одиночек, а лишь так же хорошо, как списки из восьми непарных одиночек. Для промежуточных условий оказалось, что ограничения емкости фрагмента и ограничения времени работают вместе, чтобы управлять отзывом. Возможно, механизм с ограниченной емкостью хранит элементы, а механизм репетиции сохраняет некоторую память последовательного порядка для этих удерживаемых элементов. Как именно эти ограничения работают вместе, пока неясно.
Преодоление загрязнения от неограниченных по емкости типов памяти
Трудно продемонстрировать истинный предел емкости, связанный с вниманием, если, как я полагаю, существуют другие типы механизмов кратковременной памяти, которые усложняют результаты. Общая способность должна включать в себя порции информации всех видов: например, информацию, полученную как от акустических, так и от визуальных стимулов, а также от вербальных и невербальных стимулов. Если это так, должны быть перекрестные помехи между одним типом загрузки памяти и другим. Однако литература часто показывает, что интерференция между сходными типами меморандумов, такими как два визуальных массива объектов или два акустически представленных списка слов, намного больше, чем между двумя разнородными типами, такими как один визуальный массив и один вербальный список. .Коккини и др. (2002) предположили, что различия между разнородными списками практически отсутствуют. Если это так, то это может служить аргументом против наличия общего междоменного хранилища кратковременной памяти.
Общая способность должна включать в себя порции информации всех видов: например, информацию, полученную как от акустических, так и от визуальных стимулов, а также от вербальных и невербальных стимулов. Если это так, должны быть перекрестные помехи между одним типом загрузки памяти и другим. Однако литература часто показывает, что интерференция между сходными типами меморандумов, такими как два визуальных массива объектов или два акустически представленных списка слов, намного больше, чем между двумя разнородными типами, такими как один визуальный массив и один вербальный список. .Коккини и др. (2002) предположили, что различия между разнородными списками практически отсутствуют. Если это так, то это может служить аргументом против наличия общего междоменного хранилища кратковременной памяти.
Мори и Коуэн (2004, 2005) поставили под сомнение этот вывод. Они предъявляли визуальный массив цветных пятен для сравнения со вторым массивом, совпадающим с первым или отличающимся от него цветом одного пятна. Перед первым массивом или сразу после него участники иногда слышали список цифр, который затем нужно было произнести между двумя массивами.В условиях низкой нагрузки список представлял собой их собственный семизначный телефонный номер, тогда как в условиях высокой нагрузки это был случайный семизначный номер. Только последнее условие мешало производительности сравнения массивов, и то только в том случае, если список должен был произноситься вслух между массивами. Это говорит о том, что извлечение семи случайных цифр способом, который также включает репетиционные процессы, зависит от некоторого типа механизма кратковременной памяти, который также необходим для зрительных массивов. Этот общий механизм может оказаться в центре внимания из-за его ограниченных возможностей.Очевидно, однако, что если список велся молча, а не читался вслух, это молчаливое обслуживание происходило без особого использования общего механизма хранения, основанного на внимании, поэтому производительность визуального массива не сильно влияла.
Перед первым массивом или сразу после него участники иногда слышали список цифр, который затем нужно было произнести между двумя массивами.В условиях низкой нагрузки список представлял собой их собственный семизначный телефонный номер, тогда как в условиях высокой нагрузки это был случайный семизначный номер. Только последнее условие мешало производительности сравнения массивов, и то только в том случае, если список должен был произноситься вслух между массивами. Это говорит о том, что извлечение семи случайных цифр способом, который также включает репетиционные процессы, зависит от некоторого типа механизма кратковременной памяти, который также необходим для зрительных массивов. Этот общий механизм может оказаться в центре внимания из-за его ограниченных возможностей.Очевидно, однако, что если список велся молча, а не читался вслух, это молчаливое обслуживание происходило без особого использования общего механизма хранения, основанного на внимании, поэтому производительность визуального массива не сильно влияла.
Типы кратковременной памяти, чей вклад в припоминание может скрыть предел емкости, могут включать любые типы активированной памяти, выпадающие из фокуса внимания. В структуре моделирования, изображенной на , это может включать функции сенсорной памяти, а также семантические функции.Сперлинг (1960) прекрасно проиллюстрировал разницу между неограниченной сенсорной памятью и категориальной памятью с ограниченными возможностями. Если за массивом символов вскоре после массива следовала подсказка частичного отчета, большинство символов в строке подсказки можно было вызвать. Если реплика задерживалась примерно на 1 с, большая часть сенсорной информации угасала, и производительность ограничивалась примерно четырьмя символами, независимо от размера массива. Основываясь на этом исследовании, ограничение в четыре символа можно рассматривать либо как ограничение емкости кратковременной памяти, либо как ограничение скорости, с которой информация может быть передана из сенсорной памяти в категориальную форму до ее распада. Однако Дарвин и др. (1972) провели аналогичный слуховой эксперимент и обнаружили предел около четырех элементов, хотя наблюдаемый период затухания сенсорной памяти составлял около 4 с. Учитывая поразительные различия между Sperling и Darwin et al. в период времени, доступный для перевода информации в категориальную форму, общий лимит из четырех элементов лучше всего рассматривать как ограничение пропускной способности, а не ограничение скорости.
Однако Дарвин и др. (1972) провели аналогичный слуховой эксперимент и обнаружили предел около четырех элементов, хотя наблюдаемый период затухания сенсорной памяти составлял около 4 с. Учитывая поразительные различия между Sperling и Darwin et al. в период времени, доступный для перевода информации в категориальную форму, общий лимит из четырех элементов лучше всего рассматривать как ограничение пропускной способности, а не ограничение скорости.
Солтс и Коуэн (2007) проверили эту концептуальную основу в серии экспериментов, в которых массивы были представлены в двух модальностях одновременно или, в другой процедуре, один за другим.Визуальный массив цветных пятен был дополнен массивом произнесенных цифр, воспроизводимых в четырех отдельных громкоговорителях, каждому из которых последовательно присваивался свой голос для облегчения восприятия. В некоторых испытаниях участники знали, что они несут ответственность за обе модальности одновременно, тогда как в других испытаниях участники знали, что они несут ответственность только за визуальные или только за акустические стимулы. Они получали массив зондов, который был таким же, как предыдущий массив (или такой же, как одна модальность в этом предыдущем массиве), или отличался от предыдущего массива идентичностью одного стимула.Задача состояла в том, чтобы определить, были ли изменения. Использование кросс-модального хранилища с ограниченной емкостью предсказывает конкретную модель результатов. Он предсказывает, что производительность на любой модальности должна быть снижена в условиях двойной модальности по сравнению с одномодальными условиями из-за нагрузки на хранилище кросс-модальности. Вот такие получились результаты. Более того, если бы кросс-модальное хранилище с ограниченной емкостью было единственным типом используемого хранилища, то сумма зрительных и слуховых способностей в условиях дуальной модальности не должна была бы превышать большую из двух унимодальных емкостей (что и произошло с быть зрительная способность).Причина в том, что в магазине с ограниченной вместимостью будет храниться одно и то же количество единиц, независимо от того, были ли они все из одной модальности или были из двух модальностей вместе взятых.
Они получали массив зондов, который был таким же, как предыдущий массив (или такой же, как одна модальность в этом предыдущем массиве), или отличался от предыдущего массива идентичностью одного стимула.Задача состояла в том, чтобы определить, были ли изменения. Использование кросс-модального хранилища с ограниченной емкостью предсказывает конкретную модель результатов. Он предсказывает, что производительность на любой модальности должна быть снижена в условиях двойной модальности по сравнению с одномодальными условиями из-за нагрузки на хранилище кросс-модальности. Вот такие получились результаты. Более того, если бы кросс-модальное хранилище с ограниченной емкостью было единственным типом используемого хранилища, то сумма зрительных и слуховых способностей в условиях дуальной модальности не должна была бы превышать большую из двух унимодальных емкостей (что и произошло с быть зрительная способность).Причина в том, что в магазине с ограниченной вместимостью будет храниться одно и то же количество единиц, независимо от того, были ли они все из одной модальности или были из двух модальностей вместе взятых. Это предсказание подтверждалось, но только в том случае, если в обеих модальностях сразу следовала постперцептивная маска, следующая за запоминаемым массивом. Постперцептивная маска включала разноцветное пятно в каждом месте визуального объекта и звук, состоящий из всех возможных наложенных цифр, из каждого громкоговорителя. Он предъявлялся достаточно долго после вызова массивов, чтобы их восприятие было полным (т.г., через 1 с; ср. Фогель и др., 2006). Предположительно, маска была способна перезаписывать различные типы сенсорно-специфических признаков в активированной памяти, оставляя после себя только более общую, категориальную информацию, присутствующую в фокусе внимания, которая, по-видимому, защищена от маскирующего вмешательства процесса внимания. Вновь было показано, что предел фокуса внимания составляет от трех до четырех предметов как для одномодальных зрительных, так и для бимодальных стимулов.
Это предсказание подтверждалось, но только в том случае, если в обеих модальностях сразу следовала постперцептивная маска, следующая за запоминаемым массивом. Постперцептивная маска включала разноцветное пятно в каждом месте визуального объекта и звук, состоящий из всех возможных наложенных цифр, из каждого громкоговорителя. Он предъявлялся достаточно долго после вызова массивов, чтобы их восприятие было полным (т.г., через 1 с; ср. Фогель и др., 2006). Предположительно, маска была способна перезаписывать различные типы сенсорно-специфических признаков в активированной памяти, оставляя после себя только более общую, категориальную информацию, присутствующую в фокусе внимания, которая, по-видимому, защищена от маскирующего вмешательства процесса внимания. Вновь было показано, что предел фокуса внимания составляет от трех до четырех предметов как для одномодальных зрительных, так и для бимодальных стимулов.
Даже без использования маскирующих стимулов можно найти фазу процесса кратковременной памяти, которая является общей для всех доменов. Cowan и Morey (2007) представили два набора стимулов для припоминания (или, в контрольных условиях, только один набор). Два набора стимулов могут включать в себя два произнесенных списка цифр, два пространственных массива цветных пятен или по одному каждому в любом порядке. После этой презентации подсказка указывала, что участник будет отвечать только за первый массив, только за второй массив или за оба массива. Три секунды последовали перед зондом. Влияние нагрузки на память можно сравнить двумя способами. Показатели тех испытаний, в которых предъявлялись два набора стимулов, и оба подразумевались для сохранения, можно было сравнить либо с испытаниями, в которых предъявлялся только один набор, либо его можно было сравнить с испытаниями, в которых предъявлялись оба набора, но сигнал позже указывал что только один набор должен был быть сохранен.Часть рабочей памяти, предшествующая сигналу, демонстрировала эффекты двойной задачи, специфичные для модальности: кодирование набора стимулов одного типа причиняло больше вреда при кодировании другого набора, если оба набора были в одной и той же модальности.
Cowan и Morey (2007) представили два набора стимулов для припоминания (или, в контрольных условиях, только один набор). Два набора стимулов могут включать в себя два произнесенных списка цифр, два пространственных массива цветных пятен или по одному каждому в любом порядке. После этой презентации подсказка указывала, что участник будет отвечать только за первый массив, только за второй массив или за оба массива. Три секунды последовали перед зондом. Влияние нагрузки на память можно сравнить двумя способами. Показатели тех испытаний, в которых предъявлялись два набора стимулов, и оба подразумевались для сохранения, можно было сравнить либо с испытаниями, в которых предъявлялся только один набор, либо его можно было сравнить с испытаниями, в которых предъявлялись оба набора, но сигнал позже указывал что только один набор должен был быть сохранен.Часть рабочей памяти, предшествующая сигналу, демонстрировала эффекты двойной задачи, специфичные для модальности: кодирование набора стимулов одного типа причиняло больше вреда при кодировании другого набора, если оба набора были в одной и той же модальности. Однако сохранение информации после сигнала показало эффекты двойной задачи, которые не были специфичны для модальности. Когда было представлено два набора, сохранение обоих из них было вредным по сравнению с сохранением только одного набора (как указано постстимульным сигналом удержания, чтобы сохранить один набор вместо обоих), и этот эффект двойного задания был одинаковым по величине независимо от того, наборы были в одной и той же или в разных модальностях.Таким образом, после первоначального кодирования сохранение рабочей памяти в течение нескольких секунд может происходить абстрактно, в фокусе внимания.
Однако сохранение информации после сигнала показало эффекты двойной задачи, которые не были специфичны для модальности. Когда было представлено два набора, сохранение обоих из них было вредным по сравнению с сохранением только одного набора (как указано постстимульным сигналом удержания, чтобы сохранить один набор вместо обоих), и этот эффект двойного задания был одинаковым по величине независимо от того, наборы были в одной и той же или в разных модальностях.Таким образом, после первоначального кодирования сохранение рабочей памяти в течение нескольких секунд может происходить абстрактно, в фокусе внимания.
Другое свидетельство существования отдельного краткосрочного хранилища
Наконец, есть другое свидетельство, которое прямо не поддерживает ни временное затухание, ни конкретное ограничение вместимости, но предполагает, что тот или иной из этих пределов существует. Бьорк и Уиттен (1974) и Ценг (1973) выдвинули аргументы временной различимости на основе так называемого непрерывного припоминания списка отвлекающих факторов, при котором эффект новизны сохраняется, даже когда за списком следует задержка перед припоминанием, заполненная отвлекающим фактором. Заполненная задержка должна была разрушить кратковременную память, но эффект новизны в любом случае возникает, при условии, что элементы в списке также разделены задержками, заполненными отвлекающими факторами, чтобы увеличить их отличие друг от друга. Однако в пользу краткосрочного запоминания другие исследования показали диссоциацию между тем, что обнаруживается при обычном немедленном воспроизведении, по сравнению с постоянным воспоминанием о отвлекающих факторах (например, эффекты длины слова обращаются в обратное при постоянном воспоминании о отвлекающих факторах: Cowan et al., 1997b; проактивное вмешательство в самые последние позиции списка только в постоянном воспоминании дистрактора: Craik & Birtwistle, 1971; Davelaar et al., 2005).
Заполненная задержка должна была разрушить кратковременную память, но эффект новизны в любом случае возникает, при условии, что элементы в списке также разделены задержками, заполненными отвлекающими факторами, чтобы увеличить их отличие друг от друга. Однако в пользу краткосрочного запоминания другие исследования показали диссоциацию между тем, что обнаруживается при обычном немедленном воспроизведении, по сравнению с постоянным воспоминанием о отвлекающих факторах (например, эффекты длины слова обращаются в обратное при постоянном воспоминании о отвлекающих факторах: Cowan et al., 1997b; проактивное вмешательство в самые последние позиции списка только в постоянном воспоминании дистрактора: Craik & Birtwistle, 1971; Davelaar et al., 2005).
Имеются также дополнительные данные нейровизуализации для краткосрочного хранения. Талми и др. (2005) обнаружили, что распознавание более ранних частей списка, но не последних нескольких элементов, активирует области в системе гиппокампа, которые обычно связаны с поиском в долговременной памяти. Это согласуется с упомянутым ранее выводом о том, что при амнезии Корсакова сохраняется память на несколько последних пунктов списка (Baddeley and Warrington, 1970; Carlesimo et al., 1995). В этих исследованиях часть эффекта недавности, основанная на кратковременной памяти, могла отражать короткий промежуток времени между предъявлением и воспоминанием нескольких последних предметов или отсутствие интерференции между предъявлением и воспоминанием последних нескольких предметов. .Таким образом, мы можем сказать, что кратковременная память существует, но часто без ясности в отношении того, является ли ограничение ограничением по времени или ограничением емкости фрагмента.
Это согласуется с упомянутым ранее выводом о том, что при амнезии Корсакова сохраняется память на несколько последних пунктов списка (Baddeley and Warrington, 1970; Carlesimo et al., 1995). В этих исследованиях часть эффекта недавности, основанная на кратковременной памяти, могла отражать короткий промежуток времени между предъявлением и воспоминанием нескольких последних предметов или отсутствие интерференции между предъявлением и воспоминанием последних нескольких предметов. .Таким образом, мы можем сказать, что кратковременная память существует, но часто без ясности в отношении того, является ли ограничение ограничением по времени или ограничением емкости фрагмента.
Различие между кратковременной памятью и рабочей памятью
Различие между кратковременной памятью и рабочей памятью омрачено некоторой путаницей, но в основном это результат того, что разные исследователи используют разные определения. Миллер и др. (1960) использовали термин «рабочая память» для обозначения временной памяти с функциональной точки зрения, поэтому с их точки зрения нет четкого различия между кратковременной и рабочей памятью. Baddeley и Hitch (1974) в значительной степени придерживались этого определения, но накладывали некоторые описания на термины, которые их отличали. Они думали о кратковременной памяти как о едином месте удержания, как описано, например, Аткинсоном и Шиффрином (1968). Когда они поняли, что данные на самом деле согласуются с многокомпонентной системой, которую нельзя свести к единому кратковременному хранилищу, они использовали термин «рабочая память» для описания всей этой системы. Коуэн (1988) придерживался многокомпонентной точки зрения, как и Баддели и Хитч, но без приверженности конкретным их компонентам; вместо этого основными подразделениями рабочей памяти считались компоненты кратковременного хранения (активированная память вместе с фокусом внимания в ней, показанные на рис. 1) и центральные исполнительные процессы, которые манипулируют хранимой информацией.По мнению Коуэна, фонологическая петля Баддели (Baddeley, 1986) и зрительно-пространственный блокнот должны рассматриваться как всего лишь два из многих аспектов активированной памяти, которые подвержены вмешательству в той степени, которая зависит от сходства между характеристиками активированного и мешающего источников информации.
Baddeley и Hitch (1974) в значительной степени придерживались этого определения, но накладывали некоторые описания на термины, которые их отличали. Они думали о кратковременной памяти как о едином месте удержания, как описано, например, Аткинсоном и Шиффрином (1968). Когда они поняли, что данные на самом деле согласуются с многокомпонентной системой, которую нельзя свести к единому кратковременному хранилищу, они использовали термин «рабочая память» для описания всей этой системы. Коуэн (1988) придерживался многокомпонентной точки зрения, как и Баддели и Хитч, но без приверженности конкретным их компонентам; вместо этого основными подразделениями рабочей памяти считались компоненты кратковременного хранения (активированная память вместе с фокусом внимания в ней, показанные на рис. 1) и центральные исполнительные процессы, которые манипулируют хранимой информацией.По мнению Коуэна, фонологическая петля Баддели (Baddeley, 1986) и зрительно-пространственный блокнот должны рассматриваться как всего лишь два из многих аспектов активированной памяти, которые подвержены вмешательству в той степени, которая зависит от сходства между характеристиками активированного и мешающего источников информации. Эпизодический буфер Баддели (2000), возможно, совпадает с информацией, хранящейся в фокусе внимания Коуэна, или, по крайней мере, представляет собой близкое к нему понятие.
Эпизодический буфер Баддели (2000), возможно, совпадает с информацией, хранящейся в фокусе внимания Коуэна, или, по крайней мере, представляет собой близкое к нему понятие.
Произошел некоторый сдвиг в определении или описании рабочей памяти, а также сдвиг в объяснении того, почему новые задачи на рабочую память гораздо лучше коррелируют с показателями интеллекта и способностей, чем простые традиционные задачи на кратковременную память. например серийный отзыв.Данеман и Карпентер (1980) предположили, что критически важным является использование задач рабочей памяти, которые включают в себя компоненты хранения и обработки, чтобы задействовать все части рабочей памяти, как описано, например, Баддели и Хитчем (1974). . Вместо этого Энгл и соавт. (1999) и Кейн и др. (2001) предположили, что критически важно, является ли задача рабочей памяти сложной с точки зрения контроля внимания. Например, Кейн и др. обнаружили, что объем рабочей памяти, связанный с задачами хранения и обработки, хорошо коррелирует со способностью подавлять естественную тенденцию смотреть в сторону внезапно появляющегося стимула и вместо этого смотреть в другую сторону, задачу антисаккады. Точно так же Конвей и соавт. (2001) обнаружили, что люди, набравшие высокие баллы в тестах на хранение и обработку рабочей памяти, замечают, что их имена в канале игнорируются при дихотическом слушании реже , чем люди с низким диапазоном; люди с большим размахом, по-видимому, лучше способны сделать выполнение своей основной задачи менее уязвимым для отвлечения внимания, но это происходит за счет того, что они немного не обращают внимания на несущественные аспекты своего окружения. В ответ на такое исследование Энгл и его коллеги иногда использовали термин «рабочая память» для обозначения только процессов, связанных с контролем внимания.Таким образом, их определение рабочей памяти кажется противоречащим предыдущим определениям, но это новое определение позволяет простое утверждение, что рабочая память сильно коррелирует со способностями, тогда как кратковременная память (переопределенная, чтобы включить только аспекты памяти, не связанные с вниманием). память) не так сильно коррелирует со способностями.
Точно так же Конвей и соавт. (2001) обнаружили, что люди, набравшие высокие баллы в тестах на хранение и обработку рабочей памяти, замечают, что их имена в канале игнорируются при дихотическом слушании реже , чем люди с низким диапазоном; люди с большим размахом, по-видимому, лучше способны сделать выполнение своей основной задачи менее уязвимым для отвлечения внимания, но это происходит за счет того, что они немного не обращают внимания на несущественные аспекты своего окружения. В ответ на такое исследование Энгл и его коллеги иногда использовали термин «рабочая память» для обозначения только процессов, связанных с контролем внимания.Таким образом, их определение рабочей памяти кажется противоречащим предыдущим определениям, но это новое определение позволяет простое утверждение, что рабочая память сильно коррелирует со способностями, тогда как кратковременная память (переопределенная, чтобы включить только аспекты памяти, не связанные с вниманием). память) не так сильно коррелирует со способностями.
Коуэн и др. (2006b), придерживаясь более традиционного определения рабочей памяти, сделали утверждение о рабочей памяти, похожее на утверждение Энгла и его коллег, но немного более сложное.Они предположили, на основе некоторых данных о развитии и корреляции, что множественные функции внимания имеют отношение к индивидуальным различиям в способностях. Контроль внимания имеет значение, но есть независимый вклад от количества объектов, которые можно удерживать во внимании, или от его объема. Согласно этой точке зрения, для того, чтобы процедура рабочей памяти хорошо коррелировала с когнитивными способностями, может быть необходимо, чтобы задача предотвращала скрытую вербальную репетицию, чтобы участник должен был полагаться на более требовательную к вниманию обработку и / или хранение для выполнения задачи. .Коуэн и др. (2005) обнаружили, что задача может быть намного проще, чем процедуры хранения и обработки. Например, в версии теста на бегущую память цифры предъявляются очень быстро, и серия останавливается в непредсказуемом месте, после чего участник должен вспомнить как можно больше элементов из конца списка. Повторение невозможно, и когда список заканчивается, информация, по-видимому, должна быть извлечена из активированных сенсорных или фонологических признаков в фокус внимания.Этот тип задач коррелировал со способностями, как и некоторые другие показатели объема внимания (Cowan et al., 2005, 2006b). У детей, которые слишком малы, чтобы использовать скрытую словесную репетицию (в отличие от детей старшего возраста и взрослых), даже простое задание на диапазон цифр служило превосходной корреляцией со способностями.
Повторение невозможно, и когда список заканчивается, информация, по-видимому, должна быть извлечена из активированных сенсорных или фонологических признаков в фокус внимания.Этот тип задач коррелировал со способностями, как и некоторые другие показатели объема внимания (Cowan et al., 2005, 2006b). У детей, которые слишком малы, чтобы использовать скрытую словесную репетицию (в отличие от детей старшего возраста и взрослых), даже простое задание на диапазон цифр служило превосходной корреляцией со способностями.
Другое исследование подтверждает эту идею о том, что тест на рабочую память будет хорошо коррелировать с когнитивными способностями в той мере, в какой он требует использования внимания для хранения и/или обработки.Гавенс и Барруйе (2004) провели исследование развития, в котором они контролировали сложность и продолжительность задачи обработки, возникающей между элементами, которые нужно вспомнить. По-прежнему существовала разница в продолжительности развития, которую они приписывали развитию базовой способности, что могло отражать увеличение объема внимания в процессе развития (ср.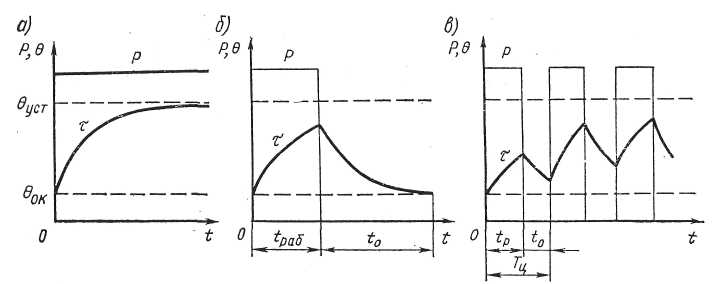 Cowan et al., 2005). Лепин и др. (2005) показали, что для хорошо коррелируемой со способностями задачи, связанной с хранением и обработкой, важно, чтобы компонент обработки задачи (в данном случае чтение букв вслух) выполнялся достаточно быстро, чтобы предотвратить различные типы репетиция, чтобы прокрасться между ними (см. также Conlin et al., 2005).
Cowan et al., 2005). Лепин и др. (2005) показали, что для хорошо коррелируемой со способностями задачи, связанной с хранением и обработкой, важно, чтобы компонент обработки задачи (в данном случае чтение букв вслух) выполнялся достаточно быстро, чтобы предотвратить различные типы репетиция, чтобы прокрасться между ними (см. также Conlin et al., 2005).
В нескольких статьях хранение и обработка данных (возможно, область действия и контроль внимания?) противопоставлены друг другу, чтобы выяснить, что более важно для учета индивидуальных различий. Фогель и др. (2005) использовали задачу визуального массива, модифицированную для использования с компонентом потенциалов, связанных с событием, который указывает на хранение в зрительной рабочей памяти, называемой активностью контралатеральной задержки (CDA). Выяснилось, что эта активность зависит не только от количества релевантных объектов на дисплее (т.например, красные полосы под разными углами, которые необходимо запомнить), но иногда и количество нерелевантных объектов, которые следует игнорировать (например, синие полосы). Было обнаружено, что для лиц с большим размахом CDA для двух релевантных объектов был одинаковым независимо от того, были ли на дисплее также два нерелевантных объекта. Однако для людей с низким размахом CDA для двух релевантных объектов в сочетании с двумя нерелевантными объектами был аналогичен CDA для дисплеев только с четырьмя релевантными объектами, как будто нерелевантные объекты нельзя было исключить из рабочей памяти.Одним из ограничений исследования является то, что разделение участников на группы с высоким и низким диапазоном также основывалось на CDA, а задача, используемая для измерения CDA, неизбежно требовала избирательного внимания (к одной половине дисплея) в каждом испытании, независимо от того, в него входили предметы несоответствующего цвета.
Было обнаружено, что для лиц с большим размахом CDA для двух релевантных объектов был одинаковым независимо от того, были ли на дисплее также два нерелевантных объекта. Однако для людей с низким размахом CDA для двух релевантных объектов в сочетании с двумя нерелевантными объектами был аналогичен CDA для дисплеев только с четырьмя релевантными объектами, как будто нерелевантные объекты нельзя было исключить из рабочей памяти.Одним из ограничений исследования является то, что разделение участников на группы с высоким и низким диапазоном также основывалось на CDA, а задача, используемая для измерения CDA, неизбежно требовала избирательного внимания (к одной половине дисплея) в каждом испытании, независимо от того, в него входили предметы несоответствующего цвета.
Голд и др. (2006) исследовали аналогичные проблемы в поведенческом дизайне и проверяли разницу между пациентами с шизофренией и нормальными участниками контрольной группы. Каждое испытание начиналось с указания обратить внимание на одну часть дисплея за счет другой (например,г. , полосы одного релевантного цвета, но не другого, нерелевантного цвета). Отображение пробы представляло собой набор, релевантность которого определялась в большинстве испытаний (в некоторых экспериментах, 75%), тогда как иногда отображение пробы представляло собой набор, на который не подавалась подсказка. Это позволило отдельно измерить контроль внимания (преимущество элементов с сигналом над элементами без сигнала) и емкость рабочей памяти (среднее количество элементов, вызванных из каждого массива, добавленное к наборам с сигналом и без сигнала). В отличие от первоначальных ожиданий явный результат заключался в том, что разница между группами заключалась в способности, а не в контроле внимания.Было бы интересно узнать, можно ли получить такой же результат для нормальных людей с высоким и низким размахом, или вместо этого сравнение покажет разницу в контроле внимания между этими группами, как Vogel et al. (2005) должен предсказать. Фридман и др. (2006) обнаружили, что не все центральные исполнительные функции коррелируют со способностями; обновление рабочей памяти помогло, а торможение и переключение внимания — нет.
, полосы одного релевантного цвета, но не другого, нерелевантного цвета). Отображение пробы представляло собой набор, релевантность которого определялась в большинстве испытаний (в некоторых экспериментах, 75%), тогда как иногда отображение пробы представляло собой набор, на который не подавалась подсказка. Это позволило отдельно измерить контроль внимания (преимущество элементов с сигналом над элементами без сигнала) и емкость рабочей памяти (среднее количество элементов, вызванных из каждого массива, добавленное к наборам с сигналом и без сигнала). В отличие от первоначальных ожиданий явный результат заключался в том, что разница между группами заключалась в способности, а не в контроле внимания.Было бы интересно узнать, можно ли получить такой же результат для нормальных людей с высоким и низким размахом, или вместо этого сравнение покажет разницу в контроле внимания между этими группами, как Vogel et al. (2005) должен предсказать. Фридман и др. (2006) обнаружили, что не все центральные исполнительные функции коррелируют со способностями; обновление рабочей памяти помогло, а торможение и переключение внимания — нет.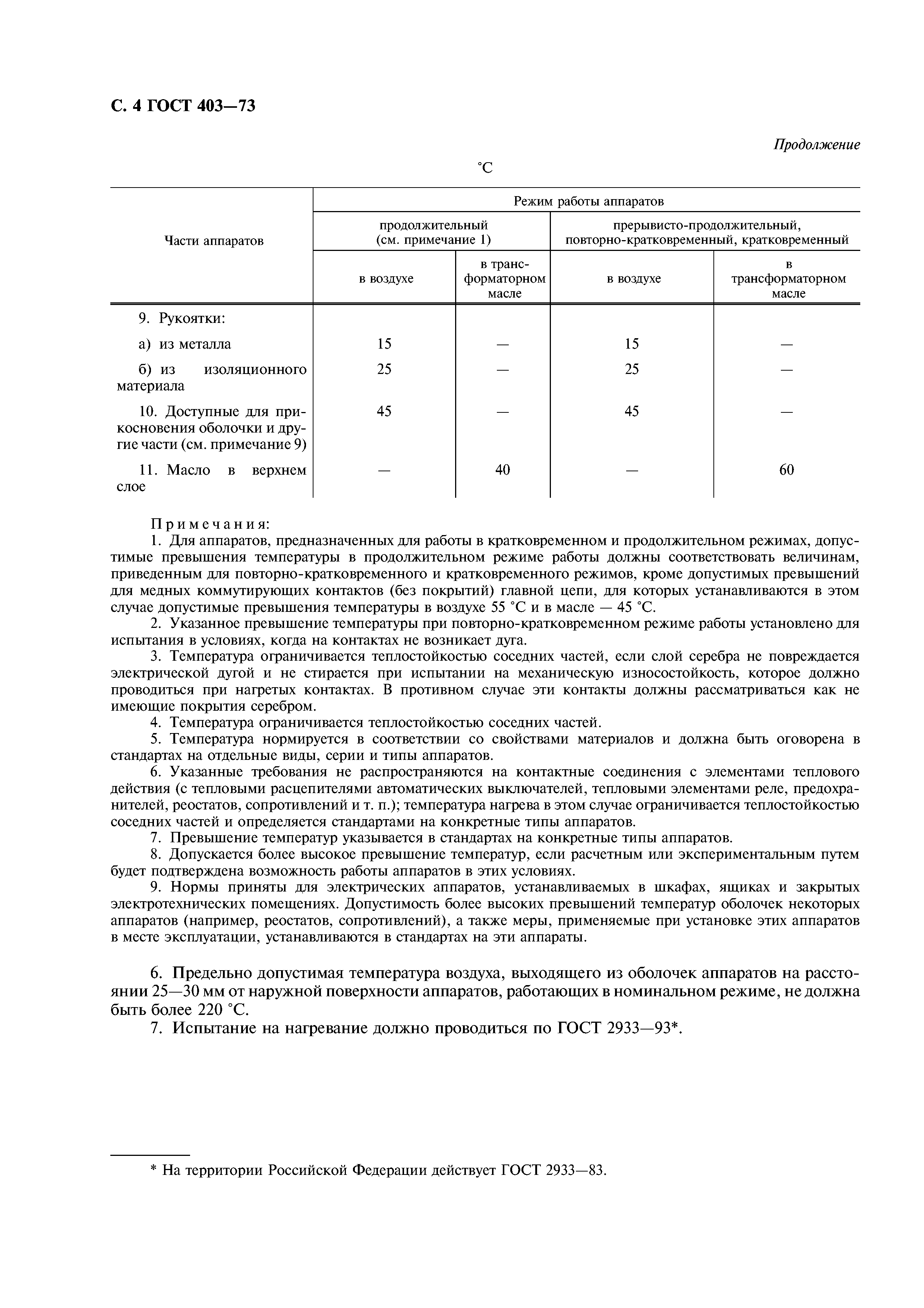 С другой стороны, напомним, что Cowan et al. (2006b) обнаружили, что задача на контроль внимания связана со способностями.
С другой стороны, напомним, что Cowan et al. (2006b) обнаружили, что задача на контроль внимания связана со способностями.
Таким образом, вопрос о том, различаются ли кратковременная память и рабочая память, может быть вопросом семантики. Существуют явные различия между простыми последовательными задачами на припоминание, которые не очень хорошо коррелируют с тестами способностей у взрослых, и другими задачами, требующими памяти и обработки или памяти без возможности репетиции, которые гораздо лучше коррелируют со способностями. Использовать ли термин «рабочая память» для последнего набора задач или зарезервировать этот термин для всей системы сохранения кратковременной памяти и манипулирования ею — дело вкуса.Более важный и содержательный вопрос может заключаться в том, почему одни задачи коррелируют со способностями гораздо лучше, чем другие.
Заключение
Различие между долговременной и кратковременной памятью зависит от того, можно ли продемонстрировать наличие свойств, специфичных для кратковременной памяти; основные кандидаты включают временное затухание и ограничение емкости фрагмента. Вопрос распада все еще остается открытым для обсуждения, в то время как растет поддержка ограничения емкости блока. Эти ограничения обсуждались в рамках, показанных на рис.
Вопрос распада все еще остается открытым для обсуждения, в то время как растет поддержка ограничения емкости блока. Эти ограничения обсуждались в рамках, показанных на рис.
Различие между кратковременной памятью и рабочей памятью зависит от принятого определения. Тем не менее, существенный вопрос заключается в том, почему одни тесты на кратковременную память служат одними из лучших коррелятов когнитивных способностей, а другие — нет. Ответ, кажется, указывает на важность системы внимания, используемой как для обработки, так и для хранения. Эффективность этой системы и ее использование в рабочей памяти существенно различаются у разных людей (например,г., Конвей и др., 2002; Кейн и др., 2004 г.; Cowan et al., 2005, 2006b), а также улучшение по мере развития в детстве (Cowan et al., 2005, 2006b) и снижение в пожилом возрасте (Naveh-Benjamin et al., 2007; Stoltzfus et al., 1996; Коуэн и др., 2006c).
Благодарность
Эта работа была выполнена при содействии гранта NIH R01 HD-21338.
Ссылки
- Аткинсон Р.С., Шиффрин Р.М. Человеческая память: предлагаемая система и процессы управления ею. В: Spence KW, Spence JT, редакторы.Психология обучения и мотивации: достижения в исследованиях и теории. Том. 2. Нью-Йорк: Академик Пресс; 1968. С. 89–195. [Google Scholar]
- Баддели А. Эпизодический буфер: новый компонент рабочей памяти? Тенденции Познан. науч. 2000;4:417–423. [PubMed] [Google Scholar]
- Baddeley AD. Оксфордская серия психологии № 11. Оксфорд: Clarendon Press; 1986. Рабочая память. [Google Scholar]
- Baddeley AD, Hitch G. Рабочая память. В: Bower GH, редактор. Психология обучения и мотивации.Том. 8. Нью-Йорк: Академик Пресс; 1974. С. 47–89. [Google Scholar]
- Baddeley AD, Scott D. Кратковременное забывание при отсутствии упреждающего торможения. QJ Exp. Психол. 1971; 23: 275–283. [Google Scholar]
- Баддели А.Д., Томсон Н., Бьюкенен М. Длина слова и структура кратковременной памяти. J. Вербальное обучение. Вербальное поведение. 1975; 14: 575–589. [Google Scholar]
- Baddeley AD, Warrington EK. Амнезия и различие между долговременной и кратковременной памятью. J. Вербальное обучение.Вербальное поведение. 1970; 9: 176–189. [Google Scholar]
- Barrouillet P, Bernardin S, Camos V. Ограничения по времени и совместное использование ресурсов в области рабочей памяти взрослых. Дж. Эксп. Психология: Ген. 2004; 133: 83–100. [PubMed] [Google Scholar]
- Barrouillet P, Bernardin S, Portrat S, Vergauwe E, Camos V. Время и когнитивная нагрузка на рабочую память. Дж. Эксп. Психол. Учиться. Мем. Познан. 2007; 33: 570–585. [PubMed] [Google Scholar]
- Бьорк Р.А., Уиттен В.Б. Чувствительные к новизне процессы поиска в долгосрочном свободном воспоминании.Познан. Психол. 1974; 6: 173–189. [Google Scholar]
- Broadbent DE. Восприятие и общение. Нью-Йорк: Пергамон Пресс; 1958. [Google Scholar]
- Broadbent DE. Магическое число семь через пятнадцать лет. В: Кеннеди А. , Уилкс А., редакторы. Исследования долговременной памяти. Оксфорд, Англия: Wiley; 1975. С. 3–18. [Google Scholar]
- Brown GDA, Preece T, Hulme C. Память на основе осциллятора для последовательного порядка. Психол. 2000; 107:127–181. [PubMed] [Google Scholar]
- Бантинг М.Ф., Коуэн Н., Солтс Дж.С.Как работает оперативная память? QJ Exp. Психол. 2006; 59: 1691–1700. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Карлезимо Г.А., Саббадини М., Фадда Л., Кальтаджироне К. Различные компоненты в словарном списке забывают чистых амнезиаков, дегенеративных слабоумных и здоровых субъектов. кора. 1995; 31: 735–745. [PubMed] [Google Scholar]
- Чен З., Коуэн Н. Ограничения фрагментов и длины при немедленном воспроизведении: согласование. Дж. Эксп. Психол. Учиться. Мем. Познан. 2005; 31: 1235–1249. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cocchini G, Logie RH, Della Sala S, MacPherson SE, Baddeley AD.Параллельное выполнение двух задач памяти: свидетельство существования систем рабочей памяти, специфичных для предметной области. Мем. Познан. 2002; 30:1086–1095. [PubMed] [Google Scholar]
- Conlin JA, Gathercole SE, Adams JW. Детская рабочая память: исследование ограничений производительности в сложных задачах. Дж. Эксп. Детская психология. 2005;90:303–317. [PubMed] [Google Scholar]
- Conway ARA, Cowan N, Bunting MF. Новый взгляд на феномен коктейльной вечеринки: важность объема рабочей памяти. Психон. Бык.2001; 8: 331–335. [PubMed] [Google Scholar]
- Conway ARA, Cowan N, Bunting MF, Therriault DJ, Minkoff SRB. Скрытый переменный анализ объема рабочей памяти, объема кратковременной памяти, скорости обработки и общего подвижного интеллекта. Интеллект. 2002; 30: 163–183. [Google Scholar]
- Conway ARA, Kane MJ, Bunting MF, Hambrick DZ, Wilhelm O, Engle RW. Задания на объем рабочей памяти: методический обзор и руководство пользователя. Психон. Бык. 2005; 12:769–786. [PubMed] [Google Scholar]
- Коуэн Н.Развитие концепций хранения памяти, избирательного внимания и их взаимных ограничений в системе обработки информации человека. Психол. Бык. 1988; 104: 163–191. [PubMed] [Google Scholar]
- Коуэн Н. Объем вербальной памяти и время устного воспроизведения. Дж. Мем. Ланг. 1992; 31: 668–684. [Google Scholar]
- Коуэн Н. Оксфордская серия психологии № 26. Нью-Йорк: Издательство Оксфордского университета; 1995. Внимание и память: интегрированная структура. [Google Scholar]
- Коуэн Н.Модель встроенных процессов рабочей памяти. В: Мияке А., Шах П., редакторы. Модели рабочей памяти: механизмы активного обслуживания и исполнительного контроля. Кембридж, Великобритания: Издательство Кембриджского университета; 1999. С. 62–101. [Google Scholar]
- Коуэн Н. Волшебное число 4 в кратковременной памяти: переосмысление умственной емкости памяти. Поведение наук о мозге. 2001; 24:87–185. [PubMed] [Google Scholar]
- Коуэн Н. Объем оперативной памяти. Хоув, Восточный Суссекс, Великобритания: Psychology Press; 2005.[Google Scholar]
- Коуэн Н., Обюшон А.М. Психон. Бык. Rev. Кратковременная потеря памяти с течением времени без вмешательства ретроактивных стимулов. (в печати) [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Chen Z, Rouder JN. Постоянная способность в непосредственной задаче припоминания серий: логическое продолжение Миллера (1956) Psychol. науч. 2004; 15: 634–640. [PubMed] [Google Scholar]
- Cowan N, Elliott EM, Saults JS, Morey CC, Mattox S, Hismjatullina A, Conway ARA. О способности внимания: ее оценка и роль в рабочей памяти и когнитивных способностях.Познан. Психол. 2005; 51:42–100. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Elliott EM, Saults JS, Nugent LD, Bomb P, Hismjatullina A. Переосмысление скоростных теорий когнитивного развития: увеличение скорости припоминания без ущерба для точности. Психол. науч. 2006а; 17:67–73. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Fristoe NM, Elliott EM, Brunner RP, Saults JS. Объем внимания, контроль внимания и интеллект у детей и взрослых. Мем. Познан.2006б; 34:1754–1768. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Morey CC. Как можно исследовать пределы удержания рабочей памяти при выполнении двух задач? Психол. науч. 2007; 18: 686–688. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Naveh-Benjamin M, Kilb A, Saults JS. Развитие зрительной рабочей памяти на протяжении всей жизни: когда привязка признаков затруднена? Дев. Психол. 2006с; 42:1089–1102. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Nugent LD, Elliott EM, Ponomarev I, Saults JS.Роль внимания в развитии кратковременной памяти: возрастные различия вербального объема восприятия. Детский Дев. 1999; 70: 1082–1097. [PubMed] [Google Scholar]
- Cowan N, Saults JS, Nugent LD. Роль абсолютного и относительного количества времени в забывании в непосредственной памяти: случай сравнения высоты тона. Психон. Бык. Ред. 1997а; 4:393–397. [Google Scholar]
- Коуэн Н., Вуд Н.Л., Ньюджент Л.Д., Трейсман М. В краткосрочной вербальной памяти есть два эффекта длины слова: противоположные эффекты длительности и сложности. Психол. науч. 1997b; 8: 290–295. [Google Scholar]
- Craik FIM, Birtwistle J. Проактивное торможение в свободном воспоминании. Дж. Эксп. Психол. 1971; 91: 120–123. [Google Scholar]
- Crowder RG. Упадок кратковременной памяти. Акта Психол. 1982; 50: 291–323. [PubMed] [Google Scholar]
- Crowder RG. Кратковременная память: где мы находимся? Мем. Познан. 1993; 21: 142–145. [PubMed] [Google Scholar]
- Данеман М., Карпентер П.А. Индивидуальные различия в рабочей памяти и чтении. J Вербальное обучение.Вербальное поведение. 1980; 19: 450–466. [Google Scholar]
- Daneman M, Merikle PM. Рабочая память и понимание языка: метаанализ. Психон. Бык. 1996; 3:422–433. [PubMed] [Google Scholar]
- Darwin CJ, Turvey MT, Crowder RG. Слуховой аналог процедуры частичного отчета Сперлинга: свидетельство кратковременного слухового запоминания. Познан. Психол. 1972; 3: 255–267. [Google Scholar]
- Davelaar EJ, Goshen-Gottstein Y, Ashkenazi A, Haarman HJ, Usher M. Новый взгляд на упадок кратковременной памяти: эмпирические и вычислительные исследования эффектов новизны.Психол. 2005; 112:3–42. [PubMed] [Google Scholar]
- Ebbinghaus H. Перевод Х.А. Ругер и К. Э. Буссениус. Нью-Йорк: Педагогический колледж Колумбийского университета; 1885–1913. Память: вклад в экспериментальную психологию. (Первоначально на немецком языке, Ueber das gedächtnis: Untersuchen zur Experimentellen Psychologie) [Google Scholar]
- Engle RW. Объем рабочей памяти как исполнительное внимание. Курс. Реж. Психол. науч. 2002; 11:19–23. [Google Scholar]
- Engle RW, Tuholski SW, Laughlin JE, Conway ARA.Рабочая память, кратковременная память и общий подвижный интеллект: латентно-вариативный подход. Дж. Эксп. Психол. Генерал 1999; 128: 309–331. [PubMed] [Google Scholar]
- Ericsson KA, Kintsch W. Долговременная рабочая память. Психол. 1995; 102:211–245. [PubMed] [Google Scholar]
- Friedman NP, Miyake A, Corley RP, Young SE, DeFries JC, Hewitt JK. Не все исполнительные функции связаны с интеллектом. Психол. науч. 2006; 17: 172–179. [PubMed] [Google Scholar]
- Gavens N, Barrouillet P.Задержки удержания, эффективности обработки и ресурсов внимания в развитии охвата рабочей памяти. Дж. Мем. Ланг. 2004; 51: 644–657. [Google Scholar]
- Гланцер М., Куниц А.Р. Два механизма хранения в свободном отзыве. J. Вербальное обучение. Вербальное поведение. 1966; 5: 351–360. [Google Scholar]
- Гленберг А.М., Суонсон, Северная Каролина. Теория временной различимости эффектов новизны и модальности. Дж. Эксп. Психол. Учиться. Мем. Познан. 1986; 12:3–15. [PubMed] [Google Scholar]
- Gold JM, Fuller RL, Robinson BM, McMahon RP, Braun EL, Luck SJ.Интактный контроль внимания за кодированием рабочей памяти при шизофрении. Дж. Ненормальный. Психол. 2006; 115: 658–673. [PubMed] [Google Scholar]
- Greene RL. Влияние дизайна эксперимента: пример парадигмы Брауна-Петерсона. Может. Дж. Эксп. Психол. 1996; 50: 240–242. [Google Scholar]
- Guttentag RE. Требование умственного усилия к кумулятивной репетиции: исследование развития. Дж. Эксп. Детская психология. 1984; 37: 92–106. [Google Scholar]
- Hebb DO. Организация поведения.Нью-Йорк: Уайли; 1949. [Google Scholar]
- Хоккей Р. Скорость представления в оперативной памяти и прямое манипулирование стратегиями обработки ввода. QJ Exp. Психол. А. 1973; 25: 104–111. [Google Scholar]
- Джеймс В. Принципы психологии. Нью-Йорк: Генри Холт; 1890. [Google Scholar]
- Кейн М.Дж., Блекли М.К., Конуэй А.Р., Энгл Р.В. Взгляд контролируемого внимания на объем рабочей памяти. Дж. Эксп. Психол. Ген. 2001; 130:169–183. [PubMed] [Google Scholar]
- Кейн М.Дж., Хэмбрик Д.З., Тухольски С.В., Вильгельм О., Пейн Т.В., Энгл Р.Э.Общая емкость рабочей памяти: латентно-вариативный подход к объему вербальной и зрительно-пространственной памяти и рассуждениям. Дж. Эксп. Психол. Ген. 2004; 133:189–217. [PubMed] [Google Scholar]
- Keppel G, Underwood BJ. Упреждающее торможение при кратковременном удержании отдельных элементов. J. Вербальное обучение. Вербальное поведение. 1962; 1: 153–161. [Google Scholar]
- Kyllonen PC, Christal RE. Способность к рассуждению — это (немного больше, чем) объем рабочей памяти? Интеллект. 1990; 14: 389–433. [Google Scholar]
- Лепин Р., Барруйе П., Камос В.Что делает объемы рабочей памяти столь предсказуемыми для высокого уровня познания? Психон. Бык. 2005; 12:165–170. [PubMed] [Google Scholar]
- Левандовски С., Дункан М., Браун GDA. Время не вызывает забывания при краткосрочном серийном воспоминании. Психон. Бык. 2004; 11:771–790. [PubMed] [Google Scholar]
- McGeoch JA. Забвение и закон неиспользования. Психол. 1932; 39: 352–370. [Google Scholar]
- Мелтон А.В. Значение кратковременной памяти для общей теории памяти.J. Вербальное обучение. Вербальное поведение. 1963; 2: 1–21. [Google Scholar]
- Миллер Г.А. Магическое число семь плюс-минус два: некоторые ограничения нашей способности обрабатывать информацию. Психол. 1956; 63:81–97. [PubMed] [Google Scholar]
- Миллер Г.А. Джордж А. Миллер. В: Линдзи Г., редактор. История психологии в автобиографии. Том. VIII. Стэнфорд, Калифорния: Издательство Стэнфордского университета; 1989. С. 391–418. [Google Scholar]
- Миллер Г.А., Галантер Э., Прибрам К.Х. Планы и структура поведения.Нью-Йорк: Холт, Райнхарт и Уинстон, Инк.; 1960. [Google Scholar]
- Мори К.С., Коуэн Н. Когда визуальная и вербальная память конкурируют: свидетельство междоменных ограничений в рабочей памяти. Психон. Бык. 2004; 11: 296–301. [PubMed] [Google Scholar]
- Мори К.С., Коуэн Н. Когда визуальные и вербальные воспоминания конфликтуют? Важность загрузки и извлечения рабочей памяти. Дж. Эксп. Психол. Учиться. Мем. Познан. 2005; 31: 703–713. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Nairne JS.Запоминание в краткосрочной перспективе: дело против стандартной модели. Анну. Преподобный Психолог. 2002; 53:53–81. [PubMed] [Google Scholar]
- Наве-Бенджамин М. , Коуэн Н., Килб А., Чен З. Возрастные различия в немедленном последовательном воспроизведении: формирование и емкость диссоциирующих фрагментов. Мем. Когнит. 2007; 35: 724–737. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Neath I, Surprenant A. Человеческая память. 2-е изд. Белмонт, Калифорния: Уодсворт; 2003. [Google Scholar]
- Nipher FE. О распределении ошибок в числах, записанных по памяти.Транс. акад. науч. Святой Луи. 1878;3:ccx–ccxi. [Google Scholar]
- Peterson LR, Peterson MJ. Кратковременное запоминание отдельных словесных единиц. Дж. Эксп. Психол. 1959; 58: 193–198. [PubMed] [Google Scholar]
- Солтс Дж. С., Коуэн Н. Центральный предел емкости для одновременного хранения зрительных и слуховых массивов в рабочей памяти. Дж. Эксп. Психол. 2007; 136: 663–684. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sperling G. Информация доступна в кратких визуальных презентациях.Психол. моногр. 1960;74 (Whole No. 498.) [Google Scholar]
- Stoltzfus ER, Hasher L, Zacks RT. Рабочая память и поиск: подход торможения-ресурса. В: Richardson JTE, Engle RW, Hasher L, Logie RH, Stoltzfus ER, Zacks RT, редакторы. Рабочая память и человеческое познание. Нью-Йорк: Издательство Оксфордского университета; 1996. С. 66–88. [Google Scholar]
- Talmi D, Grady CL, Goshen-Gottstein Y, Moscovitch M. Нейровизуализация кривой последовательного положения: тест моделей с одним магазином и с двумя магазинами.Психол. науч. 2005; 16: 716–723. [PubMed] [Google Scholar]
- Tulving E, Patkau JE. Одновременное влияние контекстуального ограничения и частоты слов на немедленное запоминание и изучение вербального материала. Может. Дж. Психол. 1962; 16: 83–95. [PubMed] [Google Scholar]
- Turvey MT, Brick P, Osborn J. Упреждающее вмешательство в кратковременную память как функция интервала сохранения предыдущего элемента. QJ Exp. Психол. 1970; 22: 142–147. [Google Scholar]
- Цзэн О.Дж.Л. Положительный эффект недавности в отсроченном свободном отзыве.J. Вербальное обучение. Вербальное поведение. 1973; 12: 436–439. [Google Scholar]
- Ансворт Н., Энгл Р.В. Характер индивидуальных различий в объеме рабочей памяти: активное сохранение в первичной памяти и контролируемый поиск во вторичной памяти. Психол. 2007; 114:104–132. [PubMed] [Google Scholar]
- Vogel EK, McCollough AW, Machizawa MG. Нейронные показатели выявляют индивидуальные различия в управлении доступом к рабочей памяти. Природа. 2005; 438: 500–503. [PubMed] [Google Scholar]
- Vogel EK, Woodman GF, Luck SJ.Динамика консолидации зрительной рабочей памяти. Дж. Эксп. Психол. Гум. Восприятие. Выполнять. 2006; 32:1436–1451. [PubMed] [Google Scholar]
- Waugh NC, Norman DA. Первичная память. Психол. 1965; 72:89–104. [PubMed] [Google Scholar]
- Wickelgren WA. Односледовая теория хрупкости динамики памяти. Мем. Познан. 1974; 2: 775–780. [PubMed] [Google Scholar]
 Вербальное поведение. 1975; 14: 575–589. [Google Scholar]
Вербальное поведение. 1975; 14: 575–589. [Google Scholar] , Уилкс А., редакторы. Исследования долговременной памяти. Оксфорд, Англия: Wiley; 1975. С. 3–18. [Google Scholar]
, Уилкс А., редакторы. Исследования долговременной памяти. Оксфорд, Англия: Wiley; 1975. С. 3–18. [Google Scholar]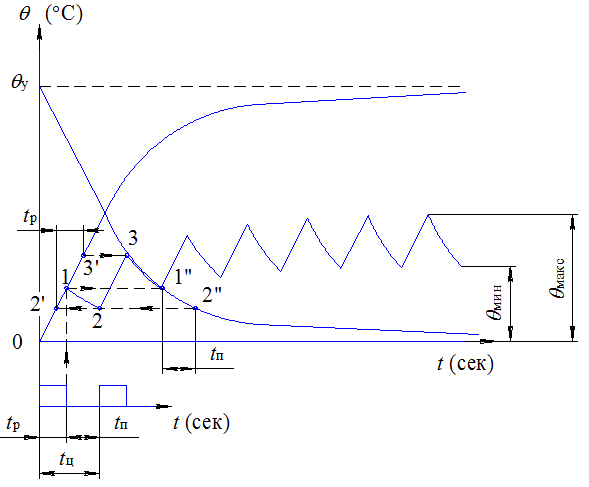 Мем. Познан. 2002; 30:1086–1095. [PubMed] [Google Scholar]
Мем. Познан. 2002; 30:1086–1095. [PubMed] [Google Scholar] Психол. Бык. 1988; 104: 163–191. [PubMed] [Google Scholar]
Психол. Бык. 1988; 104: 163–191. [PubMed] [Google Scholar] (в печати) [Бесплатная статья PMC] [PubMed] [Google Scholar]
(в печати) [Бесплатная статья PMC] [PubMed] [Google Scholar] Как можно исследовать пределы удержания рабочей памяти при выполнении двух задач? Психол. науч. 2007; 18: 686–688. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Как можно исследовать пределы удержания рабочей памяти при выполнении двух задач? Психол. науч. 2007; 18: 686–688. [Бесплатная статья PMC] [PubMed] [Google Scholar] Психол. науч. 1997b; 8: 290–295. [Google Scholar]
Психол. науч. 1997b; 8: 290–295. [Google Scholar] Новый взгляд на упадок кратковременной памяти: эмпирические и вычислительные исследования эффектов новизны.Психол. 2005; 112:3–42. [PubMed] [Google Scholar]
Новый взгляд на упадок кратковременной памяти: эмпирические и вычислительные исследования эффектов новизны.Психол. 2005; 112:3–42. [PubMed] [Google Scholar] Не все исполнительные функции связаны с интеллектом. Психол. науч. 2006; 17: 172–179. [PubMed] [Google Scholar]
Не все исполнительные функции связаны с интеллектом. Психол. науч. 2006; 17: 172–179. [PubMed] [Google Scholar]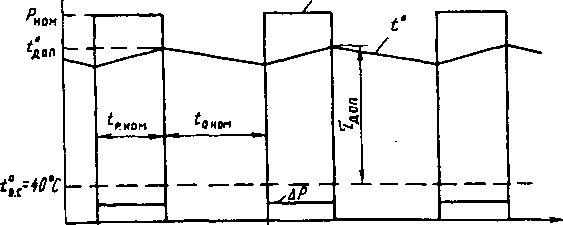 [Google Scholar]
[Google Scholar] Упреждающее торможение при кратковременном удержании отдельных элементов. J. Вербальное обучение. Вербальное поведение. 1962; 1: 153–161. [Google Scholar]
Упреждающее торможение при кратковременном удержании отдельных элементов. J. Вербальное обучение. Вербальное поведение. 1962; 1: 153–161. [Google Scholar] Психол. 1956; 63:81–97. [PubMed] [Google Scholar]
Психол. 1956; 63:81–97. [PubMed] [Google Scholar] , Коуэн Н., Килб А., Чен З. Возрастные различия в немедленном последовательном воспроизведении: формирование и емкость диссоциирующих фрагментов. Мем. Когнит. 2007; 35: 724–737. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Коуэн Н., Килб А., Чен З. Возрастные различия в немедленном последовательном воспроизведении: формирование и емкость диссоциирующих фрагментов. Мем. Когнит. 2007; 35: 724–737. [Бесплатная статья PMC] [PubMed] [Google Scholar] Рабочая память и поиск: подход торможения-ресурса. В: Richardson JTE, Engle RW, Hasher L, Logie RH, Stoltzfus ER, Zacks RT, редакторы. Рабочая память и человеческое познание. Нью-Йорк: Издательство Оксфордского университета; 1996. С. 66–88. [Google Scholar]
Рабочая память и поиск: подход торможения-ресурса. В: Richardson JTE, Engle RW, Hasher L, Logie RH, Stoltzfus ER, Zacks RT, редакторы. Рабочая память и человеческое познание. Нью-Йорк: Издательство Оксфордского университета; 1996. С. 66–88. [Google Scholar] Вербальное поведение. 1973; 12: 436–439. [Google Scholar]
Вербальное поведение. 1973; 12: 436–439. [Google Scholar]Повторное воздействие кратковременного поведенческого стресса устраняет ранее существовавшее депрессивно-подобное поведение, вызванное стрессом, у мышей
Животные
Семинедельные самцы мышей C57/BL6 были приобретены у Daehan BioLink (Ымсон, Чхунбук, Республика Корея). Восьминедельные самцы и самки мышей ICR (CD1) были приобретены у OrientBio. Inc. (Соннам, Кёнгидо, Республика Корея) и использовались в качестве селекционеров. Всех мышей содержали попарно в стандартных прозрачных пластиковых клетках в помещении с регулируемой температурой (23–24 °C) и влажностью (50–60%) с пищей и питьевой водой, доступными вволю. В помещении для животных поддерживался 12-часовой цикл свет/темнота (свет включался в 7 часов утра) в специальной свободной от патогенов среде. Эксперименты на животных проводились в соответствии с рекомендациями по уходу за животными Комитета по этике животных Женского университета Ихва, и все протоколы, использованные в настоящем исследовании, были одобрены Институциональным комитетом по уходу и использованию животных (IACUC) Женского университета Ихва (IACUC16-018). и IACUC19-015).
Восьминедельные самцы и самки мышей ICR (CD1) были приобретены у OrientBio. Inc. (Соннам, Кёнгидо, Республика Корея) и использовались в качестве селекционеров. Всех мышей содержали попарно в стандартных прозрачных пластиковых клетках в помещении с регулируемой температурой (23–24 °C) и влажностью (50–60%) с пищей и питьевой водой, доступными вволю. В помещении для животных поддерживался 12-часовой цикл свет/темнота (свет включался в 7 часов утра) в специальной свободной от патогенов среде. Эксперименты на животных проводились в соответствии с рекомендациями по уходу за животными Комитета по этике животных Женского университета Ихва, и все протоколы, использованные в настоящем исследовании, были одобрены Институциональным комитетом по уходу и использованию животных (IACUC) Женского университета Ихва (IACUC16-018). и IACUC19-015).
Кратковременный иммобилизационный стресс
Мышей по отдельности помещали в 50-мл полипропиленовую коническую пробирку с множеством вентиляционных отверстий и фиксировали в течение 5, 10, 15 мин или указанного времени, начиная с 10:00, и эту обработку повторяли в течение указанное количество дней. После каждого сеанса фиксации мышей возвращали в их домашние клетки.
Хронический иммобилизационный стресс (CRST)
CRST проводили, как описано ранее в ссылке. 60 . Мышей индивидуально ограничивали в хорошо вентилируемой конической пробирке объемом 50 мл в течение 2 ч ежедневно с 10:00 до 20:00.м. до 12 часов дня, и эта процедура повторялась в течение 14 дней. После каждого ежедневного сеанса иммобилизации мышей помещали в их домашние клетки со свободным доступом к пище и воде.
Хронический стресс социального поражения (CSDS)
Лечение CSDS и отбор восприимчивых к стрессу мышей проводили, как описано ранее в ссылке. 61 . Мыши C57/BL6 индивидуально подвергались воздействию нового агрессора ICR в течение 10 минут, чтобы вызвать физический стресс поражения, а затем содержались до конца дня в отсеке клетки агрессора ICR, разделенном перфорированной акриловой перегородкой.Эту процедуру повторяли в течение 10 дней с другим агрессором ICR каждый день. Агрессоры ICR были предварительно отобраны на основе следующих критериев: латентность к начальной атаке была менее 60 с, но выше 30 с, и любые агрессоры не проявляли чрезмерно агрессивных атак, таких как тяжелые ранения и кровотечение во время воздействия социального поражения, и если они это делали не отвечающие критериям отбора, они исключались из следующего эксперимента.
Чувствительные к стрессу и устойчивые мыши были отобраны на основе коэффициента социального взаимодействия отдельных животных, оцененного с помощью теста социального взаимодействия на 11-й день.Мышей, получавших CSDS, по отдельности помещали в открытое поле (45 см ×45 см × 40см), содержащее пустую перфорированную интерактивную коробку, расположенную с одной стороны открытого поля, и позволяли исследовать открытое поле в течение 2,5 мин. Регистрировалось время, затраченное на изучение пустого поля взаимодействия в поле. В то время как испытуемую мышь возвращали в ее домашнюю клетку на 1 мин, агрессора ICR помещали в коробку для взаимодействия, а затем испытуемой мыши давали возможность свободно исследовать открытое поле в течение 2,5 мин. Фиксировалось время взаимодействия с агрессором ИКР в поле взаимодействия в поле.Коэффициент социального взаимодействия (СИ) определяли как отношение общего времени нахождения в зоне взаимодействия с ИКР-агрессором к времени нахождения в зоне взаимодействия в отсутствие ИКР-агрессора. Когда отношение SI было ниже 1,0, мышей относили к группе восприимчивых, тогда как отношение SI было 1,0 и выше, мышей относили к группе устойчивых. Восприимчивых к CSDS мышей использовали для дальнейшего исследования.
Материнский стресс (МС) и постнатальный стресс
Самцов и самок мышей ICR в возрасте 8–9 недель скрещивали, и беременных самок случайным образом распределяли в группу материнского стресса (МС) или нормальную (N) контрольную группу.Беременных самок с РС ежедневно лечили 2-часовым ограничением свободы в течение 2 часов (10:00–12:00) с 8,5 дня после полового акта (dpc) до родов (в 19,5–20,5 dpc). Нормальных (N) беременных самок содержали без процедуры стресса параллельно с самками MS. Потомство обеих групп отнимали от груди на 20-й день после рождения (PN20) и выращивали парами с представителями одного пола в клетках в стандартных условиях. Для групп мышей, назначенных для лечения CRST, в PN49 потомство от N контрольных матерей или матерей с MS помещали отдельно в однополые пары из разных пометов, чтобы предотвратить возможные эффекты помета.Затем мышей N и MS подвергали ежедневному 2-часовому иммобилизации в течение 14 дней (во время PN50–PN63). После этого половину мышей, получавших CRST, подвергали режиму RS5, а оставшуюся половину использовали в качестве контрольной группы CRST.
Иммуногистохимия
Иммуногистохимия проводилась, как описано ранее 38,39 . Мышей перфузировали 4% параформальдегидом транскардиальным методом, а изолированный мозг постфиксировали при 4 °C в течение ночи. Каждый мозг был коронарно срезы толщиной 40 мкм с использованием вибратома (VT1000S, Leica Instruments, Nussloch, Германия).Собранные срезы блокировали на 1 час 4% бычьим сывороточным альбумином в фосфатно-солевом буфере (PBS), содержащем 0,1% Triton X-100, а затем инкубировали с первичными антителами при 4°C в течение ночи. После промывки срезы реагировали в течение 90 мин с биотинилированными вторичными антителами: анти-кроличьим IgG (BA-1000, Vector Laboratories, Burlingame, CA, USA) или анти-мышиным IgG (BA-9200, Vector Laboratories) в разведении 1:200. в ПБСТ. Сигналы визуализировали с помощью набора ABC Elite (PK-6200, Vector Laboratories).
Для анализа экспрессии c-Fos , индуцированной кратковременным иммобилизирующим стрессом, мышам CRST на 8-й день постстрессорного периода вводили однократное 5-минутное ограничение или 15-минутное ограничение (S5 × 1d и S15 × 1d соответственно) и умерщвлены через 20 и 10 минут соответственно. Другие группы мышей CRST подвергались ежедневному 5-минутному или 15-минутному сдерживанию в течение 7 дней и умерщвлялись через 20 и 10 минут после последнего 5-минутного или 15-минутного сдерживания, соответственно, на 8-й день после стресса (S5). × 8d и S15 × 8d соответственно).
Для анализа экспрессии c-Fos , индуцированной инъекцией CORT в низкой дозе, мышам CRST вводили инъекции CORT в дозах 0,1, 0,5 или 1,0 мг кг кг -1 в день в течение 7 дней и умерщвляли через 25 мин после инъекции. дополнительная инъекция CORT на 8-й день после стресса. Саннивейл, Калифорния, США), а также с использованием шестибалльной оценочной шкалы, как описано ранее 62 .Уровень экспрессии c-Fos оценивали по числовой шкале от 0 до 5; 0 соответствует 0–50 c-Fos -положительных клеток/мм 2 в области подсчета; +1 для 51–100 c-Fos -положительных клеток/мм 2 ; +2 для 101–200 c-Fos -положительных клеток/мм 2 ; +3 для 201–350 c-Fos -положительных клеток/мм 2 ; +4 для 351–600 c-Fos -положительных клеток/мм 2 ; и +5 для >600 c-Fos -положительных клеток/мм 2 .Среднее значение экспрессии c-Fos в каждой области было округлено до ближайшей десятой.
В качестве первичных антител использовали анти- c-Fos (sc-52, 1:300 для иммунофлуоресценции, 1:2000 для иммуногистохимии; sc-271243, 1:300, Santa Cruz Biotechnology, CA, USA), анти- Glu4 (G9282, Sigma-Aldrich, Миссури, США), анти-GAD67 (MAB5406, EMD Millipore, CA, США), анти-GR (sc-136209, 1:500, Santa Cruz Biotechnology; 12041 S, 1:500, Cell Signaling Technology, Массачусетс, США), анти-Fkpb5 (GTX84491, 1:50, Genetex, Калифорния, США), анти-NR1 (sc-1467, 1:50, Santa Cruz Biotechnology), анти-NR2A (sc-9056 , 1:50, Santa Cruz Biotechnology), анти-NR2B (ab93610, 1:100, Abcam, Cambridge, UK), анти-p-CaMKIIα (Thr286) (sc-12886, 1:1000, Santa Cruz Biotechnology) и анти-p-ERK1/2 (Thr202/Tyr204) (4370 s, 1:500, Cell Signaling Technology).Для иммунофлуоресцентного окрашивания использовали следующие флуоресцентно-меченые вторичные антитела: антикроличьи IgG DyLight488 (DI-1488; 1:500, Vector Laboratories), антикроличьи IgG DyLight594 (DI-1094; 1:500, Vector Laboratories), антимышиный IgG DyLight488 (DI-2488; 1:500, Vector Laboratories), антимышиный IgG DyLight594 (DI-2594; 1:500, Vector Laboratories) и антикозий IgG-FITC (sc-2356, 1: 200, Санта-Крус-Биотехнология). Окрашенные срезы монтировали с помощью монтажного раствора против выцветания, содержащего DAPI (H-1200; Vector Laboratories).Окрашенные иммунофлуоресцентные изображения анализировали с использованием микроскопа Olympus BX 51, оснащенного камерой DP71 и программой MetaMorph (Molecular Devices).
ПЦР-анализ в реальном времени
ПЦР в реальном времени проводили, как описано ранее 38,39 . Тотальную РНК очищали из ткани головного мозга с использованием реагента TRIzol (15596018, Invitrogen Life Technologies, Карлсбад, Калифорния, США) и обрабатывали ДНКазой I (M610A, Promega, Миссури, США) для устранения загрязнения геномной ДНК. Обратную транскрипцию суммарной РНК (2 мкг) проводили в объеме 20 мкл с использованием системы обратной транскрипции (A3500, Promega, Миссури, США).ПЦР в реальном времени проводили с использованием 10 мкл 2X iQ™ SYBR Green Supermix (№ 170882, Bio-Rad Laboratories, Фостер-Сити, Калифорния, США), 1 мкл каждого из 5 пмоль/мкл прямого и обратного праймеров и 4 мкл комплементарную ДНК (кДНК) (разведение 1/8 конверсии) в общем объеме 20 мкл с использованием детектора системы ПЦР в реальном времени CFX 96 (Bio-Rad Laboratories). Для анализа данных количественной ПЦР использовали Bio-Rad CFX Manager 3.1. Наборы праймеров, использованные в ПЦР, перечислены в дополнительных данных 5.Мышей анестезировали внутрибрюшинной инъекцией (300 мкл на мышь) 2,5% авертина (2,2,2-трибромэтанол; 300 мг кг -1) (T48402, Sigma-Aldrich) в трет-амиловом спирте (24048-6, Sigma-Aldrich), разбавленный до 2,5% авертина физиологическим раствором. Кровь собирали из брюшной аорты умерщвленных мышей утром (8:00–12:00), а сыворотку получали центрифугированием при 3000× g и 4 °C в течение 15 мин, а затем хранили при –80 °C до использования. . Каждый разведенный образец сыворотки (10 мкл) смешивали на 96-луночном планшете с равным объемом раствора реагента для вытеснения стероидов и сыворотки, разбавленной в соотношении 1:40 или 1:80 в буфере для анализа ELISA, входящем в набор для ELISA с кортикостероном (ADI -901-097, Enzo Life Sciences, Нью-Йорк, США).Реакцию инкубировали в течение 2 ч при комнатной температуре на орбитальном шейкере, вращающемся со скоростью 120 об/мин. Реакционную смесь из каждой лунки удаляли, а планшет промывали промывочным буфером. Раствор субстрата pNpp, входящий в комплект, добавляли по 200 мкл на лунку, и планшет инкубировали в течение 1 ч при комнатной температуре без встряхивания. Наконец, поглощение при 405 нм измеряли с помощью спектрофлуориметра (SpectraMax ® M5, Molecular Devices, Саннивейл, Калифорния, США) и анализировали с использованием программного обеспечения SoftMax Pro версии 5.4. Уровни кортикостерона анализировали с использованием онлайн-инструмента анализа данных для ELISA компании Enzo Life Science (http://www.myassays.com).
Хемогенетическая модуляция активности определенных нейронов
Хемогенетическая манипуляция нейронов проводилась с использованием системы DREADD, как описано ранее 63 . Мышей анестезировали смесью кетамина и ксилазина (3,5:1). Для активации PL-нейронов использовали вирусный вектор hM3D, управляемый промотором CaMKIIα, AAV8-CaMKIIα-hM3D(Gq)-mCherry (3 × 10 10 вирусных частиц, мл -1 ) или вирусный вектор hM3D, управляемый промотором hSyn, AAV8-hSyn. -hM3D(Gq)-mCherry (4.3 × 10 10 вирусных частиц мл -1 ) билатерально вводили в PL с использованием системы стереотаксической инъекции (Vernier Stereotaxic Instrument, Leica Biosystems, Wetzlar, Germany) и шприца Hamilton с иглой 30-го калибра. Для ингибирования нейронов PL вирусный вектор hM4D, управляемый промотором CaMKIIα, AAV8-CaMKIIα-hM4D(Gi)-mCherry (1,6 × 10 10 вирусных частиц мл -1 ) билатерально инъецировали в PL. Через 7 дней восстановления мышей подвергали воздействию CRST. Мышам CRST вводили носитель или N-оксид клозапина (CNO) (0.1, 1 или 3 мг кг −1 ) за 30 мин до лечения с 5-минутным ограничением по схеме эксперимента.
Для трансдукции hM4D(Gi) в цепи PL→dBNST вирусный вектор AAV8-hSyn-DIO-hM4D(Gi)-mCherry (3 × 10 10 вирусных частиц мл −1 ) билатерально инъецировали в PL (AP, +2,0; ML, ±0,23; DV, -2,0 мм) и ретроградный вектор Cre AAV8-EF1α-mCherry-IRES-WGA-Cre (1,5 × 10 10 вирусных частиц мл -1 ) был введен в dBNST (AP, +0.25; МЛ, ±1,15; DV, −4,2 мм), как показано в плане эксперимента. Для хемогенетического ингибирования нейронов PL→dBNST им вводили носитель или CNO (3 мг кг -1 , внутрибрюшинно) за 30 мин до обработки с 5-минутным ограничением, как показано в экспериментальной схеме. Объем инъекции и скорость инъекции для каждого целевого участка были следующими: для ФЛ — 0,5 мкл со скоростью 0,25 мкл в минуту; для dBNST — 0,3 мкл со скоростью 0,15 мкл в мин. Иглу для инъекции оставляли на месте на 5 минут, чтобы обеспечить адекватную доставку, и медленно извлекали в течение 5 минут.
Для трансдукции hM4D(Gi) в цепи PL→NAcc или PL→BLA использовали вирусный вектор AAV8-hSyn-DIO-hM4D(Gi)-mCherry (3 × 10 10 вирусных частиц мл -1 ) билатерально в ЛП (AP, +2,0; ML, ±0,23; DV, -2,0 мм), и ретроградный вектор Cre AAV8-EF1α-mCherry-IRES-WGA-Cre (1,5 × 10 10 вирусных частиц мл -1 ) вводили в NAc (AP, +1,65; ML, ±1,15; DV, -5,15 мм) или BLA (AP, -1,3; ML, ±3,35; DV, -5,2 мм), как показано в экспериментальной дизайн.
Получены AAV8-CaMKIIα-hM3D(Gq)-mCherry, AAV8-CaMKIIα-hM4D(Gi)-mCherry, AAV8-hSyn-hM3D(Gq)-mCherry и AAV8-hSyn-DIO-hM4D(Gi)-mCherry. от доктора Брайана Рота (Addgene: плазмиды № 50476, № 50477, № 50474 и № 44362 соответственно). AAV8-EF1α-mCherry-IRES-WGA-Cre был приобретен у UNC Vector Core (Университет Северной Каролины, Северная Каролина, США). Вирусные векторы делили на аликвоты по прибытии и хранили при -80°С до использования. Каждый вирусный вектор разводили в 1x PBS (13,7 мМ NaCl, 0,27 мМ KCl, 0,8 мМ Na 2 HPO4, 0.2 мМ KH 2 PO 4 , pH 7,4).
Картирование нейронных цепей с помощью антеро- и ретроградных индикаторов
Мышей анестезировали кетамином и ксилазином (3,5:1). Чтобы пометить антероградные проекции нейронов PL, AAV2/1-CaMKIIα-EYFP (1,0 × 10 9 вирусных частиц мл -1 ) (AV-1-PV1917, Пенсильванский университет, Пенсильвания, США) стереотаксически вводили в PL (AP, +2,0; ML, -0,23; DV, -2,0 мм) объемом 0,5 мкл со скоростью 0,25 мкл/мин с использованием стереотаксической инъекционной системы (Vernier Stereotaxic Instrument) и иглы 30G.Через две недели после стереотаксической инъекции мозг готовили после перфузии 4% параформальдегидом транскардиальным методом. Чтобы пометить нейронные афференты к dBNST, субъединицу B (CTB) холерного токсина, конъюгированную с Alexa Fluor 488 (0,1% веса на объем) (C34755, Molecular Probes, Юджин, Орегон, США), ретроградный индикатор, стереотаксически вводили в мозг. dBNST (AP, +0,25; ML, -1,15; DV, -4,2 мм) в объеме 0,3 мкл со скоростью 0,15 мкл в мин с помощью иглы 30G. Через неделю после стереотаксической инъекции мозг готовили после перфузии 4% параформальдегидом, как описано выше.С помощью вибратома (Leica VT 1000 S; Leica Instruments, Nussloch, Германия) мозг был срезан в коронарной части толщиной 40 мкм. Флуоресцентные изображения собирали и анализировали с помощью микроскопа Olympus BX 51, оснащенного флуоресцентным осветителем X-cite 120 (EXFO Life Science & Industrial Division, Онтарио, Канада), камерой DP71 (Olympus) и программным обеспечением MetaMorph для автоматизации микроскопии и анализа изображений ( Molecular Devices, Саннивейл, Калифорния, США).
Оптогенетические манипуляции пути PL-to-NAcc
Оптогенетические манипуляции были выполнены, как описано ранее 64 .Вирусный вектор AAV2/1-CaMKIIα-eNpHR-EYFP (1,15 × 10 9 вирусных частиц мл −1 ) или AAV2/1-CaMKIIα-EYFP (1 × 10 9 вирусных частиц мл −1 ) стереотаксически вводили в ЛП (АД +2,0; ML +0,23; DV -2,0 мм) справа в объеме 0,5 мкл со скоростью 0,25 мкл в мин с помощью стереотаксической инъекционной системы (Vernier Stereotaxic Instrument ) и иглой 30G. Через 7 дней восстановления в NAc имплантировали оптическое волокно (диаметр 200 мкм, длина 5,15 мм; B280-2508-5, Doric Lenses, Квебек, Канада) (AP, +1.65; МЛ, +1,15; DV, -5,15 мм) справа у нормальных мышей. Для оптического ингибирования нейронов PL-NAcc имплантированное оптическое волокно было подключено к 532-нм твердотельному (DPSS) желтому лазеру с диодной накачкой (GL532T3-100; Shanghai Laser & Optics Century, Шанхай, Китай) через волоконно-оптическая канюля с адаптером FC/PC (D202-2027-5; линзы Doric). В поведенческих тестах наконечник волоконно-оптической канюли был соединен с патч-кордом с помощью разрезной втулки из диоксида циркония (F210-3002, линзы Doric) и оптоволоконного поворотного соединения (B300-0010, линзы Doric).Интенсивность желтого света была установлена на уровне 4 мВт. Импульсы желтого света генерировались с помощью стимулятора (#33521 A, Agilent Technologies, Санта-Клара, Калифорния, США) и подавались с частотой 20 Гц с шириной импульса 30 мс с временным интервалом (3 минуты для теста на общительность и 2 минуты). -мин для TST или FST). Интенсивность света через оптическое волокно подтверждали с помощью детектора света (PM200; Thorlabs, Ньютон, штат Нью-Джерси, США), оснащенного датчиком света (S121C, Thorlabs), перед началом поведенческого теста.
Вирусные векторы AAV AAV2/1-CaMKIIα-eNpHR-EYFP (AV-1-26971P) и AAV2/1-CaMKIIα-EYFP (AV-1-PV1917) были приобретены у Penn Vector Core (Университет Пенсильвании, Пенсильвания, США).
Нокдаун гена с помощью малой интерферирующей РНК (миРНК)
Стереотаксическую инъекцию миРНК проводили, как описано ранее 38,64 . Мышей анестезировали смесью гидрохлорида кетамина (50 мг мл -1) и гидрохлорида ксилазина (23,3мг мл-1) (3,5:1) в дозе 2,5мкл г-1 массы тела. Один объем разбавленной (50 нг мкл -1 ) миРНК смешивали с 2,5 объемами реагента для трансфекции Neurofect (T800075, Genlatis, Сан-Диего, Калифорния, США) и 0.5 объемов 50% сахарозы и инкубируют в течение 20 мин перед инъекцией в головной мозг. Всего 0,5 мкл смеси, содержащей 5,94 нг siRNA-GR, siRNA-CaMKIIα, siRNA-ERK1, siRNA-ERK2 или siRNA-контроль, стереотаксически инъецировали в каждую сторону PL (AP, +2,0; ML, ± 0,23; DV, -2,0 мм) со скоростью 0,25 мкл в минуту с использованием стереотаксической инъекционной системы (Vernier Stereotaxic Instrument) и иглы 30G. Ткани головного мозга готовили для анализа нокдауна или изменений экспрессии генов через 48–72 ч после инъекции миРНК.Поведенческие тесты проводили между 48 и 72 часами после инъекции миРНК.
Используемые siРНК: siRNA-control (SN-1012), siRNA-CaMKIIα (#12322, NM_177407.2, NM_009792.1), siRNA-MAPK3 (ERK1) (#26417, NM_011952.2), siRNA-MAPK1( ERK2) (#26413, NM_011949.3, NM_001038663.1) или siNr3c1(GR) (#14815, NM_008173.3), все они были приобретены у Bioneer Co. (Тэджон, Корея).
Вливание лекарств через канюлю
Имплантацию канюли и введение лекарства проводили, как описано ранее 64 .Мышей анестезировали кетамином и ксилазином (3,5:1). Направляющую канюлю 26-го калибра (C315G/SPC, Plastics One, Bilaney, UK) стереотаксически имплантировали в PL (AP, +2,0; ML, +0,23; DV, -2,0 мм). Канюлю фиксировали путем нанесения стоматологического клея Super-Bond C&B™ (Sun Medical, Морияма, Япония) на поверхность черепа мыши вокруг основания канюли. Через 15 мин для фиксации канюли к поверхности черепа наносили стоматологический цемент (Vertex-Dental, AA Zeist, Нидерланды), а затем герметизировали фиктивной канюлей (C313DC, Plastics One).У мышей было более 7 дней восстановления. В то время как мышей анестезировали 1,4% изофлураном в газовой смеси 70% закиси азота и 30% кислорода, препараты вводили в ЛС в объеме 0,5 мкл через внутреннюю канюлю 33-го калибра (C315I, Plastics One). KN62 (2,5 нмоль/инъекция; I2142, Sigma-Aldrich) вводили в ЛП в объеме 0,5 мкл со скоростью 0,25 мкл в минуту посредством канюляции.
Введение лекарств
Имипрамин (I0899, Sigma-Aldrich), кортикостерон (C2505, Sigma-Aldrich), гидрохлорид NBI27914 (1591, Tocris Bioscience), RU486 (M8046, Sigma-Aldrich) и N-оксид клозапина (CNO) дигидрохлорида (6329, Tocris Bioscience) разводили в 0.9% физиологический раствор и вводят внутрибрюшинно в объеме 120 мкл.
Вестерн-блот-анализ
Вестерн-блот-анализ проводили, как описано ранее 38 . Ткани PL от трех до четырех животных гомогенизировали в буфере RIPA (50 мМ Tris-HCl, pH 7,4, 150 мМ NaCl, 0,5 % Nonidet P-40, 0,25 % дезоксихолата натрия, 1 мМ EDTA и 0,5 % Triton X-100). содержащий коктейль ингибиторов протеазы (11836153001, Roche, Mannheim, Germany). Гомогенаты тканей, загруженные 20 мкг белка на дорожку, разделяли на SDS-PAGE и переносили на PVDF (1620177, Bio-Rad Laboratories).Мембраны инкубировали с блокирующим раствором (5% снятое молоко; 232100, Difco, MI, USA) в солевом растворе на основе Трис-буфера, содержащем Твин 20 (TBST; 150 мМ NaCl, 50 мМ Трис-HCl, рН 7,4, 0,1 % Твин 20). ) с последующим разведением первичных антител в 1% обезжиренном молоке при 4 °C в течение ночи. После промывки 1x TBST шесть раз по 10 минут каждый раз мембраны инкубировали со вторичным антителом в течение 1 часа. Используемые вторичные антитела представляли собой антикроличьи IgG-HRP (sc-2004, Santa Cruz Biotechnology) и антимышиные IgG-HRP (sc-2005, Santa Cruz Biotechnology), разведенные 1:2000 в 1% обезжиренном молоке.После промывки специфические сигналы на пятнах визуализировали с использованием набора для улучшенной хемилюминесценции (ELPIS-Biotech, Тэджон, Корея). Изображения вестерн-блоттинга количественно определяли с использованием программного обеспечения для анализа Image J v1.51k (NIH Image, Bethesda, MD, USA).
Первичными антителами были анти-p-CaMKIIα (Thr286) (sc-12886, 1:2000, Santa Cruz Biotechnology), CaMKIIα (A-1) (sc-13141, 1:2000, Santa Cruz Biotechnology), анти- p-ERK1/2 (Thr202/Tyr204) (4370 s, 1:500, Cell Signaling Technology), анти-ERK1/2 (MK1) (sc-135900, 1:2000, Santa Cruz Biotechnology) и анти-β- актин (C4) (sc-47778, 1:3000, Santa Cruz Biotechnology).
Адреналэктомия
Адреналэктомия (ADX) была выполнена, как описано ранее 65 . Мышей анестезировали смесью кетамина и ксилола (3,5:1). Хирургические инструменты предварительно стерилизовали бромсептом (DAESUNG Microbiological Labs. Co., Kyonggi, Республика Корея). Мышей помещали ничком на хирургическую кровать. Спинную сторону тела покрывали 10% раствором повидон-йода (№ 261, Сунгкванг, Чунгнам, Республика Корея). Левая сторона спины была захвачена щипцами (№ 5, Fine Science Tools Inc., Северный Ванкувер, Британская Колумбия Canada), а кожу и брюшину последовательно разрезали микродиссекционными ножницами (S3271, Sigma-Aldrich). Затем надпочечник захватили щипцами (#5, Fine Science Tools Inc.) и отрезали лезвием скальпеля (S2646, Sigma-Aldrich), установленным на рукоятке скальпеля (S2896, Sigma-Aldrich). Брюшину и кожу последовательно сшивали шелковой нитью 4/0-18 мм (SK434, Ailee Company Limited, Пусан, Республика Корея). Правосторонняя операция проводилась по той же схеме, что и левосторонняя.Адреналэктомированным мышам впоследствии вместо питьевой воды давали 0,9% физиологический раствор, но им не давали заместительную терапию кортикостероном. Мышей с ложнооперированным оперированием подвергали такой же хирургической процедуре: надпочечники захватывали щипцами, но не удаляли. После 7 дней восстановления половину мышей с ADX и ложнооперированных мышей подвергали лечению RS5, а оставшиеся мыши служили контролем.
Микроматричный анализ
Микрочиповый анализ проводили, как описано ранее 38 .Были подготовлены два независимых набора групп контроля (CON), CRST и CRST + RS5. Мышей подвергали обработке CRST с последующим введением RS5, а затем умерщвляли на 22-й день после стресса. Тотальную РНК экстрагировали из объединенной ткани PL 6–7 животных в каждой группе с использованием Trizol (Invitrogen Life Technologies) с последующей очисткой с использованием колонок RNeasy. (Киаген, Валенсия, США). Очищенная РНК имела отношение A260/280 1,9–2,1, определенное с помощью спектрофотометра ND-1000 (NanoDrop, Уилмингтон, Делавэр, США) и биоанализатора Agilent 2100 (Agilent Technologies, Пало-Альто, Калифорния, США).
Суммарная РНК (550 нг для каждой) была преобразована в кДНК первой и второй цепи путем транскрипции in vitro. Затем кДНК использовали для синтеза образцов кДНК, меченных биотином. Очищенные образцы кДНК, меченные биотином, фрагментировали путем нагревания их до 94°С в течение 35 мин в буфере для фрагментации матрицы. Каждый из фрагментированных и меченных биотином образцов кДНК (750 нг) гибридизовали с массивом гранул мыши Ref8 Expression v.2 (Illumina, Inc., Сан-Диего, США) в течение 16–18 ч при 58°C, согласно инструкции производителя. инструкции.После промывки сигналы матрицы амплифицируют стрептавидином Amersham Fluorolink – Cy3 (GE Healthcare Bio-Sciences, Little Chalfont, UK) и сканируют с помощью Illumina Bead Array Reader, конфокального сканера, в соответствии с инструкциями производителя.
Качество гибридизации и общая производительность массива чипов, включая внутренние датчики контроля качества и отсканированные необработанные данные, извлеченные с помощью программного обеспечения Illumina GenomeStudio v2011.1 (Gene Expression Module v1.9.0) контролировались компанией Macrogen Inc. (Сеул, Южная Корея). Сигналы микрочипов были преобразованы в значения шкалы log2 и нормализованы методом квантилей.
Профили экспрессии генов были проанализированы с использованием Multiexperiment Viewer (MeV), версия 4.9 (http://mev.tm4.org/). Были отобраны гены, экспрессия которых повышалась или понижалась в ≥1,2 раза с помощью CRST (CON по сравнению с CRST), и гены изменялись в ≥1,2 раза после обработки RS5 мышей, получавших CRST (CRST по сравнению с CRST + RS5). Профили экспрессии 264 CRST-чувствительных генов были представлены от самого высокого до самого низкого значения на тепловой карте с последующим неконтролируемым выравниванием с соответствующими генами из CRST по сравнению с соответствующими генами.Сравнение CRST + RS5. Независимо, профили экспрессии 722 RS5-чувствительных генов были выровнены, как указано выше, на тепловой карте, а затем наложены на соответствующие гены из сравнения CON и CRST.
Анализ обогащения Gene Ontology (GO)
Анализ обогащения GO был проведен с использованием базы данных STRING (Инструмент поиска для поиска взаимодействующих генов/белков) версии 11 32 (доступно по адресу: http://string-db.org/ ). Из общего числа 896 (264 + 722) генов, отобранных, как описано в предыдущем разделе, 860 генов были отнесены к функциональным группам с терминами биологического процесса (BP), тогда как 36 генов с неопределенной идентичностью были исключены из дальнейшего анализа.Иерархия терминов GO была назначена на основе базы данных генома мыши 66 (http://www.informatics.jax.org).
Серийный номер K означает, что кластеризация использовалась для группировки 860 генов в несколько функциональных кластеров. Каждое увеличение значения K добавляло новый кластер, который содержал элементы, в основном поставляемые из самого большого кластера (кластер 1). Когда >75% генов в кластере оставались в виде кластера в следующем последовательном кластерировании, эти два интеркластера считались принадлежащими к одной и той же функциональной группе и поэтому помечались одним и тем же цветовым кодом.Уровень удержания большинства родственных интеркластеров был выше 85% при группировании в 5–10 кластеров ( k = 5–10), тогда как уровень удержания снижался после группирования в 11 или более кластеров.
Сеть функционального белок-белкового взаимодействия (PPI) (обогащение PPI p значение , <1,0e-16), состоящая из восьми кластеров ( k = 8), была выбрана в качестве репрезентативной группы: Кластер 1 содержал большинство членов (622 гена). Однако члены кластера 1 были слабо интерактивны и разбросаны вне функциональных сетей, образованных остальными семью кластерами, включающими в общей сложности 238 генов.Поэтому кластер 1 был исключен из дальнейшего анализа. Кластеры со 2 по 8 содержали 71, 39, 29, 28, 27, 25 и 19 генов соответственно. Кластеры 5 и 7, содержащие 28 и 25 генов соответственно, охватывали термины ГО «ответ на стресс» и «ответ на глюкокортикоиды», наряду с клеточной коммуникацией, установлением локализации и реакцией на эндогенные стимулы.
Потенциальные взаимодействующие вещества, которые были пропущены при скрининге микрочипов, но которые могли существовать в функциональных сетях PPI, были найдены в базе данных сети STRING.Используя краевую достоверность 0,7 и выше для классификации узлов в сетях PPI, эти усилия привели к появлению 16 новых взаимодействующих. После этого были построены функциональные сети, включающие членов кластеров 5 и 7 и 16 новых членов. Узлы и ребра для взаимодействия с членами были закодированы цветом с помощью Cytoscape StringApp v3.7.2 67 .
ПЦР в реальном времени использовали для изучения профилей экспрессии генов Nr3c1(GR), Nr3c2(MR), Fkbp5, Fkbp4, Hsp90aa1 и Hsp90ab1, произвольно выбранных из кластера 7, а также экспрессии Dusp1, CaMKIIα , Mapk3(ERK1) и Mapk1(ERK2), выбранные из кластера 5 для указанных групп.
Поведенческие оценки
Поведенческие тесты проводились, как описано ранее 38,39 . Все поведенческие тесты записывались с помощью компьютеризированной системы видеослежения (SMART; Panlab S.I., Барселона, Испания) или системы записи с веб-камеры (HD Webcam #C210; Logitech, США). Все поведенческие тесты среди групп и внутри групп проводились рандомизированным образом или попеременно в зависимости от порядка тестирования и положения в испытательном оборудовании или на тестовом поле (т.г., слева и справа, между позициями в тестовом поле). Стойкие поведенческие изменения, вызванные CRST, и эффекты RS5 были проверены вслепую двумя экспериментаторами. Всех животных содержали парами, если не указано иное. За день до поведенческих тестов смена постельных принадлежностей не производилась. Комната для поведенческого тестирования освещалась непрямым освещением 20 люкс для теста на общительность и 250 люкс для TST и FST. Мышам давали 20–30 минут для адаптации в комнате для тестирования поведения перед началом каждого поведенческого теста.Фоновый звук в комнате для тестирования был замаскирован белым шумом мощностью 65 дБ на протяжении всех поведенческих тестов. Все поведенческие тесты проводились с 9:00 до 15:00. во время светового цикла. После каждого поведенческого теста все части аппарата очищали 70%-ным этанолом.
Двухкамерный тест социального взаимодействия
Двухкамерный тест социального взаимодействия (ТСВ) был разработан как модифицированная версия трехкамерного теста 68 и U-образного полевого теста с двумя вариантами ответов 38,39 .Двухкамерный аппарат состоял из двух симметричных камер (26 см × 26 см прямоугольного пола со стенками 40 см), разделенных промежуточным проходом (8 см шириной × 10 см длиной × 26 см высотой), с пустой круглой решетчатой клеткой (12). см в диаметре × 33 см в высоту) в каждой камере. Испытуемую мышь помещали на средний путь и давали ей свободно исследовать две камеры в течение 5 минут в качестве привыкания. В то время как испытуемую мышь возвращали в ее домашнюю клетку на 2 мин, социальную мишень (C57BL/6) помещали в одну из сетчатых клеток.Как только социальная мишень стабилизировалась, испытуемую мышь помещали в середину двух камер выбора, с социальной мишенью с одной стороны и пустой клеткой с другой стороны, и позволяли свободно исследовать камеры в течение 5 мин. . Затраченное время и траектория, пройденная в каждой камере, записывались. Сторона, содержащая социальную цель, называлась целевым полем, а противоположная сторона называлась нецелевым полем. Подробные процедуры подготовки испытуемых животных, привыкания, социальных целей, тестов на общительность и социальное взаимодействие, а также обращение с комнатой для тестирования поведения были выполнены, как описано в предыдущем отчете 38,39 .
U-образный полевой тест на общительность с двумя вариантами ответов
U-образный полевой тест на общительность с двумя вариантами ответов был проведен, как описано ранее 38,39 . U-образное поле с двумя вариантами ответов состояло из открытого поля (45 см × 45 см), частично разделенного стеной (20 см в ширину × 35 см в высоту) до центральной точки для получения двух симметричных прямоугольных полей. Круглая сетчатая клетка (диаметром 12 см × высотой 33 см) была размещена на каждой стороне стены. Для привыкания испытуемой мыши давали свободно исследовать U-образное поле, содержащее пустую решетчатую клетку с каждой стороны, в течение 5 минут.В то время как испытуемую мышь возвращали в ее домашнюю клетку на 2 минуты, социальную мишень загружали в одну из клеток сетки. После этого испытуемой мыши снова давали исследовать U-образное поле в течение 5 минут, и регистрировали затраченное время и выбранную траекторию. Сторона, содержащая социальную цель, называлась целевым полем, а противоположная сторона называлась нецелевым полем.
Тест предпочтения сахарозы
Тест предпочтения сахарозы (SPT) проводили, как описано ранее 69 .Мышей приучали к двум бутылкам с водой в течение 7 дней с последующим предъявлением двух бутылок с 1% раствором сахарозы на 24 часа. Затем мышей лишали воды и раствора сахарозы на 9 часов (с 10 до 19 часов). Начиная с 17:00 мышей помещали поодиночке в клетку с едой, но без воды. В 19:00 каждой мыши давали две бутылки, одна с водой, а другая с 1% раствором сахарозы. Положение бутылок с водой и сахарозой меняли на 8, 9 и 10 часов.м. Количество потребленной воды и сахарозы измеряли путем взвешивания бутылок. Предпочтение сахарозы (%) рассчитывали как количество раствора сахарозы, потребленное по сравнению с количеством воды в течение 2 часов (8–10 часов вечера).
Испытание на подвешивание за хвост
Испытание на подвешивание за хвост проводили, как описано ранее 38,39 . Мышей индивидуально подвешивали на 6 мин, прикрепляя их хвосты клейкой лентой к потолку прямоугольной коробки, оставляя их на высоте 50 см над поверхностью стола.В течение 6-минутного периода подсчитывали кумулятивное время неподвижности. Неподвижность определяли как общее время, в течение которого все конечности и тело не двигались.
Тест принудительного плавания
Тест принудительного плавания проводился, как описано ранее 38,39 . Мышей помещали на 6 мин в прозрачный цилиндр из плексигласа (15 см в диаметре и 27 см в высоту), содержащий воду (глубина 15 см) при температуре 24°С. Суммарное время неподвижности учитывали за последние 5 мин. Неподвижность определялась как парение с неподвижными конечностями.
Анализ главных компонент и
K — кластеризация средних
Поведенческие особенности отдельных животных в множественных поведенческих тестах были проанализированы с использованием K — кластеризации средних, неконтролируемого алгоритма машинного обучения, который может разделять отдельные точки данных на отдельные подгруппы без ссылка на известные или помеченные исходы 70 . Анализ основных компонентов (PCA) был использован для преобразования поведенческих характеристик отдельных животных в двух поведенческих тестах в один собственный вектор в линейном измерении при следующих условиях: p < 0.05 теста Бартлетта на сферичность, ≥0,5 меры адекватности выборки Кайзера-Мейера-Олкина, ≥0,5 общности и ≥1,0 собственных значений в корреляционной матрице. Доля объясненной дисперсии (PVE) для уменьшения размера в настоящем исследовании находилась в диапазоне 73,4–86,3%. Все индивидуальные переменные в поведенческих тестах и все преобразованные значения были нормализованы в z баллов путем вычисления среднего значения (μ) и стандартного отклонения (σ). Оценки z были рассчитаны, как указано ниже.
$$z=(x-\mu )/\,\sigma$$
(1)
K — означает кластеризацию поведенческих признаков отдельных животных, измеренных с помощью трех поведенческих тестов, представленных в трехмерных матрицах. Поведенческие особенности отдельных животных в четырех-пяти поведенческих тестах и другие физиологические измерения отдельных животных визуализировались в трехмерной матрице для удобства после уменьшения размеров компонентов TST × FST или SIT × SPT.{2}\,$$
(2)
J = объективная функция, K = Количество кластеров ( K = 2, в этом исследовании), N = количество предметов группы, x
9 I = значение из поведенческих испытаний , c j = центроидное значение кластеров.
Наконец, доля (%) разделенных точек данных в двух кластерах была оценена для каждой экспериментальной группы.Кластеризацию PCA и K -means проводили с использованием программного обеспечения SPSS 25.0 (IBM, Armonk, NY, USA).
Статистический анализ
Статистический анализ проводили с использованием программного обеспечения GraphPad Prism 6 (GraphPad Software, Inc., Калифорния, США). Сравнение двух выборок проводили с использованием двустороннего критерия Стьюдента t , а множественные сравнения выполняли с использованием однофакторного дисперсионного анализа и апостериорного критерия Ньюмена-Кеулса. Все данные были представлены как среднее ± SEM, а статистические различия были приняты при p < 0.05, если не указано иное. Статистические данные результатов всех рисунков представлены в дополнительных данных 4.
Сводка отчета
Дополнительная информация о дизайне исследования доступна в Сводке отчета об исследовании природы, связанной с этой статьей.
Переключение визуального режима, усвоенное путем многократной адаптации к цвету
Основные версии:
1) Ключевым явлением, которое необходимо объяснить, является уменьшение начала быстрого эффекта после изменения условий освещения.Идея о том, что есть две стадии адаптации, одна из которых может быть обусловлена прогнозами внешней среды, захватывающая. Авторы продвигают вероятностную модель обнаружения изменений, но не сообщают подробностей. Следует упомянуть литературу о двухстадийных моделях адаптации (например, Pugh and Augenstein, 1977).
Мы согласны с тем, что в рукописи не было акцентировано внимание на предыдущей литературе по двухэтапным моделям адаптации, и наша редакция исправила эту оплошность.Мы также добавили спекулятивное обсуждение механистических моделей. В частности, теперь мы упомянем работу, упомянутую выше в нашем обсуждении, наряду с Fairchild и Reniff, (1995) и Rinner и Gegenfurtner, (2000), оба из которых также предлагают «быстрые» и «медленные» механизмы адаптации.
Подраздел «Отношение к предыдущей работе»:
«Принято считать, что цветовая адаптация имеет «быстрый» и «медленный» механизм и включает как рецепторные, так и пострецепторные зрительные процессы (например, Augenstein and Pugh, 1977; Fairchild and Reniff, 1995; Rinner and Gegenfurtner, 2000).Одна правдоподобная интерпретация наших результатов зависит от этих хорошо изученных механизмов; возможно, благодаря практике механизм быстрой адаптации стал способен производить более сильные и быстрые эффекты. В литературе по двигательному обучению эта возможность была названа «метаобучением», поскольку она влияет на параметры, определяющие скорость адаптации, что само по себе является своего рода обучением (Zarahn et al., 2008). Однако возможны и другие альтернативные механизмы, включая хранение и поиск адаптированных состояний (Lee and Schweighofer, 2009).В будущей работе будут изучены эти и другие возможности (см. также ниже)».
2) Авторы иногда изображают свой основной вывод как «увеличение степени адаптации зрительной системы сразу после надевания очков» или как обучение «быстрому переходу в частично адаптированное состояние». Твои не кажутся эквивалентными механизмами. Первый относится к нейронным цепям, которые адаптируются более быстро/экстенсивно после взаимодействия с окружающей средой. Второй предполагает эффект обучения, когда короткие временные рамки (т.г. секунды) адаптация такая же, но обучение позволяет цепям начать с форы. Было бы полезно четко изложить эти сценарии в тексте и использовать последовательную характеристику основного результата.
К сожалению, в этом очень важном вопросе рукопись была неясна, и мы соответствующим образом ее переработали. В исправленной рукописи мы теперь используем термин «быстрая корректировка» во введении и результатах. Мы добавили текст во Введение, утверждающий, что этот язык предназначен только для обозначения эмпирического наблюдения, что как только наблюдатели надевали очки, они лучше к ним приспосабливались и что ряд различных механизмов, таких как упомянутые выше, может объяснить это.Мы добавили к Обсуждению, что одна или обе приведенные выше интерпретации согласуются с нашими нынешними данными.
Введение:
«Мы предположили, что цветовая адаптация ускорится и/или получит фору в течение нескольких дней, так что наблюдатели испытают гораздо меньшее воспринимаемое изменение цвета мира, когда наденут красные очки, что свидетельствует о том, что они научились для переключения режимов. Поскольку это может включать механизмы, выходящие за рамки классической адаптации, мы будем использовать термин «быстрая адаптация» для обозначения этого возможного эмпирического свидетельства переключения режима — того, что как только наблюдатели надевали очки, их эффекты были менее заметными.В ходе обсуждения будут рассмотрены различные потенциальные механизмы, лежащие в основе сдвига».
Подраздел «Отношение к предыдущей работе»:
«Одна правдоподобная интерпретация наших результатов зависит от этих хорошо изученных механизмов; возможно, благодаря практике механизм быстрой адаптации стал способен производить более сильные и быстрые эффекты. В литературе по двигательному обучению эта возможность была названа «метаобучением», поскольку она влияет на параметры, определяющие скорость адаптации, что само по себе является своего рода обучением (Zarahn et al., 2008). Однако возможны и другие альтернативные механизмы, включая хранение и поиск адаптированных состояний (Lee and Schweighofer, 2009). В будущей работе будут изучены эти и другие возможности (см. также ниже)».
3) Интересно, что последействие не меняется с опытом. Хотя испытуемые быстрее приспосабливаются к очкам (например, они усвоили соответствующее состояние адаптации, красные точки на рис. 2), они не научились отменять эффект снятия очков (рис. 2, зеленые точки).Авторы должны обсудить, что они думают об этих выводах.
Это тоже важный момент, и мы сейчас обсудим его более подробно в статье. Мы провели более подробный анализ последействия, измеренного сразу после снятия очков, и обнаружили тенденцию к тому, что оно становится менее сильным в течение нескольких дней. Что касается нашей быстрой адаптации к очкам, мы исследовали изменения в течение дня в первом 5-минутном тесте, первом одноминутном блоке и первом индивидуальном матче (см. ответ на комментарий ниже).Это изменение последействия не было достоверным при p < 0,05 ни в одном из этих анализов, но было p < 0,1 во всех трех. Теперь мы обращаемся к этой тенденции в обсуждении, где мы предполагаем, что она предоставляет дополнительные доказательства улучшения переключения режимов с опытом. Снижение эффекта обучения в течение нескольких дней может быть связано с тем, что наблюдатели уже частично научились быстро переключаться на естественные условия наблюдения благодаря ежедневному опыту или даже эволюции.
Подраздел «Другие результаты настоящей работы»:
«В связи с этим последующий эффект, измеренный сразу после снятия красных очков, сместился к исходному уровню в течение нескольких дней, что означает более быструю адаптацию к знакомым естественным условиям с течением времени.Однако эта тенденция была относительно небольшой, имела лишь скромную статистическую достоверность и могла быть характерна для переключения из неестественных условий красных очков. Небольшой эффект, если он реальный, может быть связан с тем, что наблюдатели уже частично научились быстро приспосабливаться к естественной среде, что, как упоминалось выше, остается нерешенным спором».
4) Мы благодарим авторов за представление резюме индивидуальных различий, наблюдаемых в данных, которые велики. Происхождение этих различий неясно, и вместе со смешанными отчетами в нескольких исследованиях такого рода, о которых сообщается в литературе, требуется некоторая осторожность при интерпретации результатов.На ум приходит возможность того, что некоторые субъекты «думают о том, что они видят» и реагируют на этом основании. Это сложный вопрос, который мешает большинству исследований постоянства и не является причиной для задержки публикации настоящей работы, но его следует обсудить.
Это еще один важный момент, который мы добавили в наше Обсуждение. Прошлые работы показали, что наблюдатели могут демонстрировать различную степень экспериментально измеренного цветового постоянства в зависимости от того, просили ли их сделать суждения об отражательной способности поверхности или об отраженном свете (Arend and Reeves, 1986; Arend and Goldstein, 1987; Radonjić and Brainard, 2016). ).Наблюдатели потенциально могли использовать любую из этих стратегий, в зависимости от того, о чем они думали во время выполнения задачи. Чтобы устранить влияние выбора стратегии, мы дали инструкцию: «Ваша задача — настроить маленькое пятнышко на желтый цвет, в котором нет ни красного, ни зеленого, в зависимости от света, попадающего в ваш глаз. Старайтесь не думать о том, какого цвета должен быть пластырь» как в тестах с красными очками, так и без них. Тем не менее, все еще возможно, что некоторые наблюдатели, которые не следовали инструкциям, могли более или менее «думать» при вынесении своих уникальных желтых суждений.Теперь мы обсуждаем эту возможность в рукописи как один из источников больших индивидуальных различий, которые мы здесь обнаружили.
Подраздел «Индивидуальные отличия»:
«Каковы источники индивидуальных различий в способности научиться быстро переключаться между двумя состояниями? Прошлые работы показали, что наблюдатели могут демонстрировать очень разную степень экспериментально измеренного цветового постоянства в зависимости от того, просили ли их сделать суждения об отражательной способности поверхности или об отраженном свете (Arend and Reeves, 1986; Arend and Goldstein, 1987; Radonjić and Brainard, 2016).В данной задаче наблюдатели потенциально могут использовать любую из этих стратегий. Мы дали конкретные инструкции, чтобы ограничить влияние выбора стратегии (см. Материалы и методы), однако все еще возможно, что некоторые наблюдатели могли более или менее «думать» при вынесении своих уникальных желтых суждений. Это может быть одним из источников индивидуальных различий, которые мы здесь обнаружили».
5) Насколько вы уверены, что испытуемые соблюдали инструкции по ношению очков в течение дня? Может ли соответствие быть связано с индивидуальными различиями?
Мы считаем, что соответствие было хорошим, и приносим свои извинения за то, что не обсуждали его в оригинальной рукописи.В первый и второй день мы отправили наблюдателям текстовые напоминания о ношении очков в течение дня. Кроме того, каждый день, когда участники приходили на дневную сессию, ревизор лично спрашивал их, соблюдают ли они инструкции относительно ношения очков. Все сообщили, что носят очки по 5 часов каждый день. Два участника сообщили, что им пришлось отложить ношение очков примерно на 30 минут, один раз в один день, и, соответственно, их сеансы тестирования были отложены на 30 минут, чтобы обеспечить полное время «надевания очков».Возможно, что наблюдатели могли не сообщать о каком-либо несоблюдении, но мы считаем, что это вряд ли будет очень частым. Теперь мы обсуждаем соответствие в рукописи.
Подраздел «Индивидуальные отличия»:
«Теоретически их можно объяснить соблюдением правил ношения очков, но мы внимательно следили за соблюдением требований, и отказов было очень мало. Будущая работа может выяснить, могут ли индивидуальные различия в других аспектах восприятия цвета или зрения в целом объяснять индивидуальные различия в переключении режимов.
6) Постоянство цвета: Непонятно, в чем логика и как производились расчеты постоянства цвета.
Чтобы сделать идеальный прогноз постоянства, нужно начать с некоторого анализа того, какая отражательная способность поверхности соответствует уникальному желтому цвету в состоянии без очков, на основе некоторых предположений об окружающем освещении и некоторых ограничениях на функции отражательной способности поверхности. Затем спрашивается, какие настройки будут соответствовать этой функции отражения поверхности при измененном освещении (здесь вызванном очками).Подобный расчет, возможно, был сделан, но он не предусмотрен.
Почему предложенная метрика является мерой постоянства? Обоснование используемой метрики (и сделанных заявлений) является единственной ссылкой. Пожалуйста, поясните и помогите читателям лучше понять, как индекс фиксирует постоянство.
Это предложение является отличным подходом для исследований, проводимых с освещением и/или поверхностями, смоделированными с помощью устройства отображения. Наше понимание этого предположения заключается в том, что идеальное предсказание постоянства можно сделать, охарактеризовав уникальные желтые настройки наблюдателей с точки зрения основного физического стимула, в данном случае отражательной способности поверхности.Предсказание идеального постоянства состоит в том, что уникальный желтый цвет должен соответствовать одному и тому же коэффициенту отражения поверхности с красными стеклами и без них. Из-за использования технологии отображения и, возможно, задачи наблюдателя по настройке цвета, а не моделируемой отражательной способности, эксперимент может не иметь прямого доступа к нижележащим поверхностям и источникам света, и поэтому независимо от физического совпадения (т. е. в коэффициентах отражения) должны быть оценены с использованием подхода к моделированию, описанного рецензентом.
Иногда, однако, есть более прямой доступ к основной физике.В эксперименте можно, например, попросить наблюдателей выбрать уникальный желтый цвет из большого набора физических (не смоделированных) бумаг Манселла при различных физических источниках освещения. В этом случае идеальное предсказание совпадения будет состоять в том, что одна и та же бумага (с одинаковым коэффициентом отражения) будет выбрана для разных источников света. В этом случае для хорошего предсказания идеального постоянства не требуются никакие модельные расчеты.
Мы считаем, что использование цветных фильтров дает нам второй подход.В частности, предсказание идеального постоянства, т. е. соответствует ли физика спички, наблюдаемой в красных очках, физике спички, наблюдаемой без красных очков, в нашем случае сводится к тому, соответствует ли физическое содержание наблюдаемой сцены был идентичен в двух условиях просмотра. У нас нет доступа к предполагаемым функциям отражения наблюдателей, и действительно возможно, что они восприняли наш тест как источник света, а не как поверхность, но мы предполагаем, что если бы свет, выходящий из монитора, был настроен одинаково в очках и без них, тогда было сделано идеальное совпадение, потому что одинаковая физическая ситуация в мире воспринималась одинаково.То есть единственная разница между двумя уникальными желтыми настройками заключалась в том, что в одном случае наблюдатель носил красные очки — и физический мир, и создаваемое им восприятие (то есть уникальный желтый цвет) были постоянными в двух настройках. Нам это кажется разумным определением постоянства, и мы приносим свои извинения за то, что оно не было объяснено лучше в рукописи. Теперь мы делаем это более полно.
Теперь рецензент также правильно отмечает, что возникает дополнительный вопрос о том, какие единицы использовать для расчета того, насколько наблюдатель близок к предсказанию идеального постоянства.Конечно, возможно множество различных единиц, если они скорректированы с учетом эффектов красных очков, и они могут отражать либо то, насколько близок излучаемый свет к предсказанию идеального постоянства, либо насколько близок предполагаемый коэффициент отражения поверхности к этому предсказанию. . Чтобы избежать повторения наших результатов слишком много раз, теперь мы решили использовать для этой цели единицы, которые были предложены в комментарии 7), и добавили цитаты, подтверждающие их использование.
Подраздел «Базовый уникальный желтый цвет стал немного зеленее в течение нескольких дней»:
«Одно из определений идеального постоянства цвета — это когда один и тот же физический объект, поверхность или источник света, воспринимается как идентичный в различных условиях просмотра.В экспериментах на мониторах, где экспериментаторы имеют прямой доступ только к интенсивности пикселей, воспринимаемые поверхности обычно оцениваются с помощью моделирования вероятных источников света и поверхностей. Однако использование в нашем исследовании цветных очков дает нам более прямой подход.
В частности, если бы наблюдатели в нашем эксперименте имели идеальное цветовое постоянство, то одни и те же физические пиксели на мониторе, независимо от того, воспринимались ли они как поверхности или как источники света (наш эксперимент был неоднозначным в этом отношении), должны казаться уникальными желтыми как с, так и с без очков, несмотря на резкое влияние очков на спектр света, попадающего в глаз.Если эти условия выполняются, то единственное различие между двумя уникальными настройками желтого цвета будет заключаться в различии условий просмотра: то есть один и тот же физический мир (пиксели монитора) будет восприниматься одинаково (т. е. уникальный желтый цвет) в двух ситуациях, разумный определение идеального постоянства цвета.
Чтобы оценить величину постоянства, мы охарактеризовали физический цвет, достигающий глаза, используя относительное усиление длинноволновых (L) и средневолновых (M) фоторецепторов.Эта мера предполагает, что уникальные настройки желтого цвета соответствуют точке баланса между откликами колбочек L и M, где масштабный коэффициент (усиление) может применяться к откликам одного из классов колбочек: L = k*M. Влияние адаптации или другой пластичности на уникальный желтый цвет можно количественно определить, решив k, который равен L/M (Neitz et al., 2002)».
7) Очевидно, что существует несколько различных способов представления этих данных. Угол оттенка в стимуле четкий и прямой, и соотношение L/(L+M), использованное на рис. 4, также точное.Другой альтернативой является относительное усиление колбочек L и M при уникальном желтом цвете при допущении, что уникальный желтый цвет представляет L-kM = 0 для некоторого k в каждом состоянии адаптации. Тогда это дает k = L/M. Приведет ли это ориентированное на выигрыш выражение данных к большему пониманию, чем версия L/(L+M)? Возможно, стоит взглянуть, поскольку он отсылает данные к гипотетическому механистическому состоянию зрительной системы.
Это отличное предложение: мы согласны с тем, что относительное усиление колбочек L и M в уникальном желтом цвете лучше характеризует механизм корректировки весовых коэффициентов колбочек.Это было ранее использовано Нейцем (2002) для выявления пластического нейронного механизма, который компенсирует большие различия в соотношении колбочек L и M в сетчатке в разных популяциях. Теперь мы перестроили рисунок 4 в единицах L/M. Затем мы количественно определяем постоянство цвета с помощью этого устройства, обращаясь к комментарию 6). Это рассчитывается как отношение между (1) евклидовым расстоянием между базовой линией и первой уникальной желтой настройкой в очках и (2) расстоянием между базовой линией и предсказанием полной адаптации/идеального цветового постоянства (см. выше).
Будет ли просмотр этого представления или представления L/(L+M) в расширенном масштабе времени сеанса, отдельно для каждого сеанса, раскрывать больше динамики, особенно если (как предлагается ниже в контексте рисунка S2) отдельные настройки сделанные один раз в минуту во внутрисеансовых блоках, показывались явно.
Мы согласны с тем, что также важно рассмотреть более тонкую динамику эффектов адаптации. Теперь мы разъясняем в документе, что каждая точка на нашем исходном графике представляет собой средние настройки для 5-минутного теста, и что каждый тест, в свою очередь, состоит из 5 одноминутных блоков.В каждом блоке наблюдатели устанавливают несколько совпадений, и мы можем использовать их для более точного изучения хода времени, что мы и делаем в ответ на комментарий 10) ниже.
Рисунок 2 — дополнение к рисунку 2 показывает уникальные настройки желтого цвета, представленные в виде угла оттенка, для каждого 1-минутного блока. Чтобы сохранить участок управляемого размера, мы усреднили утреннюю и дневную сессию в течение дня, получив в общей сложности 250 блоков. Это усреднение не повлияло на общие тенденции в данных. На рисунке одна точка представляет один блок, а 5 соединенных точек показывают 5 блоков каждого теста.Верхняя часть рисунка представляет настройки, выполненные без очков. Черные точки — это базовые настройки перед тем, как надеть красные очки. В нижней части рисунка показаны настройки, выполненные в очках. По причинам, описанным в комментарии 10) ниже, средние значения на этом рисунке были рассчитаны при исключении первого совпадения из тестов 2-5, хотя, опять же, это не повлияло на общую картину наших результатов.
Мы включаем это в дополнительный материал к статье (Рисунок 2 — дополнение к рисунку 2), заменяя части наших предыдущих дополнительных рисунков, которые также показывали детали времени.Несколько факторов усложняют интерпретацию сюжета: нельзя просто «соединить все точки», чтобы увидеть мелкомасштабное представление временного хода адаптации. Во-первых, между тестами проходит разное количество времени, от 5 до 15 минут (полный график тестирования см. на рис. 1C). Кроме того, в промежутках между тестами наблюдатели просматривали неконтролируемые стимулы, хотя в очках, в зависимости от обстоятельств, они могли существенно отличаться по цветовой статистике от нашего тестового дисплея (см. также ответ на комментарий 10)).Наконец, в тесте, безусловно, была адаптация к самому тестовому дисплею. Учет этих факторов для того, чтобы сделать выводы о точном временном ходе, затруднен, и мы планируем рассмотреть эту проблему в будущей публикации, поскольку точное время обычно является ортогональной точкой для нашей гипотезы о переключении режима. Тем не менее, в данных есть интересные тенденции, и мы планируем поделиться нашим полным набором данных с заинтересованными читателями. Обратите внимание, что эти факторы не делают недействительными наши анализы, в которых используется только первый 1-минутный блок (или первое совпадение) после надевания или снятия очков, чтобы охарактеризовать эффекты во время сеансов тестирования.
Подраздел «Привыкание к очкам стало быстрее и сильнее»:
«Рисунок 2 — дополнение к рисунку 2 показывает полную временную динамику наших результатов в зависимости от минутных блоков. У нас не было априорных ожиданий относительно тонких тенденций от блока к блоку, поэтому мы оставили их изучение для будущей работы».
8) Одним из аспектов дизайна, который, вероятно, важен, является то, что наблюдатели чередовали две среды несколько раз в день. Возможно, важным фактором является частота изменений окружающей среды.Другой фактор заключается в том, что среда, в которой они находились, была умеренно сложной, офис.
На рис. 1 видно, что окружающая среда для эксперимента менялась день ото дня с изменениями наружного освещения, проникающего через окна. Была ли какая-то характеристика этого? Обеспокоены ли авторы тем, что это изменение могло повлиять на результаты? Могут ли они дать какое-либо руководство будущим исследователям, которые хотели бы попытаться воспроизвести свои эксперименты как можно точнее, с точки зрения размера комнаты, относительной площади, занимаемой окнами, внутреннего освещения, уровня окружающего освещения по отношению к дисплею и т. д.
Важно отметить, что частота изменений в окружающей среде может влиять на эффект обучения, который мы здесь наблюдаем; в настоящее время мы проводим последующие эксперименты, в которых испытуемые носили те же пары очков, что и в настоящем исследовании, в течение 5 часов непрерывно в день в течение 5 дней, что приводит к меньшему чередованию двух сред в течение каждого дня. Повышенная адаптация к очкам также была обнаружена у 4 испытуемых, которых мы тестировали до сих пор. Дальнейший анализ будет проведен после того, как мы соберем больше данных, но сбор идет медленно из-за пандемии, поэтому мы предпочитаем опубликовать этот результат в будущей статье.На данный момент мы добавили это как будущее направление в обсуждение.
«Проверка того, влияют ли такие факторы, как частота изменений окружающей среды, на наблюдаемый нами эффект обучения, является важным направлением будущих исследований».
Мы приносим искренние извинения за недопонимание из-за нашего нечеткого описания рисунка 1. На рисунке 1D показано изображение, которое наблюдатели просматривали на экране компьютера во время тестирования, когда они выполняли уникальные настройки желтого цвета.Он не отображает саму нашу тестовую среду. Фиксированное изображение офиса (не визуализированная сцена, не изменявшаяся со временем) было представлено на тестовом дисплее, чтобы дать наблюдателям контекстную информацию при внесении корректировок; в частности, чтобы визуальной системе было очевидно, что они носят очки. Это может быть не так, если тестирование проводилось при просмотре небольшого пятна на черном экране.
Наш эксперимент проводился в подземной комнате без окон, а окружающее освещение обеспечивалось стабильными верхними комнатными светильниками.Экран просматривался через 3-футовый «туннель» с войлочной подкладкой, так что окружающий свет, попадающий на наш тестовый дисплей, не был существенным фактором. Субъекты сидели перед туннелем, их головы находились у входа в туннель. Теперь мы опишем нашу испытательную комнату и более четко покажем ее в рукописи. Наконец, теперь мы также поясняем, что между сеансами тестирования наблюдатели свободно перемещались в своей повседневной жизни, испытывая различные условия освещения.
Обозначение рисунка 1D:
«Фиксированное изображение офиса и линии горизонта было представлено на тестовом дисплее, чтобы предоставить наблюдателям контекстную информацию при выполнении настроек.
Наблюдатели наблюдали за тестовым дисплеем через трехфутовый туннель, обитый войлоком, на откалиброванном мониторе в полностью освещенной лабораторной комнате».
Подраздел «Процедура»:
«Наблюдатели завершили все тесты в полностью освещенной комнате (без окон) с целью измерения перцептивного опыта в контексте, похожем на их естественную среду, при адаптации к очкам. Экран просматривался через 3-футовый «туннель» с войлочной подкладкой, поэтому окружающий свет, попадающий на наш тестовый дисплей, не был значительным фактором.Наблюдатели сидели перед «туннелем», положив головы на подставку для подбородка, расположенную у его входа».
9) Расскажите, пожалуйста, немного больше о конвертации в мегабайты и отображаемых стимулах.
a) Как были масштабированы основные параметры конуса пиков относительно друг друга при расчете LMS для последующего расчета L/L+M и S/L+M. Похоже, что эти две величины — Lmb и Smb, переданные при вычислении LM и S. Не уверен, что S — лучший выбор обозначения для последнего.
b) Какая функция фотопической светимости использовалась для определения номинальной изосветимости. Учитывая использование основ Стокмана-Шарпа, можно сделать вывод, что новый стандарт CIE представляет собой взвешенную сумму этих, или вы могли бы использовать CIE 1931, или Джадда-Воса, или…
Теперь мы предоставим более подробное описание преобразования в мегабайтное пространство и отображаемые стимулы. Мы добавили следующую информацию в наш раздел «Материалы и методы».
Подраздел «Процедура»:
«Стимулы были созданы с использованием модифицированной версии цветового пространства Маклеода-Бойнтона (MacLeod and Boynton, 1979), масштабированы и сдвинуты таким образом, чтобы начало координат соответствовало номинальной точке белого источника света C, и чтобы чувствительность была примерно уравнена по двум оси (Вебстер и др., 2000).
Мы начали с вычисления откликов колбочек из спектра стимулов, используя основные принципы колбочек Смита и Покорни (1975), масштабированные таким образом, чтобы сумма откликов колбочек L и M была равна 1, а ответы S-колбочек, деленные на эту сумму, также равнялись 1. Мы затем вычисляются начальные координаты в цветовом пространстве Маклеода-Бойнтона как r мб = (LM)/(L+M) и b мб = S/(L+M). Наконец, мы масштабировали и сместили эти координаты:
.
лм = (r мб -.6568) х 2168
S = (b мб –.01825) х 6210
Где LM — масштабированная красно-зеленая координата, а S — масштабированная координата S-конуса, 0,6568 и 0,01825 — координаты Маклеода-Бойнтона источника света C, а 2168 и 6210 — константы, которые масштабируют оси LM и S так, что значение 1 примерно равно порогу обнаружения (Webster and Mollon, 1995).
Все настройки упали вдоль номинально изо-яркой плоскости (определяемой осями LM и S, с яркостью, установленной на 51 кандела/м 2 ), когда очки не были надеты, чтобы уменьшить влияние яркости на суждения.Функцией фотопической светимости, которую мы использовали для определения номинальной светосилы, была CIE Photopic V(λ), модифицированная Джаддом (1951)».
Для расчета L/M в исправленной рукописи (и L/(L+M) в оригинале) мы использовали основные принципы конуса Стокмана и Шарпа (2000) с пиками основных параметров конуса, масштабированными до 1.
b) Укажите фактический радиус круга цветового тона, используемого при регулировке, а также расстояние между углами тона для грубой и точной регулировки, чтобы кто-то мог произвести ваши стимулы.
Радиус цветового круга, использованного в этом исследовании, равнялся 80, что соответствует хроматическому контрасту. Этот контраст находится на номинальном пороге и поддерживался постоянным во время процедуры настройки. Более грубые и точные шаги регулировки составляли +/- 5 и +/- 1 градус соответственно на каждое нажатие кнопки. Теперь мы сообщаем об этих параметрах в разделе «Материалы и методы».
Подраздел «Процедура»:
«Радиус используемого цветового круга составлял 80, что соответствует хроматическому контрасту, примерно в 80 раз превышающему порог обнаружения (см. выше), и оставался постоянным во время процедуры настройки.
Наблюдатели могли регулировать угол стимула с более грубыми или тонкими шагами в 5 или 1 градус оттенка соответственно за одно нажатие кнопки».
c) Был ли использован полный круг оттенков или были конечные точки? Если конечные точки, насколько вы уверены, что испытуемые не использовали их в качестве эталона и не считали шаги оттуда или менее явно привязывали свои корректировки к предполагаемой средней точке предоставленного диапазона? Изучение таких стратегий может маскироваться под обучение адаптации.
Это также важный момент, который мы сейчас добавляем в исправленную рукопись. Мы не использовали полный круг оттенков, и у нас была зеленая конечная точка с углом оттенка 200 градусов и красная конечная точка с углом оттенка 360 градусов. В начале каждого испытания угол оттенка стимулов устанавливался случайным образом в диапазоне 290 ± 45 градусов. Мы отслеживали ответы наблюдателей и сохраняли каждый шаг их корректировок. Только несколько наблюдателей в некоторых блоках в начале эксперимента достигли зеленого или красного предела, что делает маловероятным их использование в качестве опорной точки.
Подраздел «Процедура»:
«Нажатие кнопки не имело никакого эффекта, как только наблюдатели достигли зеленой конечной точки с углом оттенка 200 градусов и красной конечной точки с углом оттенка 360 градусов. В начале каждого испытания угол оттенка стимулов устанавливался случайным образом в диапазоне 290 ± 45 градусов. Мы отслеживали ответы наблюдателей и сохраняли каждый шаг их корректировок. Изучение этих данных подтвердило, что они не использовали красную или зеленую конечную точку в качестве привязки для своих настроек (например, всегда двигались к конечной точке, а затем перемещались на фиксированное количество шагов назад).
d) Какова яркость настраиваемого тестового участка? В подразделе «Аппарат» говорится, что яркость фона составляла 41,85 кд/м2, но позже и на картинке это описывается/показывается как черный цвет, что удивительно, если только окружающая среда в комнате не была очень яркой.
Мы приносим свои извинения за нечеткость описания нашего тестового дисплея и недопонимание, вызванное рис. 1D. Как отмечалось выше, на рисунке 1D показано, что было представлено на мониторе наблюдателям, когда они делали уникальные настройки желтого цвета.Сцена в офисе — это не та комната, в которой тестировались наблюдатели; это фоновое изображение, отображаемое на мониторе и предназначенное для предоставления наблюдателям контекста во время внесения корректировок. Угол обзора черного квадрата составлял 5,7 градуса, и он отделяет тестовую заплатку 0,5 градуса от офисного изображения (фон). Яркость фона составляла 20 кд/м2, а яркость тестового участка — 51 кд/м2. Теперь мы добавили эту информацию в статью.
Подраздел «Процедура»:
«Средняя яркость фонового изображения офиса составила 20 кандел/м 2 .
«Все настройки упали вдоль номинально изо-яркой плоскости (определяемой осями LM и S, с яркостью, установленной на 51 кандела/м 2 ), когда очки не были надеты, чтобы уменьшить влияние яркости на суждения».
10) Рисунок S2, интерпретация верхней панели. Характер результатов немного сложно интерпретировать. Мы ожидаем наименьшую адаптацию для первой настройки, поэтому в целом они должны быть ниже по оси Y, чем соответствующие точки на рисунке 2.Во многих случаях это не так. См., например, первую группу очков в настройках. Любой комментарий? Первые настройки просто очень шумные? Было бы понятнее, если бы каждая отдельная настройка была нанесена на график, а не просто дано сравнение первой со средним значением.
Мы благодарим рецензента за наблюдение и согласны с тем, что первая уникальная настройка желтого цвета, сделанная в очках в каждом тесте, должна быть зеленее/ниже по оси Y, чем последующие совпадения.Итак, мы рассмотрели первый матч в первом блоке каждого теста (для определения терминов: каждый пятиминутный тест состоял из 5 одноминутных блоков; каждое утреннее или дневное занятие состояло из 5 тестов в очках, разделенных 5-10 минут по графику, показанному на рис. 1С рукописи). Мы обнаружили, что в очках первое совпадение в первом блоке во всех тестах, кроме первого, в каждом сеансе было неожиданно больше по углу оттенка (менее зеленого), чем последующие совпадения в этом блоке. Но первый матч в первом блоке первого теста сразу после надевания очков ничем не отличался от других матчей.
Этот шаблон показан на изображении ответа автора 1, на котором показаны первые восемь отдельных совпадений в каждом из 5 блоков, усредненные по дням. Связанные красные точки — это первые совпадения в каждом блоке, связанном между тестами, а остальные цветные линии — последующие 2–8 совпадений в каждом блоке. Первые совпадения в первом блоке тестов 2-5, сделанные в очках, были намного выше по оси ординат, чем другие совпадения (т. е. красные линии в тестах 2-5 для блока 1 являются выбросами).
Эта закономерность имеет простое объяснение: тесты 2-5 проводились после 5-10-минутного перерыва, во время которого наблюдатели смотрели видео или писали текстовые сообщения на своих телефонах или ноутбуках.Однако первый тест сразу же последовал за «базовым» тестом без очков, когда наблюдатели смотрели на наш тестовый дисплей. По мнению большинства наблюдателей, экраны телефонов или компьютеров, которые они смотрели во время перерыва, выглядели более красными, чем другие объекты в окружающей среде, когда они носили очки, вероятно, потому, что они имели более высокую яркость, чем другие предметы в помещении. Это было особенно верно для белого фона во время текстовых сообщений. Таким образом, после просмотра экрана своего телефона/ноутбука в течение определенного периода времени наблюдатели, вероятно, стали более приспособленными к красному оттенку, чем после просмотра нашего тестового дисплея.Затем, когда наблюдатели переходили со своего телефона/компьютера на тестовый дисплей, они находились в более адаптированном состоянии, поэтому они устанавливали первое уникальное желтое совпадение более красным, чем другие совпадения, которые были сделаны после того, как некоторое время смотрели на тестовый дисплей. Этот эффект отсутствовал при первом совпадении в первом блоке первого теста при надевании очков, потому что не было перехода между разными дисплеями до первого теста (который проводился сразу после базового теста).
Поскольку эти 4 балла (первое совпадение первого блока в очках в тесте 2-5), по-видимому, были получены в разных условиях, мы удалили их при вычислении средних значений блоков на рис. во всех блоках теста на них не повлияло включение этих 4 точек в заметной степени, поэтому мы оставили их без изменений (хотя мы убедились, что все статистические тенденции сохраняются, если исключить выбросы).Обратите также внимание на то, что на цифры, отображающие только первый тест в очках (т. е. на диаграммы рассеяния на рис. 3 и на рис. 3 — дополнение 1), эти выбросы также не влияют из-за непосредственного перехода от базового теста. Однако наш предыдущий график всего временного хода с использованием первого совпадения, как заметил рецензент, был искажен, как мы теперь знаем, этими точками-выбросами, и поэтому мы удалили его из рукописи.
https://дои.org/10.7554/eLife.61179.sa2
Отрицание: когда помогает, когда вредит
Отрицание: когда помогает, когда причиняет боль
Отрицание — это механизм выживания, который дает вам время, чтобы приспособиться к неприятным ситуациям, но оставление в отрицании может помешать лечению или вашей способности решать проблемы.
Персонал клиники Майо
Если вы отрицаете, вы пытаетесь защитить себя, отказываясь принять правду о том, что происходит в вашей жизни.
В некоторых случаях первоначальное краткосрочное отрицание может быть полезным, поскольку оно дает вам время приспособиться к болезненной или стрессовой проблеме. Это также может быть предвестником каких-то изменений в вашей жизни. Но у отрицания есть темная сторона. Узнайте, когда отрицание вредно для здоровья и как его преодолеть.
Понимание отрицания и его цель
Отказ признать, что что-то не так, — это способ справиться с эмоциональным конфликтом, стрессом, болезненными мыслями, угрожающей информацией и беспокойством.Вы можете отрицать все, что заставляет вас чувствовать себя уязвимым или угрожает вашему чувству контроля, например, болезнь, зависимость, расстройство пищевого поведения, личное насилие, финансовые проблемы или конфликты в отношениях. Вы можете отрицать, что что-то происходит с вами или с кем-то еще.
Когда вы отрицаете, вы:
- Не признает трудную ситуацию
- Старайтесь не смотреть в глаза фактам проблемы
- Преуменьшение возможных последствий проблемы
Когда отрицание может быть полезным
Отказ смотреть фактам в лицо может показаться нездоровым.Иногда, однако, короткий период отрицания может быть полезен. Отрицание дает вашему разуму возможность бессознательно поглощать шокирующую или тревожную информацию в темпе, который не приведет вас к психологическому штопору.
Например, после травматического события вам может потребоваться несколько дней или недель, чтобы осознать, что произошло, и справиться с предстоящими трудностями. Представьте, что может случиться, если вы обнаружите ком в горле. Вы можете почувствовать прилив страха и адреналина, когда представите, что это рак.
Итак, вы игнорируете припухлость, надеясь, что она исчезнет сама по себе. Но когда комок все еще там неделю спустя, вы обратитесь к врачу.
Этот тип отрицания является полезной реакцией на стрессовую информацию. Вы изначально отрицали мучительную проблему. Но по мере того, как ваш разум усваивал эту возможность, вы начали подходить к проблеме более рационально и начали действовать, обратившись за помощью.
Когда отрицание может быть вредным
А что, если бы вы продолжали отрицать шишку? Что, если бы вы никогда не обращались за помощью? Если отрицание сохраняется и мешает вам предпринять соответствующие действия, например, проконсультироваться с врачом, это вредная реакция.
Рассмотрим эти примеры нездорового отрицания:
- Студент колледжа становится свидетелем жестокой стрельбы, но утверждает, что на него это не повлияло.
- Партнер пожилого мужчины на последнем этапе жизни отказывается обсуждать с ним медицинские указания и завещания, настаивая на том, что ему становится лучше.
- Кто-то периодически пропускает утренние рабочие собрания из-за чрезмерного выпивки накануне вечером, но настаивает на том, что это не проблема, потому что работа все еще выполняется.
- Супружеская пара звонит так много долгов по кредитной карте, что они отбрасывают счета в сторону, потому что они не могут их открыть.
- Родители подростка-наркомана продолжают давать своему ребенку деньги на «одежу».
- Человек с болью в груди и одышкой не верит, что эти симптомы сигнализируют о сердечном приступе, и медлит с обращением за помощью.
В подобных ситуациях отказ может помешать вам или вашему близкому получить помощь, например, лечение или консультацию, или решить проблемы, которые могут выйти из-под контроля — и все это с потенциально разрушительными долгосрочными последствиями.
Проходя мимо отрицания
Столкнувшись с ошеломляющим поворотом событий, нормально сказать: «Я просто не могу думать обо всем этом прямо сейчас». Возможно, вам потребуется время, чтобы разобраться с тем, что произошло, и адаптироваться к новым обстоятельствам. Но важно понимать, что отрицание должно быть лишь временной мерой — оно не изменит реальной ситуации.
Не всегда легко понять, сдерживает ли вас отрицание. Сила отрицания может меняться со временем, особенно у людей с хроническими заболеваниями — некоторые периоды связаны с меньшей защитой, а в другие периоды отрицание может быть намного сильнее.Однако, если вы чувствуете, что зашли в тупик, или если кто-то, кому вы доверяете, предполагает, что вы отрицаете, вы можете попробовать следующие стратегии:
- Честно изучите, чего вы боитесь.
- Подумайте о потенциальных негативных последствиях бездействия.
- Позвольте себе выразить свои страхи и эмоции.
- Попытайтесь выявить иррациональные убеждения относительно вашей ситуации.
- Журнал о вашем опыте.
- Откройтесь надежному другу или любимому человеку.
- Примите участие в группе поддержки.
Если вы не можете справиться со стрессовой ситуацией самостоятельно — вы застряли на этапе отрицания — подумайте о том, чтобы обратиться к специалисту по психическому здоровью. Он или она может помочь вам найти здоровые способы справиться с ситуацией, а не пытаться притворяться, что ее не существует.
Когда близкому человеку нужна помощь, чтобы преодолеть отрицание
Вас может расстраивать, когда кто-то, кого вы любите, отрицает важную проблему.Но прежде чем требовать, чтобы ваш любимый человек смотрел правде в глаза, сделайте шаг назад. Попытайтесь определить, нужно ли ему или ей просто немного времени, чтобы решить проблему.
В то же время дайте человеку понять, что вы открыты для разговора на эту тему, даже если вам обоим будет неудобно. В конечном счете, это может дать вашему любимому человеку уверенность, необходимую ему или ей, чтобы двигаться вперед. Ваш любимый человек может даже почувствовать облегчение, когда вы поднимете этот вопрос.
Если ваш близкий человек отрицает серьезную проблему со здоровьем, такую как депрессия, рак или зависимость, обсуждение этой проблемы может быть особенно трудным.Выслушайте и предложите свою поддержку. Не пытайтесь заставить кого-то обратиться за лечением, это может привести к гневным столкновениям. Предложите встретиться вместе с врачом или психиатром.
Получите самую свежую медицинскую информацию от экспертов Mayo Clinic.
Зарегистрируйтесь бесплатно и будьте в курсе научных достижений, советов по здоровью и актуальных тем, связанных со здоровьем, таких как COVID-19, а также экспертных знаний по управлению здоровьем.
Узнайте больше об использовании данных Mayo Clinic.
Чтобы предоставить вам наиболее актуальную и полезную информацию, а также понять, какие
информация полезна, мы можем объединить вашу электронную почту и информацию об использовании веб-сайта с
другая информация о вас, которой мы располагаем. Если вы пациент клиники Майо, это может
включать защищенную информацию о здоровье.Если мы объединим эту информацию с вашей защищенной
медицинской информации, мы будем рассматривать всю эту информацию как
информацию и будет использовать или раскрывать эту информацию только так, как указано в нашем уведомлении о
практики конфиденциальности. Вы можете отказаться от получения сообщений по электронной почте в любое время, нажав на
ссылка для отписки в письме.
Подписаться!
Спасибо за подписку
Наш электронный информационный бюллетень Housecall будет держать вас в курсе самой последней медицинской информации.
Извините, что-то пошло не так с вашей подпиской
Повторите попытку через пару минут
Повторить попытку
09 апреля 2020 г.
Показать ссылки
- Левенсон Дж.Л.Психологические факторы, влияющие на другие заболевания: клинические признаки, оценка и диагностика. http://www.uptodate.com/home. По состоянию на 7 февраля 2017 г.
- Уолш Д. Отрицание и способность принимать решения. В: Паллиативная медицина. Филадельфия, Пенсильвания: Saunders Elsevier; 2009. http://www.clinicalkey.com. По состоянию на 7 февраля 2017 г.
- Шварц МЗ. Реакции пациента. В: Учебник физической диагностики: История и экспертиза. 7-е изд. Филадельфия, Пенсильвания: Saunders Elsevier; 2014.http://www.clinicalkey.com. По состоянию на 7 февраля 2017 г.
- Старзомская М. и др. Подводные камни в исследовании нервной анорексии: риск артефактов, связанных с отрицанием болезни и методами их предотвращения. Психиатрия Данубина. 2016;28:202.
- Приспосабливаются к жизни после того, как попали в заложники или были похищены. Американская психологическая ассоциация. http://www.apa.org/helpcenter/hostage-kidnap.aspx. По состоянию на 7 февраля 2017 г.
- Клык XY и др. Является ли отрицание неадекватным механизмом выживания, который удлиняет догоспитальную задержку у пациентов с инфарктом миокарда с подъемом сегмента ST? Журнал психосоматических исследований.2016;91:68.
- Прорыв барьеров отрицания. Сеть действий по уходу. http://caregiveraction.org/breaking-through-barriers-denial. По состоянию на 7 февраля 2017 г.
Подробнее
.
Родители чувствуют, что они «не справляются» с возвращением в школу
В первые дни пандемии, когда школы внезапно закрылись, а миллионы сотрудников были отправлены на работу из дома, многие родители с нетерпением ждали падения, когда момент, когда они верили, что жизнь вернется в нормальное русло.
Но возвращение в школу этой осенью совсем не рутина, поскольку школы по всей стране частично или полностью переходят на дистанционный режим. То, что многие считали краткосрочной проблемой, может перерасти в 2021 год и далее. В результате работающие родители паникуют, пытаясь понять, как совмещать дистанционное обучение и работу на полный рабочий день.
Многих беспокоит их способность поддерживать бесперебойную работу, не забывая о детях и рабочих обязанностях.Более половины, 54%, работающих родителей говорят, что они чувствуют себя виноватыми, потому что они не могут в полной мере заботиться о своих детях, а 43% сообщают о чувстве вины, когда они заботятся о своих семьях, потому что они не сосредотачиваются на своих рабочих обязанностях. к недавнему опросу Catalyst «Влияние Covid-19 на работающих родителей». В августе в онлайн-опросе приняли участие 1000 работающих родителей детей школьного возраста в США.
«У меня постоянно что-то не получается, — говорит 36-летняя Элизабет Виггс.«Выбор заключается в том, что либо я плохо работаю на работе, либо плохо воспитан. и быть родителем — чувствовать, что у них просто огромные трещины, пробегающие по их основанию. Это похоже на кризис идентичности».
Менеджер по маркетингу из Сиэтла Элизабет Виггс и ее дочь. часов вне традиционного рабочего дня.Менеджер по маркетингу компании Zillow из Сиэтла, Виггс обычно просыпается около 4:30 утра, чтобы приступить к работе до того, как проснется ее 6-летняя дочь. «Обычно это дает мне по крайней мере два часа. Скрестим пальцы, она ложится спать поздно, и тогда у меня появляется немного больше времени», — говорит Виггс.
Но как только ее дочь встает, ее день становится немного более сложным, тем более что школа ее дочери этой осенью полностью удалена. «Я не пользовался услугами по уходу за детьми, — говорит Виггс. «Моя работа требует, чтобы я много времени проводила на собраниях, поэтому [моя дочь] проводит много времени перед телевизором.Это моя забота о детях».
Виггс делит опеку с отцом своей дочери. «Это помогает немного снять напряжение», — говорит она, но добавляет, что ее дочь проводит с ней большую часть рабочей недели. с папой — просто вау — я только и делаю, что работаю. Но как приятно просто сосредоточиться только на чем-то одном, поэтому я вхожу в сумасшедший режим гиперфокусировки». родителей, опрошенных Catalyst, планируют сократить работу этой осенью, если их дети не вернутся в школу на полный рабочий день.В том числе и те, кто решил сократить рабочий день, перейти на неполный рабочий день и даже уволиться с работы, по крайней мере временно.
Увеличить IconArrows, указывающие наружу
«Я не думаю, что это исчезнет в ближайшее время», — говорит Лаура Никель, 35-летняя мать-одиночка, живущая в пригороде Чикаго. Имея это в виду, Никель говорит, что она думала о том, как морально и физически подготовить себя и свою 7-летнюю дочь к этому в долгосрочной перспективе.
Чтобы заставить это работать, Никель начала свою работу на день раньше, но она также группирует все звонки своих клиентов по частям в течение дня, если может.Как правило, она старается закончить работу во второй половине дня, когда заканчиваются школьные занятия ее дочери в Zoom. «Мы можем вместе обедать каждый день, кататься на велосипеде или прыгать на батуте», — говорит Никель, добавляя, что хочет, чтобы пандемия прошла для ее дочери как можно более позитивно.
Директор по работе с клиентами небольшой PR-фирмы, специализирующейся на технологических компаниях и стартапах, Никель говорит, что, хотя ее работодатель поддерживает ее, она единственный родитель в своей компании, у которого есть ребенок школьного возраста.И она говорит, что ей решать, какая поддержка ей нужна, и просить о ней. Недавно ее компания рассмотрела вопрос о снижении целевых показателей оплачиваемых часов для всех сотрудников на год, и Никель поддерживает этот шаг.
Чтобы помочь своей дочери с дистанционным обучением этой осенью, Лаура Никель установила детский стол и систему корзин для всех своих школьных принадлежностей.
Источник: Лаура Никель
Некоторые работодатели активизируются, но достаточно ли этого?
«Трудно совмещать работу и рождение ребенка», — говорит Виггс, несмотря на то, что она чувствует большую поддержку со стороны своего работодателя.В начале пандемии Zillow предложила родителям возможность работать по более гибкому графику, а также предоставила 15 дней резервного ухода через Bright Horizons и 10 дней оплачиваемого «времени по уходу», которое можно использовать с перерывами и в дополнение к обычные оплачиваемые отпуска и больничные.
Фактически, Zillow является одним из нескольких крупных работодателей, которые увеличили пособия по уходу за детьми через таких партнеров, как Bright Horizon, в условиях пандемии. В прошлом месяце Citi объявил об увеличении количества льготных резервных дней по уходу за ребенком с 20 до 40 для сотрудников.Кроме того, финансовая компания развернула услуги по трудоустройству няни, а также отменила членские взносы и предложила скидки Sittercity.
Для тех родителей, которые занимаются дистанционным обучением, Citi предлагает услуги, которые могут помочь родителям найти преподавателей, которые помогут контролировать онлайн-обучение. А если сотрудники заинтересованы в домашнем обучении в небольших группах, они могут запросить помощь в поиске преподавателей и налаживании контактов с другими семьями.
«Мы хотели быть максимально гибкими, чтобы поддерживать наших коллег, — говорит Сара Вехтер, руководитель отдела кадров Citi.«Сейчас так много неуверенности в том, как долго это продлится, и мы все будем дома, поэтому мы решили не предлагать определенное количество дополнительных оплачиваемых выходных».
«Мы понимаем важность качественного ухода за детьми и то, насколько тяжело дистанционное обучение для работающих родителей», — говорит Вехтер CNBC Make It, добавляя, что она лично сталкивается с этим каждый день как мать 10-летних близнецов. «Поскольку мы все проводим большую часть времени дома, границы между работой и личным временем стираются.»
В этой ситуации, по словам Вехтер, у работодателей есть возможность сыграть большую роль, помогая сотрудникам справляться со всеми проблемами, с которыми они сталкиваются, в том числе справляться с семейными обязанностями, а не только с их рабочей нагрузкой.
Увеличить IconArrows, указывающие вовне , Bank of America и Microsoft также предлагают сотрудникам возможность отправлять своих детей в «учебные центры», управляемые Bright Horizons, которые предлагают присмотр за детьми школьного возраста в небольших группах, по словам представителя службы ухода за детьми. Компания.Благодаря партнерским отношениям с пятью ведущими поставщиками образовательных услуг, включая Sylvan Learning и Code Ninjas, программа насчитывает более 1800 учебных центров по всей стране, где дети в возрасте от 6 до 12 лет получают наставников, которые могут помочь им в учебе.
IBM также сотрудничает с Bright Horizons, предлагая сотрудникам до 25 дней субсидируемого резервного ухода в год либо в центре, либо на дому, сообщил представитель CNBC Make It. Кроме того, компания недавно обновила политику оплачиваемых отпусков, предоставив родителям и опекунам до 4 недель гибкого экстренного отпуска.Сотрудники могут использовать это время по своему усмотрению, в том числе двухчасовыми сегментами, полдня или полдня или четыре недели подряд.
Что касается присмотра и дополнительного ухода за детьми младшего возраста, в Bright Horizons в настоящее время открыто около 725 центров, и осенью планируется открыть больше. Обычно компания управляет почти 1100 центрами по уходу за детьми по всему миру.
Но эти льготы далеки от стандартных для большинства работающих родителей, которые этой осенью совмещают работу и дистанционное обучение.Опрос Catalyst показал, что в целом 46% родителей говорят, что либо их работодатель не имеет планов помощи в уходе за детьми, либо они не были проинформированы об этом.
И женщины чаще сообщают об отсутствии поддержки: 49% опрошенных матерей заявили, что они не знают о том, что их работодатель предоставляет какие-либо пособия по уходу за ребенком, по сравнению с 39% опрошенных мужчин.
Недостаточно просто дать всем больше свободного времени, говорит Никель. Работа все еще ждет вас, когда вы снова войдете в систему.Вместо этого компании должны спрашивать сотрудников, чем они могут помочь. И это будет выглядеть по-разному для всех, говорит она.
Родители беспокоятся о долгосрочных последствиях для своей карьеры
Даже если у вас есть поддерживающий работодатель, родители все еще чувствуют давление. Виггс, как и многие родители, беспокоится о возможных долгосрочных последствиях для своей карьеры.
Родители говорят, что больше не могут отделять семейную жизнь и обязанности от работы, особенно когда дети прерывают видеовстречи или их слышно на фоне телефонных звонков.«Я занимаюсь связями с общественностью, коммуникациями и маркетингом уже 15 лет, и шесть из них я провела в роли матери-одиночки. И я держала это в тайне», — говорит Никель, опасаясь, что это испортит мнение работодателей. представление о ее способности выполнять работу. Но теперь этого не скрыть, говорит она.
Стремление стать родителем беспокоит Виггса. «Я нахожусь в той точке своей карьеры, когда пытаюсь перейти на следующий уровень и подготовиться к продвижению по службе», — говорит Виггс. «Несмотря на то, что все говорят, что все в порядке, мы не ожидаем от вас многого прямо сейчас, это беспрецедентно, я знаю, что все еще хочу показать себя как можно лучше, потому что я все еще хочу, чтобы моя карьера продолжала развиваться.»
Виггс говорит, что это похоже на декретный отпуск. Независимо от того, насколько щедрой является политика в отношении декретного отпуска, в глубине вашего сознания есть мысль, что, ну, технически, это не должно замедлять мой карьерный рост. , — говорит она. И тем не менее, вы видите, как ваши коллеги-мужчины получают повышение, пока вы в отпуске, и вас, вероятно, не рассматривают, потому что вас там нет.
это сумасшедшее время, и это не должно влиять на карьерный рост людей», — говорит Виггс.«Но я также знаю, что кто-то, у кого нет детей, может выступать на более высоком уровне, чем я сейчас».
Не пропустите больше из этой серии:
Frontiers | Устойчивая синаптическая патофизиология гиппокампа после одиночных и повторных ударов сотрясением головы
Введение
Черепно-мозговая травма (ЧМТ) является ведущей причиной смерти и инвалидности в США, от которой ежегодно страдают примерно 2,8 миллиона человек, что приводит к значительным финансовым и человеческим жертвам (Taylor et al., 2017). Многочисленные патологоанатомические исследования выявили корреляцию между повторными сотрясениями мозга и хронической травматической энцефалопатией (ХТЭ), нейродегенеративным состоянием, характеризующимся когнитивными нарушениями, перепадами настроения и депрессией, и нарушением работы уязвимых областей мозга, участвующих в обработке памяти, таких как кора и гиппокамп (McKee et al. ., 2009, 2013; Макки и Робинсон, 2014). В то время как многие громкие сообщения о ЧМТ связаны с военными и спортивными событиями, большая часть населения страдает от сотрясений мозга, возникающих в основном в результате падений и насилия (CDC, 2014), с однократными или повторными сотрясениями мозга, приводящими к кратковременной потере памяти. (Майер и др., 2017) или других симптомов нейродегенеративного заболевания (Fakhran et al., 2013). Действительно, ХТЭ, связанный с ЧМТ, имеет многие из тех же патофизиологических признаков, что и болезнь Альцгеймера, включая накопление гиперфосфорилированного тау, белка, ассоциированного с микротрубочками, который участвует в функции цитоскелета (Geddes et al., 1999; McKee and Robinson, 2014). Долгосрочные последствия ЧМТ также включают повышенную вероятность судорог (Annegers et al., 1998) и потерю равновесия/аномальную походку (Ellemberg et al., 2009), что еще больше увеличивает вероятность последующих ЧМТ (Theadom et al. ., 2015). Хотя долговременная физиология, лежащая в основе сотрясения мозга, недостаточно изучена, считается, что острые последствия сотрясения мозга возникают в результате нескольких аномальных клеточных процессов, происходящих после сотрясения мозга. При воздействии происходит быстрое увеличение внеклеточного глутамата (Hinzman et al., 2010), что приводит к гиперактивации постсинаптических каналов Ca 2+ , включая рецепторы NMDA (NMDAR; Geddes et al., 2003; Biegon et al. , 2004) и потенциалзависимые каналы Ca 2+ (VGCCs; Wolf et al., 2001). Последствия повышенного содержания нейронов Ca 2+ могут сохраняться в течение часов или даже дней после удара (Sun et al., 2008) и часто сопровождаются той или иной формой когнитивных нарушений (Deshpande et al., 2008). Устойчивое изменение гомеостаза Са 2+ является центральной особенностью нескольких неврологических заболеваний, включая болезни Альцгеймера и Хантингтона, и может служить общим основным механизмом, связывающим повышенную заболеваемость деменцией в годы после ЧМТ (Giacomello et al., 2013; Шранк и др., 2020).
Ca 2+ является жизненно важным медиатором нормальной клеточной и синаптической функции. Нейрональный Ca 2+ жестко регулируется с помощью нескольких гомеостатических механизмов, при этом нарушение этих процессов способствует нарушению синаптической регуляции, изменению сигнальных каскадов и потенциальному повреждению или гибели клеток (Stutzmann, 2007; Chakroborty et al., 2009, 2019; Zhang et al. ., 2015). Постсинаптические рианодиновые рецепторы (RyR), представляющие собой каналы Ca 2+ с высокой проводимостью на мембране эндоплазматического ретикулума (ER), расположены рядом с NMDAR и VGCC на дендритных шипиках пирамидных клеток гиппокампа (Jaffe et al., 1994; Чавис и др., 1996; Тибо и др., 2007 г.; Холбро и др., 2009). Эта взаимосвязь позволяет усилить опосредованный NMDAR и VGCC приток Ca 2+ в процессе индуцированного Ca 2+ высвобождения Ca 2+ (CICR; Emptage et al., 1999; Borde et al., 2000; Holbro et al., 2009), и, как таковые, RyRs способны влиять на Ca 2+ -зависимые процессы синаптики и пластичности (Chakroborty et al., 2009, 2019). Кроме того, экспрессия RyR повышена в посмертной ткани головного мозга у людей с когнитивными нарушениями, что является признаком ЧМТ (Bruno et al., 2012; Макиннес и др., 2019). Чувствительность RyR изменяется в результате процессов фосфорилирования и окисления, которые увеличивают вероятность открытия канала, что приводит к спонтанному «протеканию» каналов и повышенному высвобождению Ca 2+ в краткосрочной перспективе, но снижает содержание депо ER и подавляет ER Ca 2+ высвобождение, если оно будет устойчивым (Liu et al., 2012; Lacampagne et al., 2017). Хотя роль RyR в краткосрочных эффектах ЧМТ изучена недостаточно, ЧМТ резко приводит к увеличению продукции активных форм кислорода (АФК), которые являются известными медиаторами гиперфосфорилирования/окисления RyR (Görlach et al., 2015). Поскольку RyR также экспрессируются пресинаптически, повышенная пресинаптическая чувствительность RyR/Ca 2+ может привести к повышенному высвобождению возбуждающего нейромедиатора глутамата, что приводит к прямому усилению постсинаптического NMDAR и RyR-опосредованному высвобождению Ca 2+ . Примечательно, что стабилизация вызванного RyR высвобождения Ca 2+ приводит к широкому спектру терапевтических эффектов, включая снижение эксайтотоксичности глутамата (Frandsen and Schousboe, 1991; Niebauer and Gruenthal, 1999), нормализацию синаптической передачи и синаптической структуры, снижение маркеров воспаления, и улучшение поведенческих результатов (Chakroborty et al., 2012; Оулес и др., 2012 г.; Бриггс и др., 2017). Таким образом, RyR могут выступать в качестве терапевтической мишени при лечении ЧМТ и родственных заболеваний.
TBI вызывает несколько патологических признаков, общих для других неврологических заболеваний, включая аномальные агрегаты белков и гиперфосфорилированный тау. В нормальных условиях гомеостаз фосфорилирования тау-белка поддерживается за счет баланса активности киназы и фосфатазы, ответственных за его фосфорилирование и дефосфорилирование соответственно (Geddes et al., 1999; Макки и Робинсон, 2014 г.). После ЧМТ активируется несколько киназ, что приводит к чистому увеличению фосфорилированного тау (Tran et al., 2012). Кроме того, повышенное содержание Са 2+ в нейронах после ЧМТ, через NMDAR, VGCC и внутриклеточные запасы может активировать специфические Са 2+ -регулируемые киназы, которые фосфорилируют тау, такие как GSK3-β и Cdk5 (Avila et al., 2004; Dash et al., 2011; Zhao et al., 2012; Wilson et al., 2014). В свою очередь, фосфорилированный тау может увеличивать внутриклеточный Ca 2+ , способствуя фосфорилированию тау (Gómez-Ramos et al., 2006; Stutzmann, 2007) и связанные с Ca 2+ синаптические дефициты. В то время как острые эксайтотоксические явления Ca 2+ были описаны в течение нескольких минут или часов после ЧМТ (Luo et al., 2011; Gurkoff et al., 2013; Arai et al., 2019), устойчивые внутриклеточные Ca 2+ дисгомеостаз, такой как наблюдаемый при БА (Stutzmann, 2007), также может возникать и лежать в основе когнитивных, гистопатологических и синаптических дефектов, которые могут возникать через недели или месяцы после травмы (Deshpande et al., 2008; Sun et al., 2008).
Предыдущая травма головы является значительным фактором риска заболеваний, связанных с деменцией, при этом время от момента травмы до появления симптомов, подобных деменции, составляет от нескольких месяцев до нескольких лет (Fleminger et al., 2003; Li et al., 2017). RyR и VGCC играют важную роль в гомеостазе Ca 2+ , синаптической передаче и кодировании памяти, несмотря на задокументированную роль нарушения регуляции Ca 2+ при нейродегенеративных заболеваниях (Huang and Malenka, 1993; Huber et al., 1995; Чакроборти и др., 2012; Oules et al., 2012), их вклад в устойчивые клеточные и синаптические дефекты, возникающие в результате ЧМТ, изучен недостаточно. Здесь мы исследуем режимы устойчивой патофизиологии, возникающие в результате однократной или повторной ЧМТ на клинически значимой модели крысы (Jamnia et al., 2017), и выявляем ключевые клеточные сигналы, дефекты синаптических цепей и гистопатологические маркеры, которые согласуются с состояниями хронических неврологических заболеваний. .
Материалы и методы
Хронология экспериментальной процедуры
Приблизительно через 1 неделю после прибытия животных подвергали ложной хирургии или однократному или повторному контролируемому воздействию на кору головного мозга (CCI).Повторные ЧКИ проводились с использованием трех последовательных воздействий, разделенных 48-часовыми интервалами. Крыс исследовали через 30 дней после последней CCI для измерения степени устойчивых синаптических и клеточных эффектов; см. изображение ниже. Электрофизиологические/двухфотонные записи и окрашивание фосфотау проводили с использованием отдельных групп животных.
Животные
Самца крыс Long-Evans с капюшоном (Лаборатория Чарльза Ривера; 200–300 г; P60-P80) содержали по два в клетке в Центре биологических ресурсов Университета медицины и науки им. Розалинды Франклин (RFUMS).Хотя мы признаем важность пола как биологической переменной, ограниченный масштаб исследования, наряду с гораздо более высокой частотой ЧМТ у самцов (CDC, 2014), означает, что в этом исследовании мы использовали только самцов крыс. Крыс содержали на цикле свет/темнота 12:12 с пищей и водой вволю . Животных обрабатывали ежедневно в течение примерно 1 недели до операции. Все эксперименты проводились в соответствии с Руководством Национального института здравоохранения по уходу и использованию животных и были одобрены Институциональным комитетом по уходу и использованию животных RFUMS.
Контролируемое воздействие на кору с закрытой головой (CCI)
Чтобы нанести закрытую ЧМТ, мы использовали модифицированный подход CCI (Leica Impact One, Leica Microsystems Inc., Buffalo Grove, IL, USA) для моделирования легкой ЧМТ (Jamnia et al., 2017). Крыс анестезировали изофлюрановой анестезией со скоростью 2,0–3,0 мл/мин и помещали в стереотаксический аппарат Копфа (Kopf, Tujunga, CA, USA) на пенопластовую подушку (толщиной 5 см) в рамке из плексигласа; анестезию поддерживали через носовой конус. Рама состояла из основания (11.43 × 24,13 см) и боковую часть (9,52 × 22,86 см), расположенную под углом 11 градусов к вертикали. Чтобы обеспечить движение головы, а также обеспечить устойчивость крысы во время удара, боковая поверхность головы слегка прилегала к рамке из плексигласа. Стереотаксическое устройство Impact One наносило кортикальный удар под углом 20 градусов от вертикали, что позволяло плоскому кончику импактора располагаться перпендикулярно поверхности головы; при необходимости это было скорректировано соответствующим образом. Все повреждения производились плоским кончиком 5 мм на 6.5 м/с на глубине 10 мм от поверхности кожи, покрывающей правую сенсомоторную кору, в течение 300 мс. Измерение глубины учитывало степень поглощения пены (приблизительно 9 мм). Ранее эта глубина была протестирована как максимальный уровень удара, который не привел к перелому черепа, но вызвал нарушение поведения (Jamnia et al., 2017). Все крысы, подвергшиеся воздействию, получали местный анальгетик, а температуру тела поддерживали на уровне 37°С в период выздоровления. После выхода из наркоза крыс возвращали в их домашние клетки и ежедневно контролировали.У животных с повторным сотрясением мозга было всего три травмы с интервалом в 48 часов; дополнительную группу крыс с однократным сотрясением мозга подвергали двум последовательным анестезиям с интервалом в 48 часов. Эта группа была помещена в анестезиологическую камеру на время, в течение которого животных с повторным сотрясением мозга анестезировали для их последующих травм (примерно 20 мин). Эта модель однократной или повторной ЧМТ, использованная нами ранее, не приводит к какому-либо явному повреждению тканей (Jamnia et al., 2017) и считается моделью однократной или повторной легкой ЧМТ или сотрясения головного мозга (Petraglia et al., 2014).
Подготовка срезов мозга
Через 30 дней после последней CCI животных глубоко анестезировали изофлураном и коронарные срезы гиппокампа (400 мкм для регистрации в полевых условиях и 250 мкм для регистрации целых клеток и визуализации Ca 2+ ) готовили, как описано ранее (Stutzmann et al. , 2004). Срезы перфузировали со скоростью 1,5–2 мл/мин стандартным раствором искусственной спинномозговой жидкости (аЦСЖ), содержащим (в мМ): 125 NaCl, 2,5 KCl, 2 CaCl 2 , 1.2 MgSO 4 , 1,25 NaH 2 PO 4 , 25,0 NaHCO 3 , 10 D-декстроза, уравновешенная с 95% O 2 – 5% CO 2 –5% CO , температура (22°C). Осмолярность поддерживали на уровне 310 мОсм. Патч-пипетки (4–5 МОм) заполняли внутриклеточным раствором, содержащим следующие вещества (в мМ): 135 K-глюконат, 10 HEPES, 10 Na-фосфокреатин, 2 MgCl 2 , 4 NaATP и 0,4 NaGTP, pH отрегулирован. до 7,3–7,4 с КОН (Sigma). Пирамидные нейроны гиппокампа CA1 были идентифицированы визуально с помощью инфракрасной дифференциально-интерференционной контрастной оптики (IRDIC) на вертикальном микроскопе Olympus BX51 через объектив 40× и идентифицированы электрофизиологически по их пассивным свойствам мембраны и частотной адаптации спайков в ответ на инъекцию деполяризующего тока. .Мембранные потенциалы были получены в режиме фиксации тока на частоте 10 кГц с помощью аналого-цифрового преобразователя Digidata 1322 и усилителя Multiclamp 700B, а затем были записаны и проанализированы с использованием программного обеспечения pClamp 10.2 (Molecular Devices).
Записи потенциала внеклеточного поля
Для регистрации потенциала внеклеточного поля срезы гиппокампа размером 400 мкм переносили в интерфейсную камеру (Harvard Apparatus), перфузировали оксигенированным aCSF (1,5 мл/мин) при комнатной температуре и обрабатывали непрерывным потоком увлажненного газа (95% O 2 /5% CO 2 ).Данные были получены на частоте 10 кГц с использованием программного обеспечения pClamp 10.2 с усилителем AxoClamp 2B и платой DigiData 1322A для оцифровки (Molecular Devices). Полевые возбуждающие постсинаптические потенциалы (пВПСП) регистрировали в лучистом слое субполя СА1 гиппокампа с помощью регистрирующих микроэлектродов (2–6 МОм), заполненных аЦСЖ. Микроэлектроды вытягивали из боросиликатных стеклянных капилляров (Harvard Apparatus) на съемнике для пипеток P-2000 (Sutter Instruments, Novato, CA, USA). Синаптические ответы пВПСП вызывали стимуляцией коллатерального/комиссурального пути Шаффера с использованием биполярного стимулирующего электрода, при этом наклон пВПСП рассчитывали как изменение потенциала (ΔV) исходной формы волны пВПСП с течением времени (t) или ΔV/t ( мВ/мс, см. рисунок 1А).Кривые наклона вход-стимул/выход-fEPSP (I-O) были созданы путем построения графика зависимости наклона fEPSP от интенсивности стимула с использованием интенсивности стимула в диапазоне от 100 до 1000 мкА. Для индукции долговременной потенциации (ДП) исходные пВПСП вызывали при 50% от максимального наклона пВПСП при частоте 0,05 Гц в течение 20 мин. LTP индуцировали с исходной интенсивностью с использованием протокола высокочастотной стимуляции (HFS), состоящего из двух наборов по 100 импульсов с частотой 100 Гц, с интервалом между каждым набором импульсов 10 сапартов. пВПСП регистрировали еще в течение 60 мин с базовой частотой после ВЧС.Сразу после ВЧС мы наблюдали посттетаническую потенциацию (ПТП; 0–2 мин после ВЧС), Ca 2+ -зависимую форму кратковременной синаптической пластичности (Zucker and Regehr, 2002). ДП наблюдалась позже и может быть разделена на транзиторную раннюю фазу, опосредованную модификацией ранее существовавших белков (Е-ДП; 15–20 мин после ВЧС), и более стойкую позднюю фазу, требующую транскрипции генов и синтеза нового белка. (L-LTP; 50–60 мин после ВЧС; Davies et al., 1989; Frey et al., 1993).Усиление парных импульсов (PPF) оценивали после записи I-O с использованием межстимульного интервала 50 мс. Были записаны три последовательные пары ответов с частотой 0,05 Гц. Все полевые записи проводились на стороне гиппокампа, ипсилатеральной к месту удара. Для каждого среза проводили только один набор полевых записей, обычно с использованием двух срезов гиппокампа толщиной 400 мкм на одну крысу.
Рисунок 1 . Однократная или повторная черепно-мозговая травма (ЧМТ) приводит к стойкой повышенной возбудимости в синапсах СА3-СА1.Крысы, подвергшиеся однократной ЧМТ (sTBI) или повторной TBI (rTBI), демонстрировали стойкую активацию в синапсах CA3-CA1, что было выявлено с помощью записей внеклеточного поля (fEPSP). (A1) Кривые вход-выход для пВПСП, полученные из записей полей гиппокампа СА1 с использованием коллатеральной стимуляции Шаффера, через 30 дней после ложной хирургии, ЧМТ или ЧМТ, демонстрирующие возрастание наклона потенциала поля (вВПСП) в результате увеличения интенсивности стимула в диапазоне от 100 мкА до 1000 мкА. ФВПСП как в группах с ЧМТ, так и в группах с ЧМТ показали более крутой наклон при большинстве интенсивностей стимула по сравнению с группой с фиктивным лечением, без существенной разницы между группами с ЧМТ и сЧМТ. (A2) Репрезентативные входные-выходные следы fEPSP, взятые у крыс с имитацией, sTBI и rTBI. (B1) sTBI и rTBI не влияли на соотношение парных импульсов fEPSP по сравнению с имитацией, что указывает на отсутствие существенной разницы в вероятности пресинаптического высвобождения глутамата между группами. (B2) Репрезентативные кривые, показывающие парные пВПСП, вызванные с межстимульным интервалом 50 мс. Все записи были на стороне гиппокампа, ипсилатеральной к месту удара ЧМТ (над правой сенсомоторной корой). # p < 0,05 для имитации по сравнению с ЧМТ. * p < 0,05 для имитации по сравнению с rTBI. Столбики погрешностей представляют собой стандартную ошибку среднего.
Ca
2+ Визуализация
Визуализация Ca 2+ в пределах отдельных пирамидных нейронов СА1 выполнялась в срезах гиппокампа с использованием специально созданной системы многофотонной визуализации с видеочастотой на основе вертикального корпуса микроскопа Olympus BX51 и импульсного ИК-лазера Ti:Saph (Spectra-Physics, Санта-Клаус). Клара, Калифорния, США; Stutzmann and Parker, 2005).Отдельные нейроны заполняли индикаторной фура-2 Ca 2+ (50 мкМ; ThermoFisher) через пипетку , как описано ранее (Stutzmann et al., 2004). Возбуждение лазера осуществлялось цугами (80 МГц) импульсов ~100 кадров/с на длине волны 780 нм. На этой длине волны возбуждение фура-2 генерирует яркий сигнал на низких уровнях Ca 2+ , что позволяет нам видеть небольшие компартменты, такие как шипы, на уровнях покоя; флуоресценция пропорционально снижается с увеличением уровней Ca 2+ , что позволяет проводить относительные измерения изменения по сравнению с исходным уровнем.Пучок фемтосекундного импульсного лазера сканировался резонансным гальванометром (General Scanning Lumonics), обеспечивающим быстрое (7,9 кГц) двунаправленное сканирование по оси x , и обычным линейным гальванометром по оси y , чтобы обеспечить полное -скорость сканирования кадров 30 кадров/сек. Лазерный луч фокусировался на ткани через 40-кратный иммерсионный объектив (числовая апертура = 0,8). Излучаемый флуоресцентный свет регистрировался фотоумножителем с широким полем (электронные трубки) для получения видеосигнала, который был захвачен и проанализирован с помощью Video Savant 5.0 (IO Industries; Лондон, Онтарио). Дальнейший анализ изображений с коррекцией фона был выполнен в автономном режиме с использованием программного обеспечения MetaMorph. Для количественной оценки покоя и изменений флуоресценции фура-2 мы выбрали область сомы пирамидальных клеток СА1 (окружающую, но исключая ядро), чтобы получить показатель цитозольного Ca 2+ (Stutzmann et al., 2003, 2004). ). Для вычитания фона была выбрана соседняя нефлуоресцентная область. Поскольку фура-2 показывает снижение флуоресценции с увеличением Ca 2+ , при использовании двухфотонного возбуждения при 780 нм сниженные базальные значения F 0 представляют собой повышенные базальные значения Ca 2+ .Относительные процентные изменения Ca 2+ рассчитывали как процентное изменение интенсивности флуоресценции по сравнению с исходным уровнем: ( F 0 /Δ F — 1) × 100 со значениями для F 0 и процентным изменением усредненные по ячейкам из каждой группы TBI. После наклеивания фура-2 давали уравновеситься в клетке в течение 10–15 мин, а после достижения флуоресцентного сигнала в клетку вводили положительный ток в режиме фиксации тока, что приводило к деполяризации клеточной мембраны и запуск активации потенциалзависимых каналов Ca 2+ (VGCC).Чтобы обеспечить согласованность между вызванными реакциями внутри и между группами, протокол деполяризации проводился сначала, до воздействия какого-либо препарата. Измерения Ca 2+ были получены от пика вызванного ответа. Применение ванны с кофеином (10 мМ) использовалось для запуска высвобождения RyR-Ca 2+ из запасов ЭР (Гаращук и др., 1997; Сандлер и Барбара, 1999). Каждый срез мозга подвергался воздействию только одного применения кофеина. Как правило, в любой день записи два среза гиппокампа использовались для измерений Ca 2+ с записью одной пирамидной клетки CA1 на срез.Минианализ 6.0.7 (Synaptosoft; Форт-Ли, Нью-Джерси, США) использовался для обнаружения и измерения событий спонтанного возбуждающего постсинаптического потенциала (сВПСП) с минимальной амплитудой 0,2 мВ и минимальной площадью 3 мВ×мс. Базовый уровень определялся по среднему значению за 1 мс непосредственно перед каждым событием или, в случае перекрывающихся событий, по кинетике пика и затухания предыдущего события с использованием «обнаружения сложного пика» в минианализе. 6-циано-7-нитрохиноксалин-2,3-дион (CNQX, Sigma-Aldrich), антагонист AMPA, использовали для подтверждения глутаматергических sEPSP.Данные sEPSP были получены только в группах симуляции и rTBI, поскольку мы наблюдали только влияние rTBI, но не sTBI, на соматический ответ RyR-Ca 2+ на кофеин (см. Рисунок 4). Все записи целых клеток/двух фотонов проводились на стороне гиппокампа, ипсилатеральной к месту удара. Несколько записей VGCC проводили на срез, но только один ответ RyR-Ca 2+ на кофеин на срез, обычно с 2–3 срезами гиппокампа размером 250 мкм на используемую крысу.
Рисунок 2 .Острая индукция синаптической пластичности гиппокампа не зависит от однократной или повторной ЧМТ. (A) Высокочастотная стимуляция (ВЧС) коллатералей Шаффера СА3-СА1 приводила к кратковременной посттетанической синаптической потенциации (ПТП), продолжавшейся несколько минут. На графике показана усредненная временная динамика ПТП в группах ложной, однократной ЧМТ (сЧМТ) и повторной ЧМТ (рЧМТ). (B) Высокочастотная стимуляция (ВЧС) коллатералей Шаффера СА3-СА1 также приводила к длительной потенциации (ДП) продолжительностью 1 час.График показывает усредненную временную динамику ранней (E-LTP) и поздней фазы LTP (L-LTP) в группах имитации, сЧМТ и rTBI. Время ВЧС указано стрелкой. (C) Гистограмма показывает процентное изменение ответа после HFS по сравнению с исходным уровнем в виде PTP (0–2 мин после HFS), E-LTP (15–20 мин после HFS) и L-LTP (50–60 мин). мин после ВЧС). ВЧС состоял из двух серий импульсов частотой 100 Гц, длительностью 1 с каждая, с интервалом между сериями импульсов 10 с. Не было значительного влияния sTBI или rTBI на величину PTP, E-LTP или L-LTP.Наложенные линии тренда представляют средние данные для имитации (синий), sTBI (оранжевый) и rTBI (красный). Все записи были на стороне гиппокампа, ипсилатеральной к месту удара ЧМТ (над правой сенсомоторной корой). Столбики погрешностей представляют собой стандартную ошибку среднего.
Рисунок 3 . ЧМТ приводит к повышенной активности потенциалзависимых каналов Ca 2+ в пирамидных нейронах CA1 гиппокампа. (A) Слева, гистограмма, суммирующая пик Ca 2+ эффектов активации потенциалзависимых каналов Ca 2+ (VGCC) (с использованием инъекции деполяризующего тока в пирамидные нейроны СА1) через 30 дней после ложной процедуры, однократная ЧМТ (sTBI) или повторная TBI (rTBI).Пиковые эффекты VGCC Ca 2+ были больше в группах с ЧМТ (серая полоса) и rTBI (черная полоса) по сравнению с имитацией (белая полоса). Количество записанных ячеек в группе указано на каждой полосе. Справа: последовательность потенциалов действия в ответ на инъекцию деполяризующего тока в пирамидный нейрон СА1 (характеристики потенциала действия не изменились ни при sTBI, ни при rTBI, см. Таблицу 1). (B) Образцы следов, иллюстрирующие эффекты активации VGCC Ca 2+ в отдельных ложных (слева), sTBI (в центре) или rTBI (справа) пирамидных нейронов CA1.Калибровочные столбцы иллюстрируют изменение % Ca 2+ по сравнению с исходным уровнем. (C) Флуоресцентные (монохромные) и псевдоцветные изображения нагруженных фура-2 пирамидных нейронов СА1. Псевдоцветные изображения иллюстрируют относительные ответы Ca 2+ на активацию VGCC в группах имитации (слева), sTBI (в центре) и rTBI (справа). * р < 0,05. Все записи были на стороне гиппокампа, ипсилатеральной к месту удара ЧМТ (над правой сенсомоторной корой). Столбики погрешностей представляют собой стандартную ошибку среднего.
Рисунок 4 . TBI приводит к увеличению покоящегося Ca 2+ и притуплению вызванного RyR высвобождения Ca 2+ в пирамидных нейронах CA1 гиппокампа. (A) Слева, столбчатая диаграмма, суммирующая усредненные значения флуоресценции Ca 2+ на основе в покое ( F 0 ) в пирамидных нейронах СА1 через 30 дней после имитации процедуры (белая полоса), одиночной ЧМТ (sTBI, серый) бар) или повторная ЧМТ (рЧМТ, черная полоса). Пониженные значения флуоресценции в покое ( F 0 ) в обеих группах ЧМТ по сравнению с ложной группой указывают на повышенный уровень Ca 2+ в покое.Справа гистограмма, показывающая, что усредненный ответ RyR-Ca 2+ на кофеин (20 мМ) был снижен в группе rTBI (черная полоса), но не в группе sTBI (серая полоса), по сравнению с симуляцией (белая полоса). . Количество записанных ячеек в группе указано на каждой полосе. (B) Образцы следов, иллюстрирующие RyR-Ca 2+ эффекты кофеина (20 мМ) в отдельных ложных (слева), sTBI (в центре) и rTBI (справа) пирамидных нейронах CA1. Калибровочные столбцы иллюстрируют изменение % Ca 2+ по сравнению с исходным уровнем. (C) Флуоресцентные (монохромные) и псевдоцветные изображения нагруженных фура-2 пирамидных нейронов СА1. Псевдоцветные изображения иллюстрируют относительные эффекты RyR-Ca 2+ кофеина в группах имитации (слева), sTBI (в центре) и rTBI (справа). * р < 0,05. Все записи были на стороне гиппокампа, ипсилатеральной к месту удара ЧМТ (над правой сенсомоторной корой). Столбики погрешностей представляют собой стандартную ошибку среднего.
Иммуногистохимия
Крыс анестезировали пентобарбиталом (200 мг/кг), а затем транскардиально перфузировали ледяным фосфатно-солевым буфером, а затем сразу же вводили 4% параформальдегид.Мозг удаляли и постфиксировали в течение 24 ч, затем разрезали на сагиттальные срезы на микротоме Leica SM2000R со столиком для замораживания толщиной 40 мкм и хранили в растворе для криоконсервации до иммуноокрашивания. Свободно плавающие срезы гиппокампа пермеабилизировали с помощью 0,1 М TBS 1,0% Triton-X (3 × 10 мин) при комнатной температуре на платформе-качалке. Срезы блокировали 0,1 М TBS 0,1% Triton-X + 10% козьей сыворотки в течение 1 ч при комнатной температуре, а затем инкубировали с первичными антителами (фосфо-Тау S262, 1:500, Invitrogen, каталожный номер 44-750G), разведенными в .1 М TBS 0,1% Triton-X + 1% козья сыворотка в течение 24 ч при 4°C на платформе-качалке. Срезы промывали 0,1 М TBS 0,1% Triton-X (3 х 5 мин) и инкубировали во вторичном антителе (Alexa Fluor 488, конъюгированном с козьим антикроличьим антителом IgG, 1:1000, Invitrogen, A-11008), разведенном в 0,1 M TBS 0,1% Triton-X + 1% козья сыворотка в течение одного часа при комнатной температуре на платформе-качалке. Срезы промывали в 0,1 М TBS 0,1% Triton-X (3 × 5 мин) и окрашивали 1:10 000 DAPI, разведенным в 0,1 М TBS, в течение 5 мин. Срезы промывали в 0.1 М TBS в течение 5 мин и смонтированы с PVA-DABCO для микроскопии. Контрольные срезы получали с использованием той же процедуры только с первичным антителом и только с вторичным антителом, соответственно, и визуализировали, чтобы убедиться в отсутствии нецелевого окрашивания. Конфокальные изображения срезов получали и анализировали с использованием 60-кратного объектива на конфокальном микроскопе Olympus Fluoview Fv10i. Настройки микроскопа были одинаковыми для всех изображений. Было получено несколько изображений каждой области гиппокампа, а также коры.Программное обеспечение MetaMorph (версия 7) использовалось для количественной оценки процентной плотности окрашивания флуоресцентно меченого фосфорилированного тау сверх порогового уровня интенсивности по всему мозгу, а также в каждом полушарии, ипсилатеральном и контралатеральном к повреждению.
Статистический анализ
Статистические различия между группами определяли с помощью дисперсионного анализа (однофакторный дисперсионный анализ или двусторонний дисперсионный анализ с повторными измерениями) с апостериорными тестами Тьюки или Даннетта , где это уместно, с использованием программного обеспечения для анализа данных SigmaPlot 12.Данные представлены как среднее ± стандартная ошибка среднего. Статистическая значимость была установлена на уровне p <0,05.
Результаты
В нашем предыдущем исследовании повторное сотрясение мозга/ЧМТ приводило к более стойким нарушениям памяти, чем однократное сотрясение мозга (Jamnia et al., 2017), с аналогичными результатами, наблюдаемыми в другом месте (Meehan et al., 2012; Weil et al., 2014). Дефицит памяти, связанный с сотрясением мозга, может быть результатом стойких дезадаптивных патофизиологических эффектов в областях мозга, которые играют роль в обучении и памяти, таких как гиппокамп.С этой целью мы проверили наличие стойкого дефицита передачи сигналов Ca 2+ в гиппокампе и нейронов, а также тау-патологию у крыс, подвергшихся однократной или повторной ЧМТ.
sTBI и rTBI приводят к гипервозбудимой синаптической передаче в цепях гиппокампа
Электрофизиологические записи в острых срезах гиппокампа головного мозга использовались для оценки изменений в локальной цепи и синаптической функции, возникающих в результате однократной или повторной ЧМТ. В общей сложности 7, 6 и 8 крыс были разделены на группы имитации, sTBI и rTBI соответственно.Записи потенциала внеклеточного поля измеряли эффекты стимуляции коллатералей Шаффера CA3-CA1 на постсинаптические потенциалы (fEPSP) с использованием функции начального наклона fEPSP в качестве меры моносинаптического выхода. Оценка функции ввода-вывода пВПСП показала, что как sTBI (11 записей/11 срезов/6 крыс), так и rTBI (18 записей/18 срезов/8 крыс) приводили к значительной потенциации в синапсах гиппокампа в диапазоне интенсивностей стимуляции. по сравнению с фиктивной группой (18 записей/18 срезов/8 крыс).Двусторонний дисперсионный анализ с повторными измерениями был значимым для эффектов однократного и повторного лечения ЧМТ ( F (2,40) = 4,08; p = 0,024), интенсивности стимуляции ( F (9,40) ) = 89,08; p <0,001) и группа лечения по сравнению с взаимодействием стимуляция-интенсивность ( F (2,9) = 4,27; p <0,001). Апостериорный анализ выявил значительную разницу между группами с ЧМТ и плацебо при 500 мкА ( p = 0.049), 600 мкА ( p = 0,02), 700 мкА ( p = 0,004), 800 мкА ( p = 0,002), 900 мкА ( p = 0,09 p 0,004) и 1,0090 = 0,003) интенсивности стимуляции и между группами rTBI и имитации при 700 мкА ( p = 0,025), 800 мкА ( p < 0,005), 900 мкА ( p < 0,004) и 192 p 91 мкА ( < 0,004) <0,002) интенсивности стимуляции (рис. 1А). Чтобы определить, отражают ли повышенные синаптические ответы, наблюдаемые в группах sTBI и rTBI, пресинаптические изменения, мы использовали отношение парных импульсов, измеряемое как отношение наклона второго fEPSP к наклону первого fEPSP пары полученных fEPSP. с интервалом 50 мс, как относительная мера вероятности высвобождения Ca 2+ -зависимого нейротрансмиттера.Односторонний ANOVA не выявил влияния ни sTBI (11 записей/11 срезов/6 крыс), ни rTBI (12 записей/12 срезов/8 крыс) на соотношение парных импульсов ( F (2,34) = 1,44; p = 0,252; рисунок 1B) по сравнению с имитацией (14 записей/14 срезов/8 крыс), что указывает на то, что вероятность пресинаптически регулируемого высвобождения везикул существенно не различается между этими группами.
В дополнение к измерению базальной синаптической передачи мы также исследовали долговременную синаптическую потенциацию (ДП) в синапсах СА3-СА1.С помощью высокочастотной стимуляции (ВЧС, две последовательности импульсов по 100 Гц по 1 с, разделенные интервалом 10 с) коллатералей Шаффера мы наблюдали последовательность синаптических изменений, состоящую из начальной кратковременной посттетанической синаптической потенциации (ПТП; 0). –2 мин после ВЧС; рис. 2А), за которым следует ранняя ДП (Э-ДП; 15–20 мин после ВЧС) и поздняя фаза ДП (L-ДП; 50–60 мин после ВЧС; рис. 2Б). PTP не претерпел значительных изменений ни при однократной (sTBI; 8 записей/8 срезов/6 крыс), ни при повторной TBI (rTBI; 10 записей/10 срезов/8 крыс; односторонний ANOVA; F (2,24) = 2.34; p = 0,12) по сравнению с имитацией (7 записей/7 срезов/7 крыс), и также не было влияния sTBI или rTBI на величину E-LTP (односторонний ANOVA; F (2,24 ) = 0,57; p = 0,57) или L-LTP (односторонний ANOVA; F (2,24) = 0,89; p = 0,43; рис. 2C). Хотя наши результаты контрастируют с результатами других исследований, показывающих стойкое ослабление LTP гиппокампа в результате легочной ЧМТ (Schwarzbach et al., 2006) или rTBI (Aungst et al., 2014), в этих исследованиях использовалась модель удара с открытым черепом/обнаженной твердой мозговой оболочкой, а не модель с закрытой головой, используемая здесь. Действительно, хотя во многих исследованиях изучались синаптические эффекты гиппокампа в результате ЧМТ с использованием модели открытой/обнаженной твердой мозговой оболочки в различные моменты времени после травмы (Miyazaki et al., 1992; D’Ambrosio et al., 1998; Atkins, 2011). ; Zhang B. et al., 2011; Zhang B.-L. et al., 2011; Titus et al., 2019), в нескольких исследованиях изучалось стойкое влияние ЧМТ на синаптическую пластичность с использованием закрытой модели одиночных ( Уайт и др., 2017) или повторяющиеся воздействия (Logue et al., 2016). Кроме того, в других закрытых исследованиях использовалась модель повреждения взрывной волной (Beamer et al., 2016; Hernandez et al., 2018) с глобальными физическими эффектами, которые менее клинически значимы для легкой ЧМТ.
Ca
2+ Динамика и свойства мембран при sTBI и rTBI
Нейрональный Ca 2+ дисгомеостаз из-за изменений в функции VGCC и RyR может привести к изменению клеточной передачи сигналов и синаптической функции, а также вызвать потерю синапсов и нейродегенерацию, если она сохраняется с течением времени.Здесь мы оценили стойкие эффекты однократной или повторной ЧМТ на уровни покоя Ca 2+ , VGCC и функцию RyR. Использование записи патчей целых клеток с 2-фотонной визуализацией Ca 2+ позволяет одновременно измерять пассивные и активные свойства мембран, включая входное сопротивление и возбудимость клеток, а также изменения в обращении Ca 2+ во время VGCC и Активация RyR. Приток Ca 2+ через VGCC был значительно повышен как в группах с ЧМТ (16 записей/12 срезов/6 крыс), так и в группах с ЧМТ (19 записей/12 срезов/8 крыс) по сравнению с имитацией (10 записей/6 срезов/8 крыс). Ф (2,44) = 7.89; р = 0,001; Рисунок 3А). Базальное содержание Ca 2+ , измеренное с использованием интенсивности флуоресценции фура-2 в покое ( F 0 ) при одинаковой мощности лазера и условиях записи, также было значительно повышено в обеих sTBI (15 записей/15 срезов/6 крыс). ) и группы rTBI (18 записей/18 срезов/8 крыс) по сравнению с плацебо (8 записей/8 срезов/8 крыс; F (2,40) = 5,0; p = 0,012; рис. 4А). Напротив, сигналы Ca 2+ , вызванные RyR, были значительно снижены только в группе rTBI (18 записей/18 срезов/8 крыс) ( F (2,39) = 4.57; р = 0,017; Фигура 4А), но не sTBI (15 записей/15 срезов/6 крыс), по сравнению с имитацией (8 записей/8 срезов/8 крыс). Наши результаты согласуются с предыдущими исследованиями, демонстрирующими устойчивые патофизиологические эффекты, возникающие в результате ЧМТ, включая усиление кортикальной возбуждающей синаптической передачи (3–5 недель после травмы; Koenig et al., 2019) и дисгомеостаз Ca 2+ в нейронах гиппокампа через 30 дней. после травмы (Sun et al., 2008). Кроме того, в обоих этих исследованиях характер воздействия (открытая головка/обнаженная твердая мозговая оболочка) заметно отличался от модели с закрытой головкой, используемой в нашем исследовании, использовалось только одно воздействие и использовались данные визуализации Ca 2+ . получен из остро изолированных нейронов гиппокампа СА3, а не из пирамидных нейронов СА1 в срезе головного мозга.Таким образом, хотя это трудно экстраполировать непосредственно на эти исследования, существует явный прецедент синаптических дефектов и дисгомеостаза Ca 2+ , возникающих в результате ЧМТ в цепях мозга, важных для обучения, памяти и познания.
В дополнение к измерению соматического высвобождения Ca 2+ , вызванного RyR, мы также исследовали базальную спонтанную синаптическую передачу в форме возбуждающих постсинаптических потенциалов (sEPSPs) и то, как активация RyR по-разному влияет на частоту и амплитуду sEPSC в группах симуляции и rTBI. .Поскольку ответ RyR-Ca 2+ был притуплен только в условиях rTBI, мы аналогичным образом тестировали влияние кофеина на сВПСП в имитации (9 записей/9 срезов/8 крыс) и rTBI (17 записей/17 срезов/8 крыс). ) группы. До применения кофеина исходная частота сВПСП была одинаковой в обеих группах (двусторонний дисперсионный анализ с повторными измерениями; F (1,51) = 0,295; p = 0,837; рис. 5А), а частота sEPSPs был аналогичным образом увеличен кофеином (парный тест t ; t = -4.52; р < 0,001; Рисунок 5А). Перед активацией RyR исходная амплитуда сВПСП также была одинаковой как в группе плацебо, так и в группе с rTBI ( F (1,51) = 0,98; p = 0,47; рис. 5B), а амплитуда сВПСП была сходной. увеличивается под действием кофеина (парное t -тест; t = -8,14; p <0,05; рис. 5B).
Рисунок 5 . Повторная ЧМТ (rTBI) не влияет на спонтанные синаптические события и вклад активации RyR. (A) Частота исходных спонтанных возбуждающих постсинаптических потенциалов (sEPSPs; серые столбцы), измеренная в пирамидных нейронах CA1 гиппокампа, не была затронута rTBI через 30 дней после травмы. Нейроны sTBI не тестировались. Как в группе симуляции, так и в группе rTBI активация RyR кофеином (20 мМ; черные столбцы) одинаково увеличивала частоту сВПСП. (B) Исходная амплитуда sEPSP (серые столбцы) также не влияла на rTBI, но активация RyR кофеином (20 мМ; черные столбцы) приводила к значительному увеличению амплитуды sEPSP в группе rTBI. (C) Репрезентативные кривые, иллюстрирующие влияние применения ванны с кофеином (20 мМ) на частоту и амплитуду сВПСП во время регистрации токового зажима целой клетки в пирамидном нейроне CA1 из группы rTBI. ** р < 0,001, * р < 0,05. Все записи были на стороне гиппокампа, ипсилатеральной к месту удара ЧМТ (над правой сенсомоторной корой). Столбики погрешностей представляют собой стандартную ошибку среднего.
Повышенное окрашивание фосфорилированного тау после ЧМТ сохраняется по всей коре и гиппокампу
Уровень содержания фосфорилированного тау-белка был исследован в гиппокампе и вышележащей коре головного мозга в группах лечения с имитацией ЧМТ, ЧМТ и ЧМТ, ипсилатерально и контралатерально от места повреждения, с использованием иммуногистохимической маркировки и визуализации с помощью конфокальной микроскопии (рис. 6).Плотность каждого флуорофора выше фонового порогового уровня усреднялась по всем изображениям, полученным для данного животного с использованием программного обеспечения Metamorph (v.7). В гиппокампе этот анализ выявил повышенное окрашивание фосфорилированного тау в группе rTBI по сравнению с группой имитации (односторонний ANOVA; F (2,11) = 7,242; p ≤ 0,01) и группой sTBI ( р ≤ 0,05). В вышележащей сенсомоторной коре значительное увеличение фосфорилированного тау в группе rTBI по сравнению с группой имитации (односторонний ANOVA; F (2,11) = 8.245; p ≤ 0,01) и группу с ЧМТ ( p ≤ 0,05). Не было никаких существенных различий в фосфорилированном тау между ипсилатеральным и контралатеральным полушариями. Этот паттерн предполагает активный процесс, который генерирует фосфорилированный тау спустя много времени после первоначального повреждения, хотя возможны и другие объяснения, такие как нарушение клиренса видов патогенных белков. Кроме того, тау-патология не обязательно ограничивается местом или полушарием повреждения, но распространяется по полушариям головного мозга с течением времени независимо от первоначального места ЧМТ, при этом более высокая нагрузка фосфорилированного тау связана с повторяющейся травмой.
Рисунок 6 . Гиппокампальный и кортикальный фосфорилированный тау-белок повышается после однократной (sTBI) или повторной TBI (rTBI). (A–C) Репрезентативные конфокальные изображения (63×, в нескольких плоскостях) показывают иммуномечение фосфорилированного тау (pTau S262, зеленый) в области СА1 гиппокампа (слева) и слое V коры (справа) ипсилатеральнее травмы у ложных (A) , sTBI (B) и rTBI (C) животных. (D) Гистограммы показывают значительное увеличение количества фосфорилированного тау-белка в гиппокампе в группе rTBI (черные) по сравнению с sTBI (серые) и группами имитации (белые; n = 16 срезов/4 крысы для каждого условия лечения) . (E) Гистограммы показывают значительное увеличение содержания фосфорилированного тау в коре головного мозга в группе rTBI (черные) по сравнению с группами sTBI (серые) и имитации (белые; n = 16 срезов/4 крысы для каждого условия лечения) . Репрезентативные изображения были получены на конфокальном микроскопе Leica Microsystems SP8. Травма была над правой сенсомоторной корой. * p ≤ 0,05 и ** p ≤ 0,01. Столбики погрешностей представляют собой стандартную ошибку среднего.
Сводка данных
В целом, наше исследование выявило группу устойчивых патогенных или дезадаптивных признаков легкой ЧМТ, которые сохраняются долгое время после острых последствий первоначальной травмы.Эти особенности могут в совокупности влиять на системы кодирования памяти в гиппокампе и могут способствовать возникновению симптомов деменции, которые могут проявиться спустя долгое время после травмы. В частности, мы показываем, что sTBI и rTBI приводят к увеличению возбуждающего синаптического выхода наряду с увеличением активности Ca 2+ и VGCC в покое и снижением функции RyR в группе rTBI. Эта комбинация повышенной синаптической возбудимости и повышенной активности VGCC может играть роль в наблюдаемом повышенном базальном уровне Ca 2+ (графическое резюме см. на рис. 7).Мы также наблюдали повышенные и градуированные уровни фосфорилированного тау в группах с ЧМТ и сЧМТ, подтверждая идею связи между повышенным возбуждением нейронов и патологией тау.
Рисунок 7 . Схема влияния ЧМТ на синаптическую передачу гиппокампа и обработку Ca 2+ . Через 30 дней после однократной (sTBI) или повторной TBI (rTBI) мы наблюдали потенциацию вызванных fEPSP, указывающую на синаптическую потенциацию, возможно, в результате постсинаптических эффектов, включая усиление эффектов AMPA-рецепторов.Функция VGCC также была увеличена в группах с ЧМТ и rTBI, и это, в сочетании с усилением синаптической передачи, может играть роль в увеличении базального цитозольного Ca 2+ , наблюдаемого в обеих группах TBI. RyR-опосредованное высвобождение Ca 2+ было снижено, но только в группе rTBI, возможно, из-за истощения запасов Ca 2+ в эндоплазматическом ретикулуме (ER).
Обсуждение
Легкая ЧМТ или сотрясение головного мозга в последние годы привлекли к себе значительное внимание, отчасти отражая возросшее осознание их распространенности среди населения в целом и их роли в качестве значительного фактора риска для БА-подобного слабоумия и других неврологических заболеваний.Несмотря на это, в нескольких исследованиях изучались и сравнивались стойкие физиологические и синаптические последствия одиночных или повторяющихся сотрясений мозга с закрытой головой. Чтобы устранить пробелы в нашем понимании между травмой головного мозга и отсроченным когнитивным дефицитом, мы использовали модель сотрясения мозга с закрытой головой при однократной и повторной ЧМТ, которая вызывает дефицит памяти и работоспособности без явной потери клеток гиппокампа обнаруживаемого повреждения клеток (Jamnia et al., 2017). ). Одним из основных ограничений большинства исследований ЧМТ на сегодняшний день было сосредоточение внимания на измерениях повреждения тканей или гистопатологии в фиксированных срезах головного мозга, препарате, в котором не могут быть обнаружены активные изменения синаптической функции в реальном времени.Здесь мы впервые использовали потенциал внеклеточного поля и электрофизиологические записи целых клеток, чтобы идентифицировать клеточные и синаптические изменения, которые в противном случае могли бы остаться незамеченными в фиксированной ткани. На уровне локальных цепей в гиппокампе мы обнаружили стойкую активацию возбуждающей синаптической передачи в синапсах СА3-СА1, которая присутствовала как в группах однократной, так и повторной ЧМТ через 30 дней после травмы. Интересно, что эти фундаментальные изменения в синаптических ответах не приводили к изменению выражения долговременной синаптической пластичности; тем не менее, это не исключает основного дефицита LTP в этой модели травмы, поскольку было продемонстрировано задействование компенсаторных механизмов для поддержания нормального LTP в скомпрометированных сетях гиппокампа (Chakroborty et al., 2012). Также не было выявлено явных изменений в фасилитации парных импульсов, которая обычно считается показателем вероятности вызванного пресинаптического выброса (Regehr, 2012). Интересно, что rTBI также не влияла на частоту mEPSCs, измеренную с использованием записей целых клеток, подтверждая наблюдение, что TBI не влияет на вероятность пресинаптического высвобождения через несколько недель после травмы. Хотя это не исключает пресинаптического компонента из-за острого, но преходящего увеличения синаптического высвобождения глутамата после ЧМТ (Huang et al., 1992).
Поскольку наши данные показывают, что синаптическая передача в гиппокампе одинаково увеличивается как при sTBI, так и при rTBI, это говорит о том, что даже одного сотрясения достаточно, чтобы вызвать долгосрочные синаптические эффекты. Хотя точная серия клеточных событий, которые приводят к этому потенцированному синаптическому состоянию, все еще исследуется, наши данные предполагают, что потенцируется постсинаптический компонент fEPSP. Поскольку пВПСП обычно опосредуется активацией постсинаптических АМРА-рецепторов (Davies and Collingridge, 1989; Tsien et al., 1996), активация AMPA-рецепторов может играть роль в наблюдаемой синаптической потенциации. Кроме того, поскольку мембранный потенциал покоя пирамидных нейронов СА1 составлял примерно -70 мВ во всех группах лечения (таблица 1), маловероятно, что существует значительный компонент пВПСП NMDAR из-за блока Mg 2+ NMDAR в этой гиперполяризованной области. мембранный потенциал. Гиперчувствительность контура может также отражать снижение функции RyR и нижележащих нейромодуляторов, регулируемых Ca 2+ (Chakroborty et al., 2015), а также патологическое накопление тау (Huijbers et al., 2019). Уровни тау связаны с частотой приступов (DeVos et al., 2013), хорошо задокументированных последствий ЧМТ, которые также могут возникать в результате гиперактивности гиппокампа (Golarai et al., 2001). Другим возможным объяснением наблюдаемой повышенной возбудимости гиппокампа является снижение ингибирующей нейротрансмиссии гиппокампа CA1, еще одна особенность, характерная для животных моделей ЧМТ и болезни Альцгеймера (Reeves et al., 1997; Hazra et al., 2013; Алмейда-Сухетт и др., 2015).
Таблица 1 . Пассивные и активные свойства мембран не изменяются при однократной или повторной черепно-мозговой травме (ЧМТ).
Уменьшение высвобождения Ca 2+ , вызванное RyR, в группе rTBI сходно с зарегистрированным снижением ответа RyR-Ca 2+ в результате механического повреждения нейронов in vitro (Weber et al., 2002), эффект опосредованный NMDAR-опосредованным притоком Ca 2+ (Ahmed et al., 2002), последующее увеличение CICR и истощение запасов ER Ca 2+ (Weber et al., 2002). Действительно, повышенный базальный цитозольный Ca 2+ , возникающий в результате единичного события TBI, предотвращается предварительной обработкой антагонистом NMDAR MK801 (Deshpande et al., 2008). Хотя мы не наблюдали снижения эффектов RyR-Ca 2+ в группе с ЧМТ, вполне вероятно, что последствия повторного механического повреждения и повторного притока NMDAR Ca 2+ являются более стойкими, чем эффекты, возникающие в результате единичного повреждения (Slemmer et al. др., 2002; Weber, 2007), что приводит к долговременной модификации передачи сигналов RyR-Ca 2+ в результате rTBI. Примечательно, что устойчивый эффект rTBI на RyR-Ca 2+ коррелирует со стойкими когнитивными нарушениями, возникающими в результате rTBI, но не с TBI (Jamnia et al., 2017), что еще больше усиливает связь между дисфункцией RyR, дисгомеостазом Ca 2+ , и когнитивные нарушения после черепно-мозговой травмы (Deshpande et al., 2008; Sun et al., 2008; Bruno et al., 2012; Liu et al., 2012).
В то время как снижение RyR-Ca 2+ наблюдалось только в группе rTBI, повышенная активность VGCC наблюдалась в обеих группах лечения и может играть роль в повышенном базальном уровне Ca 2+ , присутствующем в обеих группах TBI. Устойчивая активация синаптической передачи гиппокампа, наблюдаемая в обеих группах ЧМТ, также может быть обусловлена повышенным притоком Ca 2+ через VGCC, и имеется достаточно доказательств того, что VGCC играют важную роль в синаптической потенциации гиппокампа (Huang and Malenka, 1993; Хубер и др., 1995). Поскольку не наблюдалось значительного влияния ни sTBI, ни rTBI на свойства пассивной или активной клеточной мембраны, измеренные через 30 дней после травмы (таблица 1), наблюдаемое усиление ответа VGCC нельзя объяснить изменениями сопротивления мембраны или порога постсинаптического потенциала действия.
Тау-патология является хорошо задокументированным признаком ЧМТ и связанных с ней состояний, таких как хроническая травматическая энцефалопатия (ХТЭ) и болезнь Альцгеймера (McKee et al., 2013; McKee and Robinson, 2014; Albayram et al., 2016), однако определенные механизмы, с помощью которых фосфорилирование тау сохраняется после повреждения, еще полностью не выяснены. В условиях ЧМТ равномерно распределенный патогенный тау, наблюдаемый через 30 дней после травмы, вероятно, является результатом каскада прямой связи, поскольку период полураспада растворимого тау составляет примерно 10 дней (Yamada et al., 2015). Возможным механизмом является повышенный уровень цитозольного Ca 2+ в состоянии покоя после ЧМТ, что может повышать активность GSK3β и Cdk5 и фосфорилировать тау по остаткам серина и треонина (Avila et al., 2004). Это, в свою очередь, может нарушить гомеостаз Ca 2+ и поддержать патогенный цикл (Gómez-Ramos et al., 2006). В соответствии с этим ингибиторы GSK3β продемонстрировали терапевтический потенциал для лечения ЧМТ помимо снижения фосфорилированного тау, предположительно, путем изменения передачи сигналов через рецепторные тирозинкиназы (RTK) и пути Wnt, эффекты, которые в противном случае привели бы к накоплению патологических белков, нейронов. потеря и когнитивная дисфункция (Shim and Stutzmann, 2016).Помимо изменения гомеостаза Ca 2+ , аномально фосфорилированный тау нарушает сборку микротрубочек и аксональный транспорт, что приводит к дефициту синапсов и, в конечном счете, к потере синапсов (Ial et al., 2005). Фосфорилированный тау также может быстро распространяться и накапливаться в областях мозга, удаленных от места повреждения после ЧМТ, но синаптически связанных с ним (Edwards et al., 2020). Дополнительные механизмы широко распространенной гистопатологии могут включать противосудорожное повреждение или генерализованное нейровоспаление (Edwards et al., 2020). Точные процессы, объясняющие распространение и распространение патологии тау в этот 30-дневный момент времени, требуют дальнейшего изучения.
Несмотря на эти данные, демонстрирующие изменения в синаптической физиологии после ЧМТ, многое еще предстоит определить в отношении природы и источника устойчивых физиологических изменений в модели закрытой травмы головы и того, как это связано с дефицитом пластичности и памяти спустя долгое время после начальных компонентов травмы. вылечили. Ca 2+ дисгомеостаз является центральным патогенным элементом и наблюдался в моделях ЧМТ в несколько моментов времени (Deshpande et al., 2008; Сан и др., 2008 г.; Вебер, 2012). Передача сигналов Ca 2+ играет центральную роль в синаптической передаче, пластичности и фосфорилировании тау и, таким образом, может генерировать самоподдерживающийся патогенный цикл, как это наблюдается при хронических нейродегенеративных расстройствах, таких как болезнь Альцгеймера (Stutzmann, 2007; Chakroborty et al., 2009, 2012, 2019). В группе rTBI притупленный ответ RyR-Ca 2+ может отражать снижение запасов ER Ca 2+ в результате негерметичных или сенсибилизированных RyR; эта утечка может также способствовать наблюдаемому повышенному уровню цитозольного Ca 2+ в покое (Lacampagne et al., 2017). Кроме того, повышенное проникновение Ca 2+ через VGCC в сочетании с усилением синаптического возбуждения также может способствовать повышенной нагрузке Ca 2+ . Повышенный уровень клеточного Ca 2+ может впоследствии увеличить активность PKA, которая фосфорилирует и впоследствии сенсибилизирует RyR к утечке избытка ER Ca 2+ в цитозоль (Morimoto et al., 2009; Liu et al., 2012; Lacampagne et al. ., 2017), таким образом снижая запасы ER Ca 2+ и уменьшая вызванные ответы, одновременно способствуя повышению цитозольных уровней покоя, как это наблюдается в этом исследовании.
Несколько клинических исследований продемонстрировали положительное влияние одобренных FDA блокаторов каналов Ca 2+ , таких как нимодипин, на улучшение когнитивных функций у пациентов с ЧМТ (Teasdale et al., 1990, 1992; Bailey et al., 1991), однако последующие анализы не показали влияния на уровень смертности (Langham et al., 2003; Vergouwen et al., 2007). Аналогичным образом, RyR-отрицательный аллостерический модулятор дантролен ослабляет эксайтотоксичность глутамата in vitro (Frandsen and Schousboe, 1991) и in vivo (Niebauer and Gruenthal, 1999) и нормализует внутриклеточный гомеостаз Ca 2+ в моделях БА (Chakroborty) (Chakroborty). др., 2012, 2019), что повышает вероятность того, что измененная передача сигналов Ca 2+ , опосредованная VGCC и RyR, играет центральную роль в долгосрочных последствиях ЧМТ. Учитывая, что VGCC и RyR могут работать с прямой связью для поддержания патофизиологии ЧМТ, комбинированное воздействие на эти каналы может служить эффективной терапевтической стратегией при лечении ЧМТ.
Хотя наше исследование продолжается 30 дней после ЧМТ, необходимы дальнейшие исследования, чтобы определить, сохраняются ли эти эффекты в течение более длительных периодов времени, особенно потому, что гиперактивность гиппокампа может предшествовать синаптической депрессии и нейродегенерации (Huijbers et al., 2015). Кроме того, исследования на людях указывают на когнитивные эффекты, продолжающиеся от нескольких месяцев до нескольких лет (McInnes et al., 2019; Shen et al., 2020), что указывает на необходимость исследований на животных аналогичной продолжительности.
Выводы
Здесь мы продемонстрировали, что как однократная, так и повторная легкая ЧМТ приводит к одинаково стойкому усилению синаптической передачи в гиппокампе, эффекты, сопровождающиеся измененным гомеостазом Ca 2+ и экспрессией фосфорилированного тау. Эти данные подтверждают идею о том, что одно сотрясение мозга может привести к стойким патофизиологическим эффектам головного мозга, которые по величине аналогичны тем, которые наблюдаются после повторных сотрясений, с будущими последствиями для поведения, обучения и памяти.
Заявление о доступности данных
Необработанные данные, подтверждающие выводы этой статьи, будут предоставлены авторами без неоправданных оговорок.
Заявление об этике
Исследование на животных было рассмотрено и одобрено Университетом Розалинды Франклин IACUC Университета ДеПола IACUC.
Вклад авторов
JM, CB и NB провели эксперименты и проанализировали данные, а также внесли свой вклад в написание рукописи. DP, DK и GS участвовали в планировании эксперимента, надзоре, проведении исследования и написании рукописи.Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Финансирование было получено от пилотных проектов DePaul-RFUMS для DP, DK и GS.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Сару Мустали и Шона Шранка за помощь в редактировании.
Каталожные номера
Ахмед, С.М., Вебер, Дж.Т., Лян, С., Уиллоуби, К.А., Ситтердинг, Х.А., Ригалински, Б.А., и соавт. (2002). Активация рецептора NMDA частично способствует снижению потенциала митохондриальной мембраны и повышению уровня внутриклеточного свободного кальция в поврежденных напряжением нейронах. Дж. Нейротравма 19, 1619–1629. дои: 10.1089/089771502762300274
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Альбайрам, О., Герберт, М.К., Кондо А., Цай С.Ю., Бэксли С., Лиан Х. и соавт. (2016). Функция и регуляция конформаций тау при развитии и лечении черепно-мозговой травмы и нейродегенерации. Cell Biosci. 6:59. doi: 10.1186/s13578-016-0124-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Almeida-Suhett, C.P., Prager, E.M., Pidoplichko, V., Figueiredo, T.H., Marini, A.M., Li, Z., et al. (2015). ГАМКергическая межнейрональная потеря и снижение тормозной синаптической передачи в области CA1 гиппокампа после легкой черепно-мозговой травмы. Экспл. Нейрол. 273, 11–23. doi: 10.1016/j.expneurol.2015.07.028
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аннегерс, Дж. Ф., Хаузер, В. А., Коан, С. П., и Рокка, В. А. (1998). Популяционное исследование судорог после черепно-мозговых травм. Н. англ. Дж. Мед. 338, 20–24. дои: 10.1056/NEJM199801013380104
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Араи М., Имамура О., Кондох Н., Датеки М.и Такишима, К. (2019). Нейрональный Ca2+-зависимый белок-активатор 1 (NCDAP1) индуцирует гибель нейронов путем активации пути p53 после черепно-мозговой травмы. Дж. Нейрохим. 151, 795–809. doi: 10.1111/jnc.14803
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аунгст С.Л., Кабади С.В., Томпсон С.М., Стойка Б.А. и Фаден А.И. (2014). Повторное легкое черепно-мозговое повреждение вызывает хроническое нейровоспаление, изменения в синаптической пластичности гиппокампа и связанные с этим когнитивные нарушения. Дж. Цереб. Кровоток Метаб. 34, 1223–1232. doi: 10.1038/jcbfm.2014.75
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Авила Дж., Перес М., Лим Ф., Гомес-Рамос А., Эрнандес Ф. и Лукас Дж. Дж. (2004). Тау при нейродегенеративных заболеваниях: фосфорилирование и сборка тау. Нейротокс Рез. 6, 477–482. дои: 10.1007/BF03033284
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бейли И., Белл А., Грей Дж., Gullan, R., Heiskanan, O., Marks, P.V., et al. (1991). Исследование влияния нимодипина на исход после черепно-мозговой травмы. Акта Нейрохир. 110, 97–105. дои: 10.1007/BF01400674
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бимер М., Туммала С. Р., Гуллотти Д., Копил С., Горка С., Тед А. и другие. (2016). Первичное взрывное повреждение вызывает когнитивные нарушения и изменения в цепи гиппокампа. Экспл. Нейрол. 283, 16–28. doi: 10.1016/j.экспнейрол.2016.05.025
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бигон А., Фрай П.А., Паден С.М., Александрович А., Центер Дж. и Шохами Э. (2004). Динамические изменения рецепторов N-метил-D-аспартата после закрытой черепно-мозговой травмы у мышей: последствия для лечения неврологических и когнитивных нарушений. Проц. Натл. акад. науч. США 101, 5117–5122. doi: 10.1073/pnas.0305741101
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Борде, М., Бонанско, К., Фернандес де Севилья, Д., Ле Рэй, Д., и Буньо, В. (2000). Фиксирующий потенциал анализ потенцирования медленного Ca2+-активируемого K+ тока в пирамидных нейронах гиппокампа. Гиппокамп 10, 198–206. doi: 10.1002/(SICI)1098-1063(2000)10:2<198::AID-HIPO9>3.0.CO;2-F
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бриггс, К.А., Чакроборти, С., и Штутцманн, Г.Е. (2017). Новые пути, приводящие к ранней синаптической патологии при болезни Альцгеймера. Биохим. Биофиз. Рез. коммун. 483, 988–997. doi: 10.1016/j.bbrc.2016.09.088
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бруно, А. М., Хуанг, Дж. Ю., Беннетт, Д. А., Марр, Р. А., Гастингс, М. Л., и Штутцманн, Г. Е. (2012). Изменение экспрессии рианодиновых рецепторов при легких когнитивных нарушениях и болезни Альцгеймера. Нейробиол. Старение 33, 1001.e1–1001.e6. doi: 10.1016/j.neurobiolaging.2011.03.011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чакроборти, С., Briggs, C., Miller, M.B., Goussakov, I., Schneider, C., Kim, J., et al. (2012). Стабилизация функции Са2+-канала ER как стратегия ранней профилактики болезни Альцгеймера. PLoS One 7:e52056. doi: 10.1371/journal.pone.0052056
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чакроборти С., Гусаков И., Миллер М.Б. и Штуцманн Г.Е. (2009). Высвобождение кальция, опосредованное девиантным рианодиновым рецептором, восстанавливает синаптический гомеостаз у мышей с предсимптомной 3xTg-AD. J. Neurosci. 29, 9458–9470. doi: 10.1523/JNEUROSCI.2047-09.2009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чакроборти, С., Хилл, Э.С., Кристиан, Д.Т., Хелфрич, Р., Райли, С., Шнайдер, К., и соавт. (2019). Уменьшенный пресинаптический везикул хранит опосредованные дефекты клеточной и сетевой пластичности на ранней стадии мышиной модели болезни Альцгеймера. Мол. Нейродегенер. 14:7. doi: 10.1186/s13024-019-0307-7
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чакроборти, С., Kim, J., Schneider, C., West, A.R., and Stutzmann, G.E. (2015). Передача сигналов оксида азота задействована в качестве компенсаторного механизма для поддержания синаптической пластичности у мышей с болезнью Альцгеймера. J. Neurosci. 35, 6893–6902. doi: 10.1523/JNEUROSCI.4002-14.2015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чавис, П., Фагни, Л., Лансман, Дж. Б., и Бокерт, Дж. (1996). Функциональная связь между рианодиновыми рецепторами и кальциевыми каналами L-типа в нейронах. Природа 382, 719–722. дои: 10.1038/382719a0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Д’Амброзио Р., Марис Д. О., Грейди М. С., Винн Х. Р. и Джанигро Д. (1998). Избирательная потеря долговременной потенциации гиппокампа, но не депрессия, после перкуссионного повреждения жидкостью. Мозг Res. 786, 64–79. doi: 10.1016/s0006-8993(97)01412-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дэш, П.К., Джонсон, Д., Clark, J., Orsi, S.A., Zhang, M., Zhao, J., et al. (2011). Участие сигнального пути киназы-3 гликогенсинтазы в патологии ЧМТ и нейрокогнитивных исходах. PLoS One 6:e24648. doi: 10.1371/journal.pone.0024648
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дэвис, С. Н., и Коллингридж, Г. Л. (1989). Роль рецепторов возбуждающих аминокислот в синаптической передаче в области СА1 гиппокампа крысы. Проц. Р. Соц. Лонд. Б биол. науч. 236, 373–384. doi: 10.1098/rspb.1989.0028
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дэвис С.Н., Лестер Р.А., Рейманн К.Г. и Коллингридж Г.Л. (1989). Временно различные пре- и постсинаптические механизмы поддерживают долговременную потенциацию. Природа 338, 500–503. дои: 10.1038/338500a0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дешпанде Л.С., Сун Д.А., Сомбати С., Баранова А., Уилсон М.С., Атткиссон Э. и соавт. (2008). Изменения уровня кальция в нейронах связаны с когнитивным дефицитом после черепно-мозговой травмы. Неврологи. лат. 441, 115–119. doi: 10.1016/j.neulet.2008.05.113
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
ДеВос, С.Л., Гончарофф, Д.К., Чен, Г., Кебодо, К.С., Ямада, К., Стюарт, Ф.Р., и соавт. (2013). Антисмысловое снижение уровня тау у взрослых мышей защищает от судорог. J. Neurosci. 33, 12887–12897.doi: 10.1523/JNEUROSCI.2107-13.2013
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эдвардс, Г. III., Чжао, Дж., Даш, П.К., Сото, К., и Морено-Гонсалес, И. (2020). Черепно-мозговая травма вызывает агрегацию и распространение тау. Дж. Нейротравма 37, 80–92. doi: 10.1089/neu.2018.6348
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эллемберг Д., Генри Л. К., Маччиокки С. Н., Гускевич К. М. и Броглио С. П.(2009). Достижения в оценке сотрясения мозга при занятиях спортом: от поведенческих до методов визуализации мозга. Дж. Нейротравма 26, 2365–2382. doi: 10.1089/neu.2009.0906
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Emptage, Н., Блисс, Т.В., и Файн, А. (1999). Одиночные синаптические события вызывают опосредованное рецептором NMDA высвобождение кальция из внутренних запасов в дендритных шипиках гиппокампа. Нейрон 22, 115–124. doi: 10.1016/s0896-6273(00)80683-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фахран, С., Ягер, К., и Алхилали, Л. (2013). Симптоматические изменения белого вещества головного мозга при легкой черепно-мозговой травме напоминают патологические признаки ранней деменции Альцгеймера. Радиология 269, 249–257. doi: 10.1148/radiol.13122343
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Флемингер С., Оливер Д. Л., Лавстон С., Рабе-Хескет С. и Гиора А. (2003). Черепно-мозговая травма как фактор риска болезни Альцгеймера: данные 10 лет спустя; частичная репликация. Дж.Нейрол. Нейрохирург. Психиатрия 74, 857–862. doi: 10.1136/jnnp.74.7.857
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Франдсен, А., и Шоусбо, А. (1991). Дантролен предотвращает цитотоксичность глутамата и высвобождение Ca2+ из внутриклеточных запасов в культивируемых нейронах коры головного мозга. Дж. Нейрохим. 56, 1075–1078. doi: 10.1111/j.1471-4159.1991.tb02031.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гаращук О., Яари, Ю., и Коннерт, А. (1997). Высвобождение и секвестрация кальция рианодин-чувствительными хранилищами в нейронах гиппокампа крысы. J. Physiol. 502, 13–30. doi: 10.1111/j.1469-7793.1997.013bl.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Геддес, Д. М., Лаплака, М. С., и Каргилл, Р. С. II. (2003). Восприимчивость нейронов гиппокампа к механическим повреждениям. Экспл. Нейрол. 184, 420–427. doi: 10.1016/s0014-4886(03)00254-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Геддес, Дж.Ф., Воулз, Г.Х., Николл, Дж.А., и Ревес, Т. (1999). Нейрональные цитоскелетные изменения являются ранним следствием повторяющихся травм головы. Акта Нейропатол. 98, 171–178. дои: 10.1007/s004010051066
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Голарай, Г., Гринвуд, А.С., Фини, Д.М., и Коннор, Дж.А. (2001). Физиологические и структурные доказательства участия гиппокампа в стойкой предрасположенности к судорогам после черепно-мозговой травмы. J. Neurosci. 21, 8523–8537. doi: 10.1523/JNEUROSCI.21-21-08523.2001
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гомес-Рамос, А., Диас-Эрнандес, М., Куадрос, Р., Эрнандес, Ф., и Авила, Дж. (2006). Внеклеточный тау токсичен для нейронов. ФЭБС Письмо. 580, 4842–4850. doi: 10.1016/j.febslet.2006.07.078
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гуркофф, Г. Г., Фэн, Дж.-Ф., Ван, К. С., Изади, А., Ghiasvand, R., Shahlaie, K., et al. (2013). Ингибитор пептидазы NAAG улучшает двигательную функцию и уменьшает когнитивную дисфункцию в модели ЧМТ с вторичной гипоксией. Мозг Res. 1515, 98–107. doi: 10.1016/j.brainres.2013.03.043
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
А. Хазра, Ф. Гу, Аулах А., Берридж К., Эриксен Дж. Л. и Зибуркус Дж. (2013). Дисфункция тормозных нейронов и цепи гиппокампа в модели болезни Альцгеймера у старых мышей. PLoS One 8:e64318. doi: 10.1371/journal.pone.0064318
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эрнандес, А., Тан, К., Платтнер, Ф., Логсдон, А.Ф., Позо, К., Юсуф, М.А., и соавт. (2018). Воздействие слабой взрывной волны вызывает невропатологические эффекты, нейрофизиологический дефицит и биохимические изменения. Мол. Мозг 11:64. doi: 10.1186/s13041-018-0408-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хинцман, Дж.M., Thomas, T.C., Burmeister, J.J., Quintero, J.E., Huettl, P., Pomerleau, F., et al. (2010). Диффузное повреждение головного мозга повышает тонические уровни глутамата и вызванное калием высвобождение глутамата в отдельных областях мозга через два дня после травмы: исследование массива микроэлектродов на основе ферментов. Дж. Нейротравма 27, 889–899. doi: 10.1089/neu.2009.1238
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холбро Н., Грундиц А. и Ортнер Т. Г. (2009). Дифференциальное распределение эндоплазматического ретикулума контролирует метаботропную передачу сигналов и пластичность в синапсах гиппокампа. Проц. Натл. акад. науч. США 106, 15055–15060. doi: 10.1073/pnas.00106
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Huang, Y.Y., Colino, A., Selig, D.K., и Malenka, R.C. (1992). Влияние предшествующей синаптической активности на индукцию долговременной потенциации. Наука 255, 730–733. doi: 10.1126/science.1346729
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хуанг Ю. Ю. и Маленко Р.С. (1993). Исследование индуцированного ТЭА синаптического усиления в области СА1 гиппокампа: роль потенциалзависимых каналов Са2+ в индукции ДП. J. Neurosci. 13, 568–576. doi: 10.1523/JNEUROSCI.13-02-00568.1993
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хубер, К.М., Маук, М.Д., и Келли, П.Т. (1995). LTP, индуцированный активацией потенциал-зависимых каналов Ca2+, требует активности протеинкиназы. Нейроотчет 6, 1281–1284.дои: 10.1097/00001756-199506090-00013
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Huijbers, W., Mormino, E.C., Schultz, A.P., Wigman, S., Ward, A.M., Larvie, M., et al. (2015). Отложение β-амилоида при легких когнитивных нарушениях связано с повышенной активностью гиппокампа, атрофией и клиническим прогрессированием. Мозг 138, 1023–1035. doi: 10.1093/мозг/awv007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хейберс, В., Schultz, A.P., Papp, K.V., LaPoint, M.R., Hanseeuw, B., Chhatwal, J.P., et al. (2019). Накопление тау у клинически здоровых пожилых людей связано с гиперактивностью гиппокампа. J. Neurosci. 39, 548–556. doi: 10.1523/JNEUROSCI.1397-18.2018
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ял, К., Алонсо Адель, К., Чен, С., Чохан, М. О., Эль-Аккад, Э., Гонг, С. X., и соавт. (2005). Тау-патология при болезни Альцгеймера и других таупатиях. Биохим.Биофиз. Acta 1739, 198–210. doi: 10.1016/j.bbadis.2004.09.008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джаффе, Д.Б., Фишер, С.А., и Браун, Т.Х. (1994). Конфокальная лазерная сканирующая микроскопия выявляет потенциалзависимые кальциевые сигналы в дендритных шипиках гиппокампа. Дж. Нейробиол. 25, 220–233. doi: 10.1002/neu.480250303
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ямния, Н., Урбан, Дж. Х., Штутцманн, Г.E., Chiren, S.G., Reisenbigler, E., Marr, R., et al. (2017). Клинически значимая закрытая модель однократного и повторного сотрясения мозга у взрослой крысы с использованием устройства для контролируемого воздействия на кору головного мозга. Дж. Нейротравма 34, 1351–1363. doi: 10.1089/neu.2016.4517
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Koenig, J.B., Cantu, D., Low, C., Sommer, M., Noubary, F., Croker, D., et al. (2019). Гликолитический ингибитор 2-дезоксиглюкоза предотвращает гипервозбудимость коры после черепно-мозговой травмы. JCI Insight 5:e126506. doi: 10.1172/jci.insight.126506
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Lacampagne, A., Liu, X., Reiken, S., Bussiere, R., Meli, A.C., Lauritzen, I., et al. (2017). Посттрансляционное ремоделирование рианодинового рецептора вызывает утечку кальция, что приводит к патологиям, подобным болезни Альцгеймера, и когнитивным нарушениям. Акта Нейропатол. 134, 749–767. doi: 10.1007/s00401-017-1733-7
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лэнгэм, Дж., Голдфрад, К., Тисдейл, Г., Шоу, Д., и Роуэн, К. (2003). Блокаторы кальциевых каналов при острой черепно-мозговой травме. Кокрановская система базы данных. Ред. 4:CD000565. дои: 10.1002/14651858.CD000565
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Li, Y., Li, Y., Li, X., Zhang, S., Zhao, J., Zhu, X., et al. (2017). Травма головы как фактор риска деменции и болезни Альцгеймера: систематический обзор и метаанализ 32 обсервационных исследований. PLoS One 12:e0169650.doi: 10.1371/journal.pone.0169650
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лю, Х., Бетценхаузер, М.Дж., Райкен, С., Мели, А.С., Се, В., Чен, Б.Х., и соавт. (2012). Роль негерметичных нейрональных рианодиновых рецепторов в вызванной стрессом когнитивной дисфункции. Сотовый 150, 1055–1067. doi: 10.1016/j.cell.2012.06.052
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лог, О. К., Крамер, Н. П., Сюй, X., Перл, Д. П., и Галдзицки, З.(2016). Изменения функциональных свойств сетей гиппокампа после повторной закрытой черепно-мозговой травмы. Экспл. Нейрол. 277, 227–243. doi: 10.1016/j.expneurol.2015.12.019
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Луо П., Фей Ф., Чжан Л., Цюй Ю. и Фей З. (2011). Роль глутаматных рецепторов при черепно-мозговой травме: значение постсинаптической плотности в патофизиологии. Мозг Res. Бык. 85, 313–320. doi: 10.1016/j.мозгресбулл.2011.05.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Макиннес, К., Фризен, К.Л., Маккензи, Д.Э., Вествуд, Д.А., и Бо, С.Г. (2019). Легкая черепно-мозговая травма (mTBI) и хронические когнитивные нарушения: предварительный обзор. PLoS One 14:e0218423. doi: 10.1371/journal.pone.0218423
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Макки, А. К., Канту, Р. К., Новински, С. Дж., Хедли-Уайт, Э. Т., Gavett, B.E., Budson, A.E., et al. (2009). Хроническая травматическая энцефалопатия у спортсменов: прогрессирующая таупатия после повторной травмы головы. Дж. Невропатол. Эксп. Нейрол. 68, 709–735. дои: 10.1097/NEN.0b013e3181a9d503
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
McKee, A.C., Stern, R.A., Nowinski, C.J., Stein, T.D., Alvarez, V.E., Daneshvar, D.H., et al. (2013). Спектр заболеваний при хронической травматической энцефалопатии. Мозг 136, 43–64.дои: 10.1093/мозг/aws307
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Михан, В. П. III., Чжан, Дж., Мэнникс, Р., и Уэлен, М. Дж. (2012). Увеличение времени восстановления между травмами улучшает когнитивный результат после повторяющихся легких сотрясений головного мозга у мышей. Нейрохирургия 71, 885–891. doi: 10.1227/NEU.0b013e318265a439
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Миядзаки С., Катаяма Ю., Лиет Б.Г., Дженкинс, Л.В., ДеВитт, Д.С., Голдберг, С.Дж., и соавт. (1992). Устойчивое подавление долговременной потенциации гиппокампа после черепно-мозговой травмы у крыс. Мозг Res. 585, 335–339. дои: 10.1016/0006-8993(92)-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Моримото С., Джин О. У., Каваи М., Хосина Т., Кусакари Ю., Комукай К. и др. (2009). Зависимое от протеинкиназы А фосфорилирование рианодиновых рецепторов увеличивает утечку Са2+ в сердце мыши. Биохим. Биофиз. Рез. коммун. 390, 87–92. doi: 10.1016/j.bbrc.2009.09.071
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нибауэр, М., и Грюнталь, М. (1999). Нейропротекторные эффекты раннего и позднего введения дантролена при экспериментальном эпилептическом статусе. Нейрофармакология 38, 1343–1348. doi: 10.1016/s0028-3908(99)00059-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Улес, Б., Дель Прете, Д., Greco, B., Zhang, X., Lauritzen, I., Sevalle, J., et al. (2012). Блокада рианодиновых рецепторов уменьшает нагрузку β-амилоида и нарушения памяти у мышей Tg2576, моделирующих болезнь Альцгеймера. J. Neurosci. 32, 11820–11834. doi: 10.1523/JNEUROSCI.0875-12.2012
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Петралья, А.Л., Дашнав, М.Л., Тернер, Р.К., и Бейлз, Дж.Э. (2014). Модели легкой черепно-мозговой травмы: перевод физиологических и анатомических повреждений. Нейрохирургия 75, S34–S49. doi: 10.1227/NEU.0000000000000472
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ривз, Т.М., Лайет, Б.Г., Филлипс, Л.Л., Хамм, Р.Дж., и Повлишок, Дж.Т. (1997). Влияние черепно-мозговой травмы на торможение в гиппокампе и зубчатой извилине. Мозг Res. 757, 119–132. doi: 10.1016/s0006-8993(97)00170-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сандлер, В.М. и Барбара, Дж. Г. (1999). Высвобождение кальция, индуцированное кальцием, способствует переходным процессам кальция, вызванным потенциалом действия, в пирамидных нейронах CA1 гиппокампа. J. Neurosci. 19, 4325–4336. doi: 10.1523/JNEUROSCI.19-11-04325.1999
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шранк С., Баррингтон Н. и Штутцманн Г. Э. (2020). Дефекты обращения с кальцием и нейродегенеративные заболевания. Гавань Колд Спринг. Перспектива. биол. 12:a035212. дои: 10.1101/cshperspect.a035212
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шварцбах Э., Бониславски Д. П., Сюн Г. и Коэн А. С. (2006). Механизмы, лежащие в основе неспособности индуцировать LTP области CA1 у мышей после черепно-мозговой травмы. Гиппокамп 16, 541–550. doi: 10.1002/hipo.20183
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шэнь, И. Х., Линь, Ю.-Дж., Чен, К.-Л., и Ляо, К.-К. (2020). Нейронные корреляты торможения реакции и обработки ошибок у людей с легкой черепно-мозговой травмой: потенциальное исследование, связанное с событием. Дж. Нейротравма 37, 115–124. doi: 10.1089/neu.2018.6122
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шим С.С. и Штутцманн Г.Е. (2016). Ингибирование киназы-3 гликогенсинтазы: новая цель в лечении черепно-мозговой травмы. Дж. Нейротравма 33, 2065–2076. doi: 10.1089/neu.2015.4177
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Слеммер, Дж. Э., Мацер, Э. Дж., Де Зеув, К. И.и Вебер, Дж. Т. (2002). Повторное легкое повреждение вызывает кумулятивное повреждение клеток гиппокампа. Мозг 125, 2699–2709. doi: 10.1093/мозг/awf271
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Штуцманн, Г. Э., Каккамо, А., ЛаФерла, Ф. М., и Паркер, И. (2004). Нарушение регуляции передачи сигналов IP3 в кортикальных нейронах нокаутированных мышей, экспрессирующих связанную с болезнью Альцгеймера мутацию пресенилина 1, приводит к преувеличенным сигналам Ca2+ и изменению возбудимости мембран. J. Neurosci. 24, 508–513. doi: 10.1523/JNEUROSCI.4386-03.2004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Штуцманн, Г. Э., ЛаФерла, Ф. М., и Паркер, И. (2003). Передача сигналов Ca2+ в нейронах коры головного мозга мышей изучалась с помощью двухфотонной визуализации и фотовысвобождения инозитолтрифосфата. J. Neurosci. 23, 758–765. doi: 10.1523/JNEUROSCI.23-03-00758.2003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сан, Д.А., Дешпанде Л.С., Сомбати С., Баранова А., Уилсон М.С., Хамм Р.Дж. и соавт. (2008). Черепно-мозговая травма вызывает длительное кальциевое (Ca2+)-плато повышенных внутриклеточных уровней Ca и изменение гомеостатических механизмов Ca2+ в нейронах гиппокампа, выживших после травмы головного мозга. евро. Дж. Нейроски. 27, 1659–1672 гг. doi: 10.1111/j.1460-9568.2008.06156.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тейлор, К.А., Белл, Дж.М., Брейдинг, М.Дж., и Сюй, Л.(2017). Посещения отделений неотложной помощи в связи с черепно-мозговой травмой, госпитализации и смерти — США, 2007 и 2013 гг. MMWR Surveill. Сумма. 66, 1–16. doi: 10.15585/mmwr.ss6609a1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тисдейл Г., Бейли И., Белл А., Грей Дж., Гуллан Р., Хейсканан О. и др. (1992). Рандомизированное исследование нимодипина при тяжелой травме головы: HIT I. Британско-финская совместная исследовательская группа по травмам головы. J. Нейротравма 9, S545–S550.
Реферат PubMed | Академия Google
Тисдейл Г., Бейли И., Белл А., Грей Дж., Гуллан Р., Хейсканан У. и др. (1990). Влияние нимодипина на исход после черепно-мозговой травмы: проспективное рандомизированное контролируемое исследование. Британско-финская совместная судебная группа по травмам головы. Акта Нейрохир. Доп. 51, 315–316. дои: 10.1007/978-3-7091-9115-6_106
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Теадом, А., Пармар, П., Джонс, К., Barker-Collo, S., Starkey, N.J., McPherson, K.M., et al. (2015). Частота и влияние рецидивирующей черепно-мозговой травмы в популяционной выборке. Дж. Нейротравма 32, 674–681. doi: 10.1089/neu.2014.3579
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тибо, О., Гант, Дж. К., и Лэндфилд, П. В. (2007). Расширение кальциевой гипотезы старения мозга и болезни Альцгеймера: присматривать за магазином. Ячейка старения 6, 307–317. дои: 10.1111/j.1474-9726.2007.00295.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Титус, Д. Дж., Джонстон, Т., Джонсон, Н. Х., Лондон, С. Х., Чапаламадугу, М., Хогенкамп, Д., и др. (2019). Положительная аллостерическая модуляция никотинового ацетилхолинового рецептора α7 как лечение когнитивного дефицита после черепно-мозговой травмы. PLoS One 14:e0223180. doi: 10.1371/journal.pone.0223180
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тран, Х.Т., Санчес Л. и Броуди Д.Л. (2012). Ингибирование JNK пептидным ингибитором уменьшает таупатию, вызванную черепно-мозговой травмой, у трансгенных мышей. Дж. Невропатол. Эксп. Нейрол. 71, 116–129. doi: 10.1097/NEN.0b013e3182456aed
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Циен, Дж. З., Уэрта, П. Т., и Тонегава, С. (1996). Существенная роль синаптической пластичности, зависящей от рецептора CA1 NMDA гиппокампа, в пространственной памяти. Сотовый 87, 13:27–13:38.doi: 10.1016/s0092-8674(00)81827-9
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Vergouwen, M.D., Vermeulen, M., de Haan, R.J., Levi, M., and Roos, Y.B. (2007). Дигидропиридиновые антагонисты кальция повышают фибринолитическую активность: систематический обзор. Дж. Цереб. Кровоток Метаб. 27, 1293–1308. doi: 10.1038/sj.jcbfm.9600431
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вебер, Дж. Т., Ригалински, Б. А., и Эллис, Э.Ф. (2002). Реакции кальция на кофеин и агонисты мускариновых рецепторов изменяются в травматически поврежденных нейронах. Дж. Нейротравма 19, 1433–1443. дои: 10.1089/089771502320
0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вейл З.М., Гайер К.Р. и Карелина К. (2014). Время травмы изменяет метаболические, воспалительные и функциональные исходы после повторной легкой черепно-мозговой травмы. Нейробиол. Дис. 70, 108–116. дои: 10.1016/j.nbd.2014.06.016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уайт, Э. Р., Пинар, К., Бостром, К. А., Мекони, А., и Кристи, Б. Р. (2017). Легкая черепно-мозговая травма вызывает длительный дефицит синаптической пластичности в ювенильном гиппокампе самок. Дж. Нейротравма 34, 1111–1123. doi: 10.1089/neu.2016.4638
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уилсон С. М., Ки Ён С., Ян X.-Ф., Пак К.Д. и Ханна Р. (2014). Дифференциальная регуляция фосфорилирования белка-медиатора коллапсинового ответа 2 (CRMP2) с помощью GSK3ss и CDK5 после черепно-мозговой травмы. Фронт. Клетка. Неврологи. 8:135. doi: 10.3389/fncel.2014.00135
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вольф, Дж. А., Стис, П. К., Лусарди, Т., Мини, Д., и Смит, Д. Х. (2001). Травматическое повреждение аксонов вызывает приток кальция, модулируемый чувствительными к тетродотоксину натриевыми каналами. J. Neurosci. 21, 1923–1930. doi: 10.1523/JNEUROSCI.21-06-01923.2001
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Yamada, K., Patel, T.K., Hochgräfe, K., Mahan, T.E., Jiang, H., Stewart, F.R., et al. (2015). Анализ оборота тау-белка in vivo в мышиной модели таупатии. Мол. Нейродегенер. 10:55. doi: 10.1186/s13024-015-0052-5
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжан, Б., Chen, X., Lin, Y., Tan, T., Yang, Z., Dayao, C., et al. (2011). Нарушение синаптической пластичности в гиппокампе усугубляется метилпреднизолоном в крысиной модели черепно-мозговой травмы. Мозг Res. 1382, 165–172. doi: 10.1016/j.brainres.2011.01.065
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Zhang, B.-L., Chen, X., Tan, T., Yang, Z., Carlos, D., Jiang, R.-C., et al. (2011). Черепно-мозговая травма нарушает синаптическую пластичность гиппокампа крыс. Подбородок. Мед. J. 124, 740–745.
Реферат PubMed | Академия Google
Чжан Х., Лю Дж., Сун С., Пчицкая Э., Попугаева Э. и Безпрозванный И. (2015). Дефекты передачи сигналов кальция, возбудимости и синаптической пластичности в мышиной модели болезни Альцгеймера. J. Alzheimers Dis. 45, 561–580. дои: 10.3233/JAD-142427
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжао, С., Фу, Дж., Лю, X., Ван, Т., Чжан, Дж.и Чжао Ю. (2012). Активация сигнального пути Akt/GSK-3β/β-катенин участвует в выживании нейронов после черепно-мозговой травмы у крыс. Нейрол. Рез. 34, 400–407. дои: 10.1179/1743132812Y.0000000025
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Как COVID-19 изменит наши школы в долгосрочной перспективе?
В разгар беспрецедентного кризиса трудно заглянуть в будущее дальше, чем на несколько дней. Мы словно блуждаем в густом (и смертельном) тумане.
Некоторые комментаторы предсказывают, что это изменит наш образ жизни; один даже предсказывает, что это «изменит нас как биологический вид». Возможно, но каким образом? Мы обязательно запомним это время на всю оставшуюся жизнь. По крайней мере, ненадолго, мы будем ценить маленькие вещи в жизни немного больше. Но изменит ли это что-то принципиально в долгосрочной перспективе? Если да, то как?
Люди задавались тем же вопросом после урагана Катрина в Новом Орлеане. Жизнь здесь определяется как «до Катрины» и «после Катрины».«Никто в Новом Орлеане не стал бы утверждать, что город не изменился. Теперь город лучше защищен от улучшенных дамб и других средств управления водными ресурсами. Население меньше (и белее).
Но когда люди говорят, что «все никогда не будет прежним», они говорят о чем-то более глубоком, о том, как мы живем — о наших привычках, нормах и образе жизни. Для родителей, учителей и учащихся некоторые аспекты школьного обучения могут не вернуться к тому, что было раньше.
Возможные изменения в школах
Я думал о том, чему мы научились у Катрины, и о том, как изменится жизнь, особенно как изменятся школы.После Катрины Новый Орлеан превратился в то, что в конечном итоге стало первым в стране городом с чартерными школами. Нет, я не предсказываю, что мы будем делать что-то подобное из-за коронавируса. Но, как я объясняю в своей готовящейся к выходу книге «Чартерный школьный город», в Новом Орлеане было много могущественных сил, которые сделали школьные реформы реальностью. Некоторые из этих сил сильны и сейчас.
С COVID-19 школы быстро меняют основные методы работы. Некоторые стали старомодными заочными школами, где подавляющее большинство взаимодействий происходит по письменной почте.Другие пытались воссоздать школьную обстановку онлайн с помощью цифровых инструментов, таких как Zoom. Другие находятся между ними, направляя студентов на онлайн-репетиторство и программы практики, а также публикуя видео. Большинство людей думают, что они просто хотят вернуться к нормальной жизни. В этом есть смысл. В конце концов, школы не сделали ничего, чтобы вызвать кризис. Так зачем их менять?
Так многие думали после Катрины. Школы не вызвали опустошения, так зачем их менять? Тем не менее, они изменились. Государство взяло на себя почти все государственные школы города, в конечном итоге передав их деятельность некоммерческим чартерным организациям.Срок полномочий учителя и профсоюзный договор закончились. Зоны посещаемости, которые распределяют учащихся по школам, которые они посещают, в зависимости от того, где они живут, были упразднены, чтобы у семей была возможность выбрать любую школу по своему желанию. Лишь горстка городов когда-либо делала что-то из этого раньше — Новый Орлеан делал все сразу.
Здесь я сосредоточусь на части новоорлеанской реформы «выбор школы», потому что она особенно показательна. После «Катрины» городу, по сути, пришлось ликвидировать зоны посещаемости и переключиться на выбор школы, потому что вернувшееся население было слишком рассредоточено, чтобы в любой «зоне» могло быть достаточно учеников.Кроме того, семьи переезжали из одного временного дома в другой; зоны потребовали бы, чтобы ученики постоянно меняли школу. Таким образом, выбор школы был отчасти практическим — чтобы справиться с кризисом. Выбор школы вдруг стал необходим.
И угадайте, что случилось? Семьи привыкли к выбору. В то время как растет озабоченность по поводу расстояния, которое дети должны преодолевать, чтобы ходить в школу, и округ начинает разрешать району быть одним из факторов при распределении учеников, о возвращении к традиционным зонам посещаемости почти не было разговоров, хотя шторм и большинство его последствий давно прошли.
Переход к выбору привел и к другим изменениям. Например, окончание зон посещаемости означает, что большинство учащихся больше не ходят в те же школы, что и те, кто живет в соседнем квартале. Кроме того, это означает, что ни дети, ни родители, проживающие в одном и том же районе, не развивают такие же общественные связи, как раньше. Кроме того, как утверждают сторонники выбора, выбор позволяет учащимся сопоставлять свои потребности со школьными предложениями.
Какие уроки мы можем извлечь из этого?
Чему учит нас дело Нового Орлеана/Катрины? Во-первых, кризисы заставляют нас адаптировать . Катрина заставила измениться, и это изменение привело к выбору школы. В условиях нынешнего кризиса COVID заставляет родителей быть учителями и вынуждает всех — учащихся, родителей и учителей — адаптироваться к инструментам онлайн-обучения.
Во-вторых, люди привыкают к некоторым из этих приспособлений. Катрина сделала выбор школы в Новом Орлеане удобным. Точно так же, в то время как семьи сейчас испытывают стресс, пытаясь дать образование своим детям, они также испытывают образовательные методы и инструменты, которых они никогда раньше не видели.К ним все больше привыкают.
Это не значит, что все онлайн-инструменты очень хороши. Многие нет. Но рассмотрите следующее: предположим, учитель пробует три онлайн-инструмента во время кризиса. Она любит инструмент А, не любит инструмент Б и безразлична к инструменту С. Это не совсем похоже на рецепт массового преобразования, не так ли? Ну, на самом деле, если бы кризиса не было, учитель никогда бы не узнал ни об одном из этих инструментов и в любом случае не использовал бы B или C.Суть в том, что учителя (и, возможно, ученики и родители) теперь хотят больше пятерки, и это может изменить ситуацию.
В-третьих, наши адаптации имеют косвенные эффекты, которые приводят к другим изменениям. Необходимый переход к выбору школы изменил наши районы неожиданным образом. В условиях нынешнего кризиса переход на онлайн-инструменты также может иметь косвенные последствия.
Потенциальные долгосрочные сдвиги от COVID-19
Ниже я применяю описанную выше логику к нескольким областям политики, которые недавно обсуждались в контексте коронавируса.Первые две темы — онлайн-инструменты и полностью онлайн-обучение — приходят на ум в первую очередь. Остальное — это потенциальные долгосрочные сдвиги, которые могут попасть в категорию «косвенных».
Использование онлайн-инструментов? Из приведенных выше аргументов должно быть ясно, что школы будут намного шире использовать онлайн-инструменты. Большинство студентов в стране скоро будут иметь ноутбуки и какой-либо доступ в Интернет (хотя цифровое неравенство останется серьезной проблемой). Учителям понравятся многие инструменты, и им будет легче их использовать теперь, когда учащиеся имеют некоторый опыт работы с ними.Как недавно отметил Дэйв Деминг, онлайн-инструменты могут быть полезным дополнением к очному обучению, а не его заменой, позволяя учителям больше сосредоточиться на вовлечении учащихся и их наставничестве.
Переход к домашнему обучению и полностью виртуальному обучению? Возможны некоторые сдвиги в этом направлении. Семьи будут больше привыкать к онлайн-обучению. Однако у этого подхода есть существенный недостаток, заключающийся в том, что семьям приходится играть роль смотрителя зала и учителя.Немногие семьи хотят или могут себе это позволить, учитывая их график работы и другие обязанности. Более того, исследования постоянно показывают, что учащиеся меньше учатся в полностью виртуальной среде. Личное обучение под руководством учителя просто имеет слишком много преимуществ.
Переход на чартерные школы? Ключевой вопрос: какие школы лучше справятся с текущим кризисом? Возможно, традиционные государственные школы реагируют лучше, а это означает, что они предоставляют более качественные образовательные услуги детям и их семьям.Это возможно, потому что они предназначены для большей емкости. У них есть ИТ-отделы и директора, которые занимаются специальным обучением, закупками и многим другим. Они извлекают выгоду из экономии за счет масштаба и опыта. С другой стороны, в чартерных школах меньше правил, которым нужно следовать. С меньшим количеством государственных правил и более ограниченными профсоюзными контрактами чартерные школы могут быть более гибкими в реагировании на кризис.
Пока рано говорить, какой сектор победит. В одном недавнем отчете говорится, что организации по управлению чартерами (CMO) перешли на онлайн-обучение более агрессивно, чем традиционные государственные школы, хотя большинство чартерных школ на самом деле не управляются CMO.Кроме того, ОКУ часто получают значительные дополнительные средства от благотворителей, что дает им преимущество.
Но если какой-то сектор отреагирует лучше, это будет значимая победа, которую, несомненно, заметят родители. Школы, которые реагируют лучше всего, могут рассчитывать на то, что их выберет больше родителей, и рассчитывать на большую политическую поддержку.
Переход в частные школы? Все вышесказанное о чартерных школах также относится и к частным школам, за одним исключением: частные школы, скорее всего, пострадают в финансовом отношении.За исключением небольшой части студентов, обучающихся по ваучерам, семьи должны платить за обучение. Хотя семьи среднего класса, посещающие частные школы, не так сильно пострадают от экономического кризиса, вызванного COVID-19, ощутимо пострадают все. В течение следующего года, вероятно, мы увидим большой всплеск закрытия частных школ.
Переход к компетентностному обучению? Некоторые эксперты в области образования утверждают, что это может стать большим плюсом текущего кризиса.Однако я думаю, что это маловероятно. Как и в случае с домашним обучением, подходы, основанные на компетенциях, имеют серьезные ограничения. В то время как они позволяют учащимся учиться в своем собственном темпе, подходы, основанные на компетенциях, дробят обучение и в значительной степени полагаются на стандартизированные тесты. Студенты предоставляют свою компетенцию, и могут перейти к следующей теме, только пройдя тест. Обучение на основе компетенций является «персонализированным» в том смысле, что обучение адаптируется на основе существующих навыков, но, опять же, в рамках тестов. Некоторые из них лучше, чем другие, но я все еще не верю, что подходы, основанные на компетенциях, будут теми, к которым тяготеют учителя и ученики в условиях нынешнего кризиса.Обучение на основе компетенций слишком сильно страдает от тех же проблем, что и тестирование с высокими ставками в целом, которое потеряло популярность.
Да, в краткосрочной перспективе нам потребуется больше подходов, основанных на компетенциях, чтобы определить, какие учащиеся будут переведены в следующий класс, учитывая потерянное время обучения. Но значительный долгосрочный переход к онлайн-обучению кажется менее вероятным.
Смена ролей учеников, родителей и учителей? Переход на некоторые онлайн-инструменты может изменить роль учителей, сделав их больше похожими на коучей и наставников.Они могут указать студентам на очень хорошие онлайн-лекции, а затем быть там, чтобы дать рекомендации и отзывы, а также установить связи между темами. Роли учащихся и родителей также могут измениться. Теперь, когда у них есть больше мест для поиска, они, возможно, с большей вероятностью попытаются решить свои потребности в обучении самостоятельно. Когда роли меняются, все остальное может измениться вместе с ними, хотя и менее предсказуемым образом.
Еще один косвенный эффект: политика образования
Конечно, главное изменение в системе школьного образования, которое произошло после «Катрины», заключалось в том, что несколько лидеров решили, что на местный школьный округ больше не следует возлагать ответственность за образование детей — вот почему моя книга называется «Чартерный школьный город», а не «Традиционный школьный город». Государственная школа города.Но дело было не в силах наверху. Это было о политике.
Я не в плохом смысле. Политика — это то, как мы принимаем коллективные решения. Это включает в себя ценности и борьбу за власть. В следующем посте я рассмотрю, как кризис COVID-19 может (или не может) изменить политику образования. Может ли это привести к большим изменениям в государственной политике, которые еще не очевидны? Можем ли мы увидеть сокращение государственного и федерального регулирования? Больше гибкости для студентов, выбирающих курсы и инструменты?
Форма новоорлеанских реформ оставалась неясной до нескольких месяцев после того, как шторм обрушился на сушу.