
Расшифровка маркировки компактных люминесцентных ламп
Код из трёх цифр на упаковке лампы содержит, как правило, информацию относительно качества света (индекс цветопередачи и цветовой температуры).
Первая цифра — индекс цветопередачи в 1х10 Ra (чем выше индекс, тем достоверней цветопередача; компактные люминесцентные лампы имеют 60‒98 Ra)
Вторая и третья цифры — указывают на цветовую температуру лампы.
Таким образом, маркировка «827» указывает на индекс цветопередачи в 80 Ra, и цветовую температуру в 2700 К (что соответствует цветовой температуре лампы накаливания).
Наиболее распространены компактные люминесцентные лампы с коррелированной цветовой температурой 2700K, 4000K, 4500K, 6500K.
Кроме того, индекс цветопередачи может обозначаться в соответствии с DIN 5035, где диапазон цветопередачи 20‒100 Ra поделён на 6 частей — от 4 до 1А.
Пример маркировки КЛЛ
|
Параметр |
Значение |
|
Потребляемая мощность |
11 Вт |
|
Световой поток |
535 лм |
|
Цветовая температура |
2700 К |
|
Тип цоколя |
Е27 |
|
Напряжение |
220‒240 В |
|
Частота питающей сети |
50/60 Гц |
|
Номинальный срок службы (при работе примерно 2,7 часов в день / время службы) |
8 лет |
Работа 2,7 ч/день или 2,74 ч/день указывается производителями из-за простоты расчётов и сравнения с другими типами ламп, так как при таком графике лампа за один год прогорает примерно 1000 ч. Столь малое время работы в сутки производители объясняют средним временем работы всех ламп в квартире, включая в расчёт и те, которые используются короткое время (например в санузле).
Столь малое время работы в сутки производители объясняют средним временем работы всех ламп в квартире, включая в расчёт и те, которые используются короткое время (например в санузле).
Расшифровка обозначений светильников
Что означают краткие названия светильников?
С целью раскрыть терминологию условных обозначений светильников, мы сделали расшифровку кратких наименований светильников. Надеемся, что представленная информация будет вам полезна.
1. Первая буква указывает на источник света — Лампа:
-
-
Н – лампа накаливания
-
Л – люминесцентная лампа
-
Э – люминесцентная ртутная эритемная лампа (используется чаще всего в медицине)
-
Р – ртутная лампа ДРЛ
-
Г – ртутная лампа типа ДРИ, ДРИШ
-
Ж — натриевая лампа типа ДНаТ
-
-
К – ксеноновая трубчатая лампа
2. Втора буква указывает на способ монтажа светильника:
Втора буква указывает на способ монтажа светильника:
-
С – подвесное крепление
-
П – потолочное крепление
-
В – встраиваемый монтаж
-
П – накладной монтаж
-
Б – настенное крепление
-
Н – настольное размещение
-
Т – напольное размещение
-
К – крепление на консоль
-
Р – ручного применения
-
Г – налобное (головное) применение
3. Третья буква указывает на вид использования светильника:
-
П – светильники для промышленных и производственных зданий
-
О — для применения в общественных помещениях
-
Б – для применения в жилых помещениях
-
У — для уличного (наружного) освещения
-
Р — для применения в шахтах и рудниках
-
Т — для применения в телевизионных студиях
4. Двухзначное число (например, 42) указывают на номер серии светильника
Двухзначное число (например, 42) указывают на номер серии светильника
5. Далее цифры могут указывать количество используемых ламп в светильнике
6. Трехзначная цифра (001 -999), указывает на номер дополнительной серии
7. Если используется буква и цифра, это указывает на климатическое исполнение и тип размещения светильника.
Типы компактных люминесцентных ламп (КЛЛ)
Форма лампы
Типы компактных люминесцентных ламп
Лампа с двумя трубками или U-образная лампа
Компактная люминесцентная лампа с двумя трубками, расположенными параллельно. Обычно предназначена для настольных ламп, рабочего освещения, встраиваемых в стены и потолки светильников. Самый доступный тип КЛЛ.
Лампа с четырьмя трубками
Этот тип компактных люминесцентных ламп с четырьмя трубками в два раза короче, но выдает практически столько же света, сколько и двухтрубочные разновидности такой же мощности. Прекрасно подходит для установки в небольшие светильники.
Лампа с тремя трубками
Выдает больше света, чем четырехтрубочный вид компактных люминесцентных ламп, являясь при этом еще короче. Лампы такой формы очень часто используются в качестве замены лампе накаливания благодаря своим небольшим размерам, достигнутым за счет более эффективного использования объема, а также невысокой цене.
Лампа с трубкой в форме спирали
Всего одна трубка, закрученная в спираль. Самый компактный вид ламп, максимально приближенный по габаритам к лампе накаливания. К тому же выглядят они очень симпатично — не стыдно в люстру вкрутить. Но обычно спиральные лампы немного дороже своих более прямых родственников из-за сложной технологии производства.
Лампа типа F
В этом виде компактных люминесцентных ламп две двойные трубки расположены в одной плоскости. Производители решили, что такая форма напоминает букву F. Эти лампы идеально подходят для освещения больших помещений с помощью модульных светильников, плоских квадратных накладных или встраиваемых (в стены или потолок) светильников, а также светильников, которые направляют свет снизу вверх.
Кольцевая лампа
Немного экзотичный, но тем не менее очень эффективный тип КЛЛ. Свет такой лампы распределяется наилучшим образом именно благодаря ее кольцевой форме.
Конечно, это не все возможные разновидности компактных люминесцентных ламп, а только самые распространенные. КЛЛ могут быть и шеститрубочными, и квадратными, и других причудливых форм.
Тип цоколя
Стандартный винтовой цоколь типа E: E14, E27, E40 (число обозначает диаметр в милиметрах) — в точности такой же, как и у обычных ламп накаливания. Внутри цоколя обычно расположен ЭПРА (электронный пускорегулирующий аппарат). Такие лампы можно использовать в любом светильнике, предназначенном для ламп накаливания. Если они туда поместятся, конечно.
Цоколь типа G — цоколь со штырьками (обычно их 2 или 4), в котором, например, может располагаться либо только встроенный стартер (G23), либо вовсе отсутствовать (G24). Поэтому лампы с такими разъемами подходят только для использования в специальных моделях светильников, оснащенных дросселями или ПРА (пускорегулирующими аппаратами, чаще всего электромагнитными — ЭмПРА).
Ртутные лампы ДРВ прямого включения
Сортировать по:
Сортировать по:
| Производители электрооборудованияНажмите на логотип производителя чтобы посмотреть все его товары в этом разделе. |
 Однако для их использования требуется специальный пускорегулирующий аппарат, который серьезно осложняет конструкцию.
Однако для их использования требуется специальный пускорегулирующий аппарат, который серьезно осложняет конструкцию.
Световой поток светодиодных ламп, накаливания, ДРЛ, ДНАТ
Времена правления ламп накаливания в наших домах уже подошли к концу. Победоносное шествие начали диодные и индукционные. Теперь это не просто спираль, которая нагревается и светит. Современная светодиодка это сложный электронный осветительный прибор с блоком питания на микросхемах и высокотехнологичных кристаллах. Некоторые модели комплектуются системами дистанционного управления с пульта, датчиками движения и освещения.
Во времена СССР показателем яркости была мощность лампочки. Сейчас этот показатель отходит на второй план, теперь это значение только примерно характеризует световой поток светодиодных ламп.
Содержание
- 1. В чем измеряется световой поток?
- 2. Виды обмана
- 3. Соответствие светодиодных и накаливания
- 4. Соответствие
- 5. Люминесцентные КЛЛ
- 6. Большие люминесцентные лампочки
- 7. Галогенные
- 8. Регулировка яркости
- 9. ДРЛ и ДНАТ
- 10. Световой поток светодиодных светильников
- 11. Цветовая температура
- 12. Как вычислить светопоток
- 13. Итоги
В чем измеряется световой поток?
Единица измерения светового потока, сокращенно обозначается «лм». Этот параметр характеризует самый важный показатель современной светотехники, количество света от источника. Второй важный показатель, это количество Люмен на 1 Ватт.
Пример эффективности:
- светодиоды имеют от 60 до 200 лм/вт,
- энергосберегайки 60 лм/вт;
- диодные прожектора обычно 80-110 лм/вт.


Единица светового потока не зависит цветовой температуры источника и способа получения света. Это может быть лед кристалл, нить накала или газоразрядная дуга.
Виды обмана
Недобросовестные производители активно пользуются незнанием соответствия люмен и энергопотребления. Например, указывают в характеристиках:
- мощность 7W;
- светопоток 500лм;
- аналог лампы накаливания на 70вт.
Пожилой покупатель ориентируется только на последний пункт, где указан аналог. Светопоток аналогичной 70W должен быть 700-800лм., а не 500лм. После покупки оказывается, что новая лампочка светит хуже, поэтому требуется покупать новые, если покупали сразу комплект для люстры.
Хорошо, если производитель не обманул и указал светопоток честно. Изготовители самой дешевой светотехники завышают параметры своих светильником, лампочек и прожекторов. По результатам моего тестирования реальные мощность и светопоток бывают ниже до 30-40%.
Соответствие светодиодных и накаливания
В качестве соответствия указаны средние значения, они могут меняться примерно +/- 15%. Зависят от типа матового светорассеивателя, конструкции и комплектующих. Чаще всего спрашивают про световой поток лампы накаливания 100 ватт и 60 вт.
Зависят от типа матового светорассеивателя, конструкции и комплектующих. Чаще всего спрашивают про световой поток лампы накаливания 100 ватт и 60 вт.
Световой поток светодиодных ламп таблица
| Накаливания, Вт | Светодиодная, аналог, Вт | Поток света, Лм |
| 25 | 3 | 250 |
| 40 | 5 | 400 |
| 60 | 8 | 650 |
| 100 | 14 | 1300 |
| 150 | 22 | 2100 |
Эффективность светодиодных ламп от 70 до 110 лм/вт, но сильное влияние оказывает матовая колба из поликарбоната. В зависимости от качества на ней теряется от 10% до 30% света.
Для накаливания большую роль играет напряжение в сети 220 вольт. Изменение напряжения с 220В до 230В добавляет 10% к яркости.
Но следует учитывать, что у обычной накаливания свет распространяется на 360 градусов, у диодной около 180 градусов. При установке в люстру или светильник следует учитывать светопропускаемость установленных плафонов. Ситуацию с диодным источником света может улучшить форма плафона, если его отверстие находится напротив колбы. В таком положении меньше света будет теряться внутри и больше выходить наружу.
При установке в люстру или светильник следует учитывать светопропускаемость установленных плафонов. Ситуацию с диодным источником света может улучшить форма плафона, если его отверстие находится напротив колбы. В таком положении меньше света будет теряться внутри и больше выходить наружу.
Соответствие
..
Приличный вклад в запутывание по соответствию мощности и светопотока внесли китайцы. В советские времена светотехника соотвествовала государственным стандартам. Трудолюбивые китайцы стали производить светотехнику по собственным стандартам и импортировать в Россию. Теперь стандартная лампочка на 60W в зависимости от производителя может быть от 500лм до 700лм. По отечественным гостам этот параметр был от 600 до 650лм.
Мне попадались такие китайские, покупал 15 штук самых простых и недорогих. Вроде бы обычная, даже не мог представить, как её можно сделать плохо. В течение 1 месяца всё вышли из строя, у всех отвалилась стеклянная колба, одна даже попала мне в голову, хорошо что не разбилась.
Люминесцентные КЛЛ
У вас дома наиболее распространена разновидность люминесцентных в виде КЛЛ, Компактных Люминесцентных Ламп. В магазинах и быту называются «энергосберегающие КЛЛ». Компактность достигается за счёт скручивания светящейся трубки в спираль.
Так же активно меняют люминесцентные и энергосберегающие лампы на светодиодные. Это относится к классической форме и в виде трубок. При этом для потолочных светильников Армстронг требуется небольшая доработка по удалению ЭПРА.
Световой поток люминесцентных ламп таблица
| Накаливания | КЛЛ | Поток света, Лм |
| 25 вт | 5вт | 250 |
| 40 вт | 9вт | 400 |
| 60 вт | 13вт | 650 |
| 80 вт | 15вт | 900 |
| 100 вт | 20вт | 1300 |
| 150 вт | 35вт | 2100 |
Разновидности КЛЛ
Таблица соотвествия для КЛЛ
| Мощность КЛЛ | Средний показатель в люменах |
| 9 вт | 450лм |
| 11 вт | 550лм |
| 15 вт | 800лм |
| 20 вт | 1200лм |
| 2 5вт | 1500лм |
| 30 вт | 1900лм |
| 35 вт | 2200лм |
| 55 вт | 3900лм |
| 85 вт | 6500лм |
| 105 вт | 6700лм |
| Показатели получены от бренда Osram | |
Благодаря китайцам и экономической ситуации, которая сложилась из-за курса доллара, производители любят завышать параметры светопотока. Такое завышение позволяет выделиться среди других и увеличить продажи. Измерять светопоток сложно и требуется дорогое оборудование, поэтому простой покупатель не раскроет этот обман.
Такое завышение позволяет выделиться среди других и увеличить продажи. Измерять светопоток сложно и требуется дорогое оборудование, поэтому простой покупатель не раскроет этот обман.
Световой поток люминесцентной зависит от её формы, плотная спираль КЛЛ некоторую часть света затеняет и он остаётся внутри спирали. Света бывает больше от простых трубок, которые не имеют сложных форм.
Большие люминесцентные лампочки
К большим относятся люминесцентные трубки длиной 47см и 120см от потолочных и настенных светильников. Обозначаются T5 и T8, цоколь у них G13, G23. Наиболее популярны на 18 вт и 36 вт
При замене на светодиодные трубки учитывайте, что у них может быть матовый рассеиватель. Изготовитель запросто может указать светопоток без этого рассеивателя, на котором теряется 10-20%. Влияет и количество слоёв люминофора на стенках, от него зависит цветовая температура.
Таблица для простых
| Люминесцентные | LED аналог, Ватт | Люмены |
| 10 вт | 5 | 400 |
| 15 вт | 8 | 700 |
| 16 вт | 9 | 800 |
| 18 вт | 11 | 900 |
| 23 вт | 15 | 1350 |
| 30 вт | 20 | 1800 |
| 36 вт | 23 | 2150 |
| 38 вт | 25 | 2300 |
| 58вт | 35 | 3350 |
| Информация получена с официального сайта Osram для серии Стандарт | ||
Кроме недорогих бюджетных моделей производятся и дорогие улучшенные. Цена отличается значительно, но это окупается повышенной светоотдачей, которая больше на 50%. Светоотдача улучшенных моделей на 58вт получается, как у светодиодов, 90 лм/вт. Недостатком является высокое реактивное потребление энергии, которое зависит от показателя «коэффициент мощности».
Цена отличается значительно, но это окупается повышенной светоотдачей, которая больше на 50%. Светоотдача улучшенных моделей на 58вт получается, как у светодиодов, 90 лм/вт. Недостатком является высокое реактивное потребление энергии, которое зависит от показателя «коэффициент мощности».
Таблица для улучшенных
| Люминесцентные | LED аналог, Ватт | Люмены |
| 10 вт | 7 | 650 |
| 15 вт | 10 | 950 |
| 16 вт | 14 | 1250 |
| 18 вт | 15 | 1350 |
| 23 вт | 20 | 1900 |
| 30 вт | 25 | 2400 |
| 36 вт | 35 | 3350 |
| 38 вт | 35 | 3300 |
| 58 вт | 55 | 5200 |
| Информация получена с официального сайта Osram для серии Стандарт | ||
Срок службы обычных 15-20 тыс. часов, но есть модели со сроком работы в 75.000 – 90.000 часов, например из серии Osram LUMILUX XXT T8.
часов, но есть модели со сроком работы в 75.000 – 90.000 часов, например из серии Osram LUMILUX XXT T8.
Еще один из существенных недостатков, это снижение светового потока при низких температурах. Давление в трубке снижается и светоотдача уменьшается.
Для потолочных светильников Армстронг обычно указывают потребление энергии и световой поток, например 36W и 2800лм. Производитель умалчивает, что 2800лм это светопоток ламп без самого светильника. Ведь в нём одна сторона трубки светит в корпус, другая в помещение. Чтобы свет на стенке не терялся, ставят отражатель. Но он расположен близко к трубке, поэтому корпус трубки затеняет часть отраженного света от 15 до 20%. Поэтому реальное количество люмен для светильника Армстронг ниже, вместо 2800лм будет только 2200лм.
У светодиодных трубок T5 T8 такой проблемы нет, отражатель не требуется. Светодиоды установлены с одной стороны и светят только в сторону помещения.
Галогенные
В миниатюрных источниках освещения, таких как точечные для потолка, устанавливали галогенные лампы. Галогенка имеет минимальные размеры по сравнению с другими. Чаще всего это цоколь G9, с которым сейчас больше всего проблем. Яркость диодных зависит от размера системы охлаждения. Чтобы сделать светодиодку размером с галогенку, охлаждение приходится делать очень маленьким. Поэтому мощность диодных с цоколем G9 не превышает 300лм. В характеристиках часто завышают, указывая 400-600лм, так же не верьте параметрам на базаре Aliexpress. При использовании люстры на 6 патронов и 300лм на одну светодиодку невозможно получить хорошее освещение, придётся менять люстру.
Галогенка имеет минимальные размеры по сравнению с другими. Чаще всего это цоколь G9, с которым сейчас больше всего проблем. Яркость диодных зависит от размера системы охлаждения. Чтобы сделать светодиодку размером с галогенку, охлаждение приходится делать очень маленьким. Поэтому мощность диодных с цоколем G9 не превышает 300лм. В характеристиках часто завышают, указывая 400-600лм, так же не верьте параметрам на базаре Aliexpress. При использовании люстры на 6 патронов и 300лм на одну светодиодку невозможно получить хорошее освещение, придётся менять люстру.
Таблица соответствия для простых галогенных
| Галогенная | LED аналог | Люмены |
| 5 вт | 2 | 60 |
| 10 вт | 3 | 140 |
| 25 вт | 4 | 260 |
| 35 вт | 5 | 410 |
| 40 вт | 6 | 490 |
| 50 вт | 9 | 700 |
| Данные с сайта Осрам | ||
Разновидности галогенных
Таблица соответствия для улучшенных галогенок
| Галогенная | LED аналог | Люмены |
| 10 вт | 3 | 180 |
| 20 вт | 4 | 350 |
| 25 вт | 6 | 500 |
| 30 вт | 5 | 650 |
| 35 вт | 8 | 860 |
| 40 вт | 12 | 980 |
| 50 вт | 14 | 1200 |
| Использованы значения с официального сайта Osram для галогенок серии Standart с цоколями G4 и G9 | ||
Срок службы в среднем от 1000ч до 2000ч. Чем выше мощность, тем короче срок службы.
Чем выше мощность, тем короче срок службы.
Регулировка яркости
Устройство для регулирования мощности светового потока называется диммером.С ним совместимы только лампы накаливания, галогенки и некоторые светодиодные. Для диодных источников должен быть установлен специальный диммер, отличается минимальной мощностью , у обычного от 30W, у светодиодного диммера от 1W. Это связано с низким энергопотреблением светодиодок.
Люминесцентные и другие с блоками розжига регулирование не поддерживают. Им требуется постоянное напряжение для работы.
ДРЛ и ДНАТ
В промышленном и уличном освещении применяются лампы ДРЛ и ДНАТ, которые имеют приличную светоотдачу. При таком большом энергопотреблении используется цоколь Е40, E40.
- ДНАТ- это дуговые натриевые трубчатые;
- ДРЛ – это дуговая ртутная люминесцентная.
Эффективность лм/ватт у них на уровне простых светодиодов, но срок службы в 3-4 раза ниже. К тому же светопоток снижается быстрее, чем у диодного освещения.
Таблица аналогов для натриевых
| ДНАТ | Светодиодный аналог | Люмены |
| ДНАТ 70 | 50вт | 4.600 |
| ДНАТ 100 | 75вт | 7.300 |
| ДНАТ 150 | 110вт | 11.000 |
| ДНАТ 250 | 190вт | 19.000 |
| ДНАТ 400 | 350вт | 35.000 |
Таблица аналогов для ртутных
| ДРЛ | Светодиодный аналог | Люмены |
| ДРЛ 125 | 65вт | 6000 |
| ДРЛ 250 | 150вт | 13000 |
| ДРЛ 400 | 250вт | 23000 |
| ДРЛ 700 | 450вт | 40000 |
| ДРЛ 1000 | 600вт | 58000 |
У современных светодиодных аналогов светильников на хороших диодах, например Osram Duris, срок службы около 100.000ч. Из строя быстрее выйдет блок питания, чем LED чипы. Хороший блок питания (драйвер, преобразователь) на японских комплектующих служит до 70.000ч. Многое зависит от конденсаторов, которые теряются свою емкость и параметры питания светодиодов меняются.
Из строя быстрее выйдет блок питания, чем LED чипы. Хороший блок питания (драйвер, преобразователь) на японских комплектующих служит до 70.000ч. Многое зависит от конденсаторов, которые теряются свою емкость и параметры питания светодиодов меняются.
Световой поток светодиодных светильников
Большинство современных уличных и промышленных светильников сейчас производят на качественных светодиодах, типа NationStar, Osram Duris, Cree, LG, Samsung. Светоотдача у них на уровне 110 -120 лм/ватт.
Промышленные, уличные, производственные
| Энергопотребление, ватт | Люмены |
| 10вт | 1100-1200лм |
| 20вт | 2200-2400лм |
| 30вт | 3300-3600лм |
| 50вт | 5500-6000лм |
| 70вт | 7700-8400лм |
| 100вт | 11000-12000лм |
Бытовые прожекторы для дома
| Энергопотребление, ватт | Люмены |
| 10вт | 800-900лм |
| 20вт | 1600-1800лм |
| 30вт | 2400-2700лм |
| 50вт | 4000-4500лм |
| 70вт | 5600-6300лм |
| 100вт | 8000-9000лм |
Для бытовых прожекторов ставят LED диоды попроще, условия эксплуатации у них легче и заменить гораздо проще. Китайцы как всегда экономят и могут ставить низкокачественные ЛЕДы. Мне попадались светильники для квартиры и других жилых помещений с эффективностью 60 лм. на ватт, вместо обычных 80-90 лм/вт. По замерам получалось, что он потребляет энергии прилично, но не светит. Сперва подумал оборудование сломалось, но после калибровки ничего не изменилось, освещение было плохое.
Китайцы как всегда экономят и могут ставить низкокачественные ЛЕДы. Мне попадались светильники для квартиры и других жилых помещений с эффективностью 60 лм. на ватт, вместо обычных 80-90 лм/вт. По замерам получалось, что он потребляет энергии прилично, но не светит. Сперва подумал оборудование сломалось, но после калибровки ничего не изменилось, освещение было плохое.
Цветовая температура
Большинство из нас привыкли к теплому свету источников с нитью накаливания, переходить на нейтрально белый на хотят Как показывает практика, стоит попробовать в течение двух дней нейтрально белый свет, и потом уже никто не соглашается переходить обратно на теплый.
На фото вы видите наглядные примеры отличия цветовой температуры:
- теплый 2700К, 2900К;
- нейтральный белый 4000К;
- слегка холодноватый 5500К и 6000К.
Как вычислить светопоток
Чтобы узнать сколько люмен у источника, используйте средние значения светоотдачи:
- для диодных умножьте мощность на 80-90 лм/вт для лампочек с матовой колбой и получите светопоток;
- для диодных филаментных умножайте энергопотребление на 100 лм/вт, филаментные прозрачные с диодами в виде желтых полосок;
- люминесцентные КЛЛ умножайте на 60 лм/вт, для дорогих он может быть выше, но они гораздо быстрее теряют яркость, поэтому это значение будет более точным;
- ДНАТ 66 лм/вт для 70W, 74 лм/вт для 100W 150W 250W, 88 лм/вт у 400W;
- ДРЛ множитель будет 58 лм/вт при среднем срок службы от 12 до 18т часов, китайские могут иметь другие характеристики, они всегда умудряются сэкономить даже там, где это практически не возможно.

Итоги
Как вы видите из выше расположенных таблиц, бывают простые и улучшенные модели лампочек. Перед тем как их менять на новые, узнайте точную модель старых, обычно маркировка нанесена на корпусе. По маркировке ищите в сети сайт производителя, на котором должна находится информация о технических характеристиках. Если этого не сделать, то новое освещение может оказаться хуже старого.
Другой вариант, это взять образец с собой в магазин и показать продавцу, некоторые из них хорошо разбираются в этом. В случае с интернет-магазином, отправьте фото консультанту по электронной почте.
у меня для вас плохие новости / Хабр
Все, кто разбирался с качеством света светодиодных ламп и все, кто читал мои статьи о светодиодных лампах, знают о таком параметре, как индекс цветопередачи (CRI, он же Ra). Считается, что у качественного света для жилых помещений CRI должен быть не меньше 80.
Недавно я столкнулся с лампой, CRI у которой был вполне приличным — 83. 4, но она давала очень неприятный зеленоватый свет.
4, но она давала очень неприятный зеленоватый свет.
Я попытался разобраться, что с ней не так.
Индекс цветопередачи или colour rendering index — CRI (
ru.wikipedia.org/wiki/Индекс_цветопередачи
) — параметр, характеризующий уровень соответствия естественного цвета тела видимому (кажущемуся) цвету этого тела при освещении его данным источником света был предложен в 1965 году.
CRI — это средний уровень передачи восьми цветов R1-R8.
Иногда, помимо CRI, указывается и измеряется индекс передачи красного цвета R9. Этот индекс влияет на качество передачи тона человеческой кожи. На lamptest.ru измеренный индекс R9 указан в карточке каждой лампы.
Ещё в 2007 году Международная комиссия по освещению отметила, что «…индекс цветопередачи, обычно неприменим для прогнозирования параметров цветопередачи набора источников света, если в этот набор входят светодиоды белого цвета», однако так вышло, что все производители светодиодных ламп используют именно CRI.
В 2010 году, для более точной оценки качества передачи цвета, была разработана методика Color Quality Scale (CQS), оценивающая качество света по пятнадцати цветам.
В 2015 года был разработан стандарт ТМ-30-15, который оценивает качество света по 99 цветам.
У хороших ламп значения всех трёх индексов приблизительно равны.
А теперь вернёмся к лампе Gauss 207707102 190Lm 2W 2700K G4 12V, из-за которой я и затеял всё это исследование. Её цветовые индексы выглядят удивительно.
Значение CRI достаточно высокое — 83.4, TM30 Rf — 84.3, а вот CQS очень низкий — 35.8. Похоже, хитрый китаец намешал люминофор так, чтобы хорошо передавались именно те 8 цветов, которые учитываются при измерении CRI. Удивительно, что результат вроде как самого продвинутого индекса TM30 также оказался высоким.
Замечу, что из всех 1244 ламп, параметры которых я измерял, только у одной оказался такой низкий уровень индекса CQS. Даже у самых плохих безымянных китайских лампочек с CRI 60, CQS составляет не менее 50.
Я начал изучать значения CQS у ламп и выяснил, что встречается довольно много ламп, у которых CRI больше 80, а значение CQS составляет чуть выше 70, но свет таких ламп визуально вполне комфортный. А вот у некоторых ламп с CRI больше 80, CQS оказался около 60 и свет таких ламп визуально зеленоватый или желтоватый.
А вот у некоторых ламп с CRI больше 80, CQS оказался около 60 и свет таких ламп визуально зеленоватый или желтоватый.
Возникает вопрос, что с этим всем делать. Наверное придётся добавлять на lamptest значение CQS и учитывать его при расчёте итоговой оценки ламп, чтобы не могло получится, что лампа с высоким CRI, но некомфортным светом получала высокую оценку.
P.S: Для развития проекта lamptest.ru ищу
1. PHP-программиста, готового помочь с доработкой сайта.
2. Помощников, готовых заниматься покупкой и возвратом ламп в магазинах.
3. Лаборатории с фотометрическим шаром, готовые бесплатно измерить световой поток десятка моих образцов (для подтверждения точности моих измерений).
4. Человека, делавшего формулу расчёта оценки качества ламп в Excel (всё перелопатил, не могу найти контакты).
© 2017, Алексей Надёжин
Электротехническая продукция в Уфе
|
Торговая сеть ATOM electric работает на рынке электротехнической продукции с 2003 года и предлагает своим клиентам товары оптимального соотношения цена-качество.
|
|
 )
) )
) ) IP20, 23, 40
) IP20, 23, 40 3, GU5.3
3, GU5.3 )
)
 ) IP31
) IP31 ) серия КСРМ сборно-разборные IP31
) серия КСРМ сборно-разборные IP31 ), наборы
), наборы

Вакцина
мРНК для иммунотерапии рака | Молекулярный рак

Pantin J, Battiwalla M. Опрокидывание яблока CAR-T (Т-клеточная терапия с химерным антигенным рецептором) – устойчивость требует инноваций в США. Бр Дж Гематол. 2020;190(6):851–3.
ПабМед
Статья
Google Scholar
Hargadon KM, Johnson CE, Williams CJ. Терапия блокадой иммунных контрольных точек при раке: обзор одобренных FDA ингибиторов иммунных контрольных точек.Int Immunopharmacol. 2018;62:29–39.
КАС
пабмед
Статья
Google Scholar
Guo C, Manjili MH, Subjeck JR, Sarkar D, Fisher PB, Wang XY. Терапевтические противораковые вакцины: прошлое, настоящее и будущее. Adv Рак Res. 2013; 119: 421–75.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Чивер М.А., Хигано К.С. PROVENGE (Sipuleucel-T) при раке предстательной железы: первая терапевтическая противораковая вакцина, одобренная FDA. Клин Рак Рез. 2011;17(11):3520–6.
Клин Рак Рез. 2011;17(11):3520–6.
ПабМед
ПабМед Центральный
Статья
Google Scholar
Папахристофилу А., Хипп М.М., Клинкхардт У., Фрух М., Себастьян М., Вайс С. и др. Оценка фазы Ib самоадъювантной иммунотерапии активного рака на основе мРНК на основе протамина, BI1361849 (CV9202), в сочетании с местной лучевой терапией у пациентов с немелкоклеточным раком легкого IV стадии. J Иммунный рак. 2019;7(1):38.
ПабМед
ПабМед Центральный
Статья
Google Scholar
Rittig SM, Haentschel M, Weimer KJ, Heine A, Muller MR, Brugger W, et al. Долгосрочная выживаемость коррелирует с иммунологическим ответом у пациентов с почечно-клеточным раком, получавших иммунотерапию на основе мРНК. Онкоиммунология. 2016;5(5):e1108511.
ПабМед
Статья
КАС
Google Scholar
Фагфури Э., Пурфарзи Ф. , Фагфури А.Х., Абдоли Шадбад М., Хаджиасгарзаде К., Барадаран Б. Последние разработки вакцин на основе РНК в иммунотерапии рака. Мнение Эксперта Биол Тер. 2020: 1–8.
, Фагфури А.Х., Абдоли Шадбад М., Хаджиасгарзаде К., Барадаран Б. Последние разработки вакцин на основе РНК в иммунотерапии рака. Мнение Эксперта Биол Тер. 2020: 1–8.
Van Nuffel AM, Wilgenhof S, Thielemans K, Bonehill A. Преодоление ограничения HLA в клинических испытаниях: иммунный мониторинг терапии DC, нагруженной мРНК. Онкоиммунология. 2012;1(8):1392–4.
ПабМед
ПабМед Центральный
Статья
Google Scholar
Вольф Дж. А., Мэлоун Р. В., Уильямс П., Чонг В., Аксади Г., Яни А. и др. Прямой перенос генов в мышцы мышей in vivo. Наука. 1990; 247 (4949, часть 1): 1465–8.
КАС
пабмед
Статья
Google Scholar
Son S, Nam J, Zenkov I, Ochyl LJ, Xu Y, Scheetz L, et al. Сахарные нанокапсулы с отпечатанными микробными молекулярными структурами для вакцинации мРНК. Нано Летт. 2020;20(3):1499–509.
КАС
пабмед
Статья
Google Scholar

Pardi N, Hogan MJ, Porter FW, Weissman D. мРНК-вакцины – новая эра в вакцинологии. Nat Rev Drug Discov. 2018;17(4):261–79.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Блум К., ван ден Берг Ф., Арбутнот П. Самоамплифицирующиеся РНК-вакцины от инфекционных заболеваний. Джин Тер. 2020: 1–13.
Флемминг А. Вакцины: самоамплифицирующаяся РНК в липидных наночастицах: вакцина нового поколения? Nat Rev Drug Discov.2012;11(10):748–9.
КАС
пабмед
Статья
Google Scholar
Шахин У., Муйк А., Дерхованесян Э., Фоглер И., Кранц Л.М., Вормер М. и др. Вакцина против COVID-19 BNT162b1 вызывает реакции человеческих антител и Т-клеток Th2. Природа. 2020; 586 (7830): 594–9.
КАС
пабмед
Статья
Google Scholar
Джексон Л.А., Робертс П. С., Грэм Б.С. Вакцина мРНК SARS-CoV-2 — предварительный отчет.Ответить N Engl J Med. 2020;383(12):1191–2.
С., Грэм Б.С. Вакцина мРНК SARS-CoV-2 — предварительный отчет.Ответить N Engl J Med. 2020;383(12):1191–2.
КАС
пабмед
Google Scholar
Кобб М. Кто открыл информационную РНК? Карр Биол. 2015;25(13):R526–32.
КАС
пабмед
Статья
Google Scholar
Linares-Fernandez S, Lacroix C, Exposito JY, Verrier B. Адаптация мРНК-вакцины для уравновешивания врожденного/адаптивного иммунного ответа. Тренды Мол Мед. 2020;26(3):311–23.
КАС
пабмед
Статья
Google Scholar
Kranz LM, Diken M, Haas H, Kreiter S, Loquai C, Reuter KC, et al. Системная доставка РНК в дендритные клетки использует противовирусную защиту для иммунотерапии рака. Природа. 2016; 534(7607):396–401.
ПабМед
Статья
КАС
Google Scholar

Пулит-Пеналоза Ю.А., Щербик С.В., Бринтон М.А. Для активации экспрессии гена Oas1a с помощью IFN типа I требуются как STAT1, так и STAT2, тогда как для активации Oas1b требуется только STAT2.Вирусология. 2012;425(2):71–81.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Кумар П., Суини Т.Р., Скабкин М.А., Скабкина О.В., Хеллен К.У., Пестова Т.В. Ингибирование трансляции членами семейства IFIT определяется их способностью избирательно взаимодействовать с 5′-концевыми областями cap0-, cap1- и 5’ppp-мРНК. Нуклеиновые Кислоты Res. 2014;42(5):3228–45.
КАС
пабмед
Статья
Google Scholar
De Beuckelaer A, Grooten J, De Koker S. Интерфероны типа I модулируют CD8(+) Т-клеточный иммунитет к мРНК-вакцинам. Тренды Мол Мед. 2017;23(3):216–26.
ПабМед
Статья
КАС
Google Scholar

Broos K, Van der Jeught K, Puttemans J, Goyvaerts C, Heirman C, Dewitte H, et al. Опосредованная частицами внутривенная доставка мРНК антигена приводит к сильным антиген-специфическим Т-клеточным ответам, несмотря на индукцию интерферона I типа.Молекулярные нуклеиновые кислоты. 2016;5(6):e326.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
De Beuckelaer A, Pollard C, Van Lint S, Roose K, Van Hoecke L, Naessens T, et al. Интерфероны типа I нарушают способность вакцин мРНК Lipoplex вызывать цитолитические Т-клеточные ответы. Мол Тер. 2016;24(11):2012–20.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar
Поллард С., Рейман Дж., Де Хаес В., Верриер Б., Ван Галк Э., Нэссенс Т. и другие. IFN типа I противодействует индукции антиген-специфических иммунных ответов путем доставки мРНК-вакцин на основе липидов. Мол Тер. 2013;21(1):251–9.
2013;21(1):251–9.
КАС
пабмед
Статья
Google Scholar
Miao L, Li L, Huang Y, Delcassian D, Chahal J, Han J, et al. Доставка мРНК-вакцин с гетероциклическими липидами повышает противоопухолевую эффективность за счет STING-опосредованной активации иммунных клеток.Нац биотехнолог. 2019;37(10):1174–85.
КАС
пабмед
Статья
Google Scholar
Oberli MA, Reichmuth AM, Dorkin JR, Mitchell MJ, Fenton OS, Jaklenec A, et al. Липидные наночастицы способствовали доставке мРНК для мощной иммунотерапии рака. Нано Летт. 2017;17(3):1326–35.
КАС
пабмед
Статья
Google Scholar
Муттах Ф., Мутманн Н., Рентмайстер А.Синтетическое кэпирование мРНК. Beilstein J Org Chem. 2017;13(1):2819–32.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Шуман С. Каталитическая активность субъединиц фермента, покрывающих мРНК коровьей оспы, коэкспрессируется в Escherichia coli. Дж. Биол. Хим. 1990;265(20):11960–6.
КАС
пабмед
Статья
Google Scholar
Fuchs A-L, Neu A, Sprangers R.Общий метод быстрого и экономичного крупномасштабного производства 5′-кэпированной РНК. РНК. 2016;22(9):1454–66.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Рыдзик А.М., Кулис М., Лукашевич М., Ковальска Дж., Зуберек Дж., Дарзынкевич З.М., и соавт. Синтез и свойства аналогов кэпа мРНК, содержащих имидодифосфатный фрагмент, которые в значительной степени имитируют структуру природного кэпа, но устойчивы к ферментативному гидролизу.Биоорг Мед Хим. 2012;20(5):1699–710.
КАС
пабмед
Статья
Google Scholar
Шлаке Т.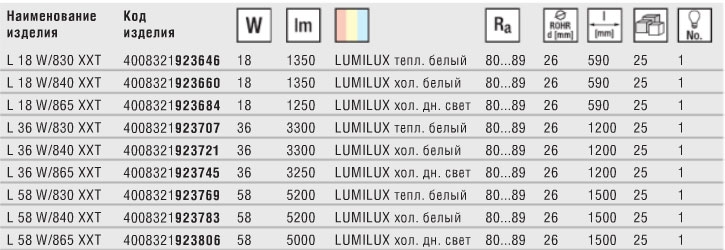 , Тесс А., Фотин-Млечек М., Каллен К.Дж. Разработка технологий мРНК-вакцины. РНК биол. 2012;9(11):1319–30.
, Тесс А., Фотин-Млечек М., Каллен К.Дж. Разработка технологий мРНК-вакцины. РНК биол. 2012;9(11):1319–30.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Vaidyanathan S, Azizian KT, Haque AKMA, Henderson JM, Hendel A, Shore S, et al.Истощение уридина и химическая модификация повышают активность мРНК Cas9 и снижают иммуногенность без очистки ВЭЖХ. молярные тер-нуклеиновые кислоты. 2018;12:530–42.
КАС
Статья
Google Scholar
Дженсен С., Томсен А.Р. Распознавание РНК-вирусов: обзор рецепторов врожденного иммунитета, участвующих в распознавании инвазии РНК-вируса. Дж Вирол. 2012;86(6):2900–10.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Ringeard M, Marchand V, Decroly E, Motorin Y, Bennasser Y. FTSJ3 представляет собой РНК-2′-O-метилтрансферазу, рекрутируемую ВИЧ, чтобы избежать врожденного иммунного восприятия. Природа. 2019;565(7740):500-+.
Природа. 2019;565(7740):500-+.
КАС
пабмед
Статья
Google Scholar
Cao J, He LJ, Lin GY, Hu CQ, Dong R, Zhang J, et al. Кэп-зависимый фактор инициации трансляции, eIF4E, является мишенью оуабаин-опосредованного ингибирования HIF-1 альфа. Биохим Фармакол.2014;89(1):20–30.
КАС
пабмед
Статья
Google Scholar
Whisenand J, Azizian K, Henderson J, Shore S, Shin D, Lebedev A, et al. Соображения по дизайну и производству cGMP терапевтических средств с мРНК. Плакат Trilink Biotechnol. https://www.trilinkbiotech.com/media/contentmanager/content/mRNA_OTS1.pdf.
Weissman D. Терапия транскриптом мРНК. Эксперт Rev Вакцины. 2015;14(2):265–81.
КАС
пабмед
Статья
Google Scholar
Орландини фон Ниссен А.Г., Полеганов М.А., Рехнер С., Плашке А., Кранц Л. М., Фессер С. и соавт. Улучшение доставки терапевтического гена на основе мРНК с помощью 3′-UTR, увеличивающих экспрессию, идентифицированных скринингом клеточной библиотеки. Мол Тер. 2019;27(4):824–36.
М., Фессер С. и соавт. Улучшение доставки терапевтического гена на основе мРНК с помощью 3′-UTR, увеличивающих экспрессию, идентифицированных скринингом клеточной библиотеки. Мол Тер. 2019;27(4):824–36.
КАС
пабмед
Статья
Google Scholar
Цзя Л., Мао И., Цзи К., Дерш Д., Юделл Д.В., Цянь С.Б.Расшифровка транслируемости и стабильности мРНК из 5’UTR. Nat Struct Mol Biol. 2020;27(9):814–21.
КАС
пабмед
Статья
Google Scholar
Thess A, Grund S, Mui BL, Hope MJ, Baumhof P, Fotin-Mleczek M, et al. Сконструированная последовательность мРНК без химических модификаций нуклеозидов обеспечивает эффективную белковую терапию у крупных животных. Мол Тер. 2015;23:С55–С.
Артикул
Google Scholar
Лима С.А., Чипман Л.Б., Николсон А.Л., Чен Ю.Х., Йи Б.А., Йео Г.В. и др. Короткие поли(а)-хвосты являются консервативным признаком высокоэкспрессированных генов. Nat Struct Mol Biol. 2017;24(12):1057–63.
Nat Struct Mol Biol. 2017;24(12):1057–63.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Willis E, Pardi N, Parkhouse K, Mui BL, Tam YK, Weissman D, et al. Вакцинация мРНК с модифицированными нуклеозидами частично преодолевает ингибирование материнскими антителами иммунных ответов de novo у мышей.Sci Transl Med. 2020;12(525):eaav5701.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Sharifnia Z, Bandehpour M, Kazemi B, Zarghami N. Дизайн и разработка модифицированной мРНК, кодирующей коровый антиген вируса гепатита С: возможное применение в производстве вакцин. Иран Биомед Дж. 2019;23(1):57–67.
ПабМед
ПабМед Центральный
Статья
Google Scholar
Pardi N, Weissman D. Нуклеозидно-модифицированные мРНК-вакцины против инфекционных заболеваний. Методы Мол Биол. 2017;1499:109–21.
Методы Мол Биол. 2017;1499:109–21.
КАС
пабмед
Статья
Google Scholar
О С, Кесслер Дж.А. Дизайн, сборка, производство и трансфекция синтетических модифицированных мРНК. Методы. 2018; 133:29–43.
КАС
пабмед
Статья
Google Scholar
Карико К., Мурамацу Х., Людвиг Дж., Вайсман Д.Генерация оптимальной мРНК для терапии: очистка ВЭЖХ устраняет иммунную активацию и улучшает трансляцию нуклеозид-модифицированной мРНК, кодирующей белок. Нуклеиновые Кислоты Res. 2011;39(21):e142.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Карико К., Бакштейн М., Ни Х., Вайсман Д. Подавление распознавания РНК толл-подобными рецепторами: влияние модификации нуклеозидов и эволюционное происхождение РНК.Иммунитет. 2005;23(2):165–75.
КАС
пабмед
Статья
Google Scholar
Kariko K, Muramatsu H, Welsh FA, Ludwig J, Kato H, Akira S, et al. Включение псевдоуридина в мРНК дает превосходный неиммуногенный вектор с повышенной трансляционной способностью и биологической стабильностью. Мол Тер. 2008; 16 (11): 1833–40.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Аранго Д., Стургилл Д., Альхусаини Н., Диллман А.А., Свит Т.Дж., Хэнсон Г. и др. Ацетилирование цитидина в мРНК способствует эффективности трансляции. Клетка. 2018;175(7):1872-+.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Wang H, Hu X, Huang M, Liu J, Gu Y, Ma L, et al. Mettl3-опосредованное метилирование мРНК m(6)a способствует активации дендритных клеток. Нац коммун. 2019; 10(1):1898.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar

Вайсман Д., Парди Н., Мурамацу Х., Карико К. ВЭЖХ-очистка транскрибированной in vitro длинной РНК. Методы Мол Биол. 2013; 969:43–54.
КАС
пабмед
Статья
Google Scholar
Байерсдорфер М., Борос Г., Мурамацу Х., Махини А., Влаткович И., Шахин У. и др. Простой метод удаления примеси дцРНК из транскрибируемой in vitro мРНК. молярные тер-нуклеиновые кислоты. 2019;15:26–35.
Артикул
КАС
Google Scholar
Shivalingam A, Taemaitree L, El-Sagheer AH, Brown T. Скварамиды и мочевины: гибкий подход к полимеразо-совместимой сборке нуклеиновых кислот. Angew Chem Int Ed Eng. 2020;59(28):11416–22.
КАС
Статья
Google Scholar
Hassett KJ, Benenato KE, Jacquinet E, Lee A, Woods A, Yuzhakov O, et al. Оптимизация липидных наночастиц для внутримышечного введения мРНК-вакцин. молярные тер-нуклеиновые кислоты. 2019; 15:1–11.
2019; 15:1–11.
КАС
Статья
Google Scholar
Ислам М.А., Райс Дж., Рисор Э., Зоуп Х., Тао В., Лим М. и др. Наночастицы мРНК-вакцины с импульсным адъювантом для иммунопрофилактического и терапевтического подавления опухолей у мышей. Биоматериалы. 2021;266:120431.
КАС
пабмед
Статья
Google Scholar
Луо М., Ван Х., Ван З., Цай Х., Лу З., Ли И. и др. STING-активирующая нановакцина для иммунотерапии рака.Нац Нанотехнолог. 2017;12(7):648–54.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Miao L, Lin J, Huang Y, Li L, Delcassian D, Ge Y, et al. Синергетические липидные композиции для опосредованной рецептором альбумина доставки мРНК в печень. Нац коммун. 2020;11(1):2424.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Де Кирсмекер Б., Клаерхаут С., Карраско Дж., Бар И., Корталс Дж., Вильгенхоф С. и др. TriMix и мРНК опухолевого антигена, электропорация вакцины дендритных клеток плюс ипилимумаб: связь между активацией Т-клеток и клиническими ответами при распространенной меланоме. J Иммунный рак. 2020;8(1):e000329.
ПабМед
ПабМед Центральный
Статья
Google Scholar
Bonehill A, Tuyaerts S, Van Nuffel AM, Heirman C, Bos TJ, Fostier K, et al.Повышение стимулирующей способности Т-клеток дендритных клеток человека путем совместной электропорации с CD40L, CD70 и конститутивно активной TLR4, кодирующей мРНК. Мол Тер. 2008;16(6):1170–80.
КАС
пабмед
Статья
Google Scholar
Hewitt SL, Bai A, Bailey D, Ichikawa K, Zielinski J, Karp R, et al. Устойчивый противораковый иммунитет при внутриопухолевом введении мРНК IL-23, IL-36gamma и OX40L. Sci Transl Med. 2019;11(477):eaat9143.
2019;11(477):eaat9143.
КАС
пабмед
Статья
Google Scholar
Брито Л.А., Коммаредди С., Майоне Д., Уэмацу Ю., Джовани С., Берланда Скорца Ф. и др. Самоамплифицирующиеся мРНК-вакцины. Ад Генет. 2015; 89: 179–233.
КАС
пабмед
Статья
ПабМед Центральный
Google Scholar
Johanning FW, Conry RM, LoBuglio AF, Wright M, Sumerel LA, Pike MJ, et al. Полинуклеотидный вектор мРНК вируса Синдбис обеспечивает пролонгированную экспрессию гетерологичного гена высокого уровня in vivo.Нуклеиновые Кислоты Res. 1995;23(9):1495–501.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Райнер Ю.О., Дрыга С.А., Камруд К.И. Альфавирусные переносчики и вакцинация. Преподобный Мед Вирол. 2002;12(5):279–96.
КАС
пабмед
Статья
Google Scholar

Циммер Г. РНК-репликоны – новый подход к иммунопрофилактике вирусов гриппа.Вирусы. 2010;2(2):413–34.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Hekele A, Bertholet S, Archer J, Gibson DG, Palladino G, Brito LA, et al. Быстро производимая вакцина SAM((R)) против гриппа H7N9 является иммуногенной для мышей. Новые микробы заражают. 2013;2(8):e52.
ПабМед
ПабМед Центральный
Google Scholar
Bernstein DI, Reap EA, Katen K, Watson A, Smith K, Norberg P, et al.Рандомизированное двойное слепое испытание фазы 1 вакцины с репликоном альфавируса против цитомегаловируса у взрослых добровольцев, не инфицированных ЦМВ. вакцина. 2009;28(2):484–93.
КАС
пабмед
Статья
Google Scholar
Лундстрем К. Самореплицирующиеся РНК-вирусы для РНК-терапии. Молекулы. 2018;23(12):3310.
2018;23(12):3310.
Центральный пабмед
Статья
КАС
пабмед
Google Scholar
Vogel AB, Lambert L, Kinnear E, Busse D, Erbar S, Reuter KC, et al. Самоамплифицирующиеся РНК-вакцины обеспечивают такую же защиту от гриппа, как и мРНК-вакцины, но в гораздо меньших дозах. Мол Тер. 2018;26(2):446–55.
КАС
пабмед
Статья
Google Scholar
Blakney AK, Zhu Y, McKay PF, Bouton CR, Yeow J, Tang J, et al. Большой — это красиво: улучшенная доставка saRNA и иммуногенность за счет биоразлагаемого катионного полимера с более высокой молекулярной массой.АКС Нано. 2020;14(5):5711–27.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Geall AJ, Verma A, Otten GR, Shaw CA, Hekele A, Banerjee K, et al. Невирусная доставка самоамплифицирующихся РНК-вакцин. Proc Natl Acad Sci U S A. 2012;109(36):14604–9.
2012;109(36):14604–9.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Manara C, Brazzoli M, Piccioli D, Taccone M, D’Oro U, Maione D, et al.Совместное введение РНК, экспрессирующей GM-CSF, является мощным инструментом для повышения эффективности вакцин на основе SAM. вакцина. 2019;37(30):4204–13.
КАС
пабмед
Статья
Google Scholar
Лу Г., Андерлуцци Г., Шмидт С.Т., Вудс С., Галлорини С., Браззоли М. и др. Доставка самоамплифицирующихся мРНК-вакцин с помощью катионных липидных наночастиц: влияние выбора катионных липидов. J Управление выпуском. 2020;325:370–9.
КАС
пабмед
Статья
Google Scholar
Андерлуцци Г., Лу Г., Галлорини С., Браззоли М., Джонсон Р., О’Хаган Д.Т. и др. Изучение влияния конструкции системы доставки на эффективность самоамплифицирующихся РНК-вакцин. Вакцины (Базель). 2020;8(2):212.
Вакцины (Базель). 2020;8(2):212.
КАС
Статья
Google Scholar
Beissert T, Perkovic M, Vogel A, Erbar S, Walzer KC, Hempel T, et al. Стратегия трансамплифицирующей РНК-вакцины для индукции мощного защитного иммунитета. Мол Тер. 2020;28(1):119–28.
КАС
пабмед
Статья
Google Scholar
Ковальски П.С., Рудра А., Мяо Л., Андерсон Д.Г. Доставка мессенджера: достижения в технологиях доставки терапевтической мРНК. Мол Тер. 2019;27(4):710–28.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Крампс Т., Элберс К. Введение в РНК-вакцины. Методы Мол Биол.2017; 1499:1–11.
КАС
пабмед
Статья
Google Scholar
Zeng C, Zhang C, Walker PG, Dong Y. Технологии приготовления и доставки мРНК-вакцин. В кн.: Актуальные проблемы микробиологии и иммунологии. Берлин, Гейдельберг: Springer; 2020 г. https://doi.org/10.1007/82_2020_2172020_217.
В кн.: Актуальные проблемы микробиологии и иммунологии. Берлин, Гейдельберг: Springer; 2020 г. https://doi.org/10.1007/82_2020_2172020_217.
Semple SC, Akinc A, Chen J, Sandhu AP, Mui BL, Cho CK, et al. Рациональный дизайн катионных липидов для доставки миРНК.Нац биотехнолог. 2010;28(2):172–176.
КАС
пабмед
Статья
Google Scholar
Jayaraman M, Ansell SM, Mui BL, Tam YK, Chen JX, Du XY, et al. Максимизация эффективности липидных наночастиц siRNA для подавления активности печеночных генов in vivo. Angew Chem Int Edit. 2012;51(34):8529–33.
КАС
Статья
Google Scholar
Akinc A, Zumbuehl A, Goldberg M, Leshchiner ES, Busini V, Hossain N, et al.Комбинаторная библиотека липидоподобных материалов для доставки терапевтических средств РНКи. Нац биотехнолог. 2008;26(5):561–9.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Dong Y, Love KT, Dorkin JR, Sirirungruang S, Zhang Y, Chen D, et al. Липопептидные наночастицы для эффективной и селективной доставки миРНК грызунам и приматам. Proc Natl Acad Sci U S A. 2014;111(11):3955–60.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Maier MA, Jayaraman M, Matsuda S, Liu J, Barros S, Querbes W, et al. Биоразлагаемые липиды, позволяющие быстро устранять липидные наночастицы для системной доставки терапевтических средств РНК-интерференции. Мол Тер. 2013;21(8):1570–8.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Whitehead KA, Dorkin JR, Vegas AJ, Chang PH, Veiseh O, Matthews J, et al. Разлагаемые липидные наночастицы с предсказуемой активностью доставки миРНК in vivo.Нац коммун. 2014;5:4277.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Fenton OS, Kauffman KJ, Kaczmarek JC, McClellan RL, Jhunjhunwala S, Tibbitt MW, et al. Синтез и биологическая оценка ионизируемых липидных материалов для доставки РНК-мессенджера в В-лимфоциты in vivo. Adv Mater. 2017;29(33). https://doi.org/10.1002/adma.201606944.
Fenton OS, Kauffman KJ, McClellan RL, Appel EA, Dorkin JR, Tibbitt MW, et al.Биоинспирированные алкениламиноспирты Ионизируемые липидные материалы для высокоэффективной доставки мРНК in vivo. Adv Mater. 2016;28(15):2939–43.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Crommelin DJA, Anchordoquy TJ, Volkin DB, Jiskoot W, Mastrobattista E. Обращаясь к холодной реальности стабильности мРНК-вакцины. Дж. Фарм. 2021;110(3):997–1001.
КАС
пабмед
Статья
Google Scholar
Zhang X, Zhao W, Nguyen GN, Zhang C, Zeng C, Yan J и др. Функционализированные липидоподобные наночастицы для доставки мРНК in vivo и редактирования оснований. Научная реклама 2020;6(34):eabc2315.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Li B, Luo X, Deng B, Wang J, McComb DW, Shi Y, et al. Оптимизация ортогонального массива липидоподобных наночастиц для доставки мРНК in vivo. Нано Летт. 2015;15(12):8099–107.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Акита Х., Ишиба Р., Тогаси Р., Танге К., Накаи Ю., Хатакеяма Х. и др. Нейтральная липидная наночастица типа оболочки, состоящая из рН-активируемого и липидоподобного материала на основе витамина Е, в качестве платформы для переносчика гена, нацеленного на почечно-клеточную карциному. J Управление выпуском. 2015; 200:97–105.
КАС
пабмед
Статья
Google Scholar
Hou X, Zhang X, Zhao W, Zeng C, Deng B, McComb DW и др. Липидные наночастицы витаминов обеспечивают адоптивный перенос макрофагов для лечения бактериального сепсиса с множественной лекарственной устойчивостью. Нац Нанотехнолог. 2020;15(1):41–6.
Нац Нанотехнолог. 2020;15(1):41–6.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Конвей А., Мендель М., Ким К., Макговерн К., Бойко А., Чжан Л. и др. Невирусная доставка мРНК нуклеазы цинковых пальцев обеспечивает высокоэффективное редактирование генома in vivo нескольких терапевтических генов-мишеней.Мол Тер. 2019;27(4):866–77.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Sabnis S, Kumarasinghe ES, Salerno T, Mihai C, Ketova T, Senn JJ, et al. Новая серия аминолипидов для доставки мРНК: улучшенное эндосомальное ускользание и устойчивая фармакология и безопасность у приматов, отличных от человека. Мол Тер. 2018;26(6):1509–19.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Рамасвами С., Тонну Н., Тачикава К., Лимфонг П. , Вега Дж. Б., Кармали П. П. и др. Системная доставка матричной РНК фактора IX для заместительной белковой терапии. Proc Natl Acad Sci U S A. 2017;114(10):E1941–E50.
, Вега Дж. Б., Кармали П. П. и др. Системная доставка матричной РНК фактора IX для заместительной белковой терапии. Proc Natl Acad Sci U S A. 2017;114(10):E1941–E50.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Finn JD, Smith AR, Patel MC, Shaw L, Youniss MR, van Heteren J, et al. Однократное введение липидных наночастиц CRISPR/Cas9 обеспечивает надежное и стойкое редактирование генома in vivo.Cell Rep. 2018;22(9):2227–35.
КАС
пабмед
Статья
Google Scholar
Thevenot J, Troutier AL, David L, Delair T, Ladaviere C. Стерическая стабилизация липидно-полимерных частиц с помощью поли(этиленгликоль)-липидов. Биомакромолекулы. 2007;8(11):3651–60.
КАС
пабмед
Статья
Google Scholar
Witzigmann D, Kulkarni JA, Leung J, Chen S, Cullis PR, van der Meel R. Технология липидных наночастиц для терапевтической регуляции генов в печени. Adv Drug Deliv Rev. 2020; 159: 344–63.
Технология липидных наночастиц для терапевтической регуляции генов в печени. Adv Drug Deliv Rev. 2020; 159: 344–63.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Kauffman KJ, Dorkin JR, Yang JH, Heartlein MW, DeRosa F, Mir FF, et al. Оптимизация составов липидных наночастиц для доставки мРНК in vivo с дробным факторным и окончательным дизайном скрининга. Нано Летт. 2015;15(11):7300–6.
КАС
пабмед
Статья
Google Scholar
Dahlman JE, Kauffman KJ, Xing Y, Shaw TE, Mir FF, Dlott CC, et al. Наночастицы со штрих-кодом для высокопроизводительного открытия целевых терапевтических средств in vivo. Proc Natl Acad Sci U S A. 2017;114(8):2060–5.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Сато Ю., Хатакеяма Х., Сакураи Ю., Хёдо М., Акита Х. , Харашима Х.Чувствительный к рН катионный липид облегчает доставку липосомальной миРНК и активность подавления активности генов in vitro и in vivo. J Управление выпуском. 2012;163(3):267–76.
, Харашима Х.Чувствительный к рН катионный липид облегчает доставку липосомальной миРНК и активность подавления активности генов in vitro и in vivo. J Управление выпуском. 2012;163(3):267–76.
КАС
пабмед
Статья
Google Scholar
Gilleron J, Querbes W, Zeigerer A, Borodovsky A, Marsico G, Schubert U, et al. Анализ на основе изображений доставки siRNA, опосредованной липидными наночастицами, внутриклеточного транспорта и эндосомального побега. Нац биотехнолог.2013;31(7):638–46.
КАС
пабмед
Статья
Google Scholar
Сахай Г., Кербес В., Алаби С., Эльтухи А., Саркар С., Зуренко С. и др. Эффективность доставки siRNA липидными наночастицами ограничена эндоцитарной рециркуляцией. Нац биотехнолог. 2013;31(7):653–8.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Wittrup A, Ai A, Liu X, Hamar P, Trifonova R, Charisse K, et al.Визуализация липидного высвобождения siRNA из эндосом и нокдауна гена-мишени. Нац биотехнолог. 2015;33(8):870–6.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Sato Y, Kinami Y, Hashiba K, Harashima H. Различная кинетика поглощения липидных наночастиц печенью между аполипопротеином E / рецептором липопротеина низкой плотности и путем N-ацетил-d-галактозамина / асиалогликопротеинового рецептора.J Управление выпуском. 2020; 322: 217–26.
КАС
пабмед
Статья
Google Scholar
Wang F, Xiao W, Elbahnasawy MA, Bao X, Zheng Q, Gong L, et al. Оптимизация длины линкера конъюгатов манноза-холестерин для усиленной доставки мРНК в дендритные клетки с помощью липосом. Фронт Фармакол. 2018;9:980.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar

Эверс М.Дж., Кулкарни Дж.А., ван дер Меел Р., Каллис П.Р., Вейдер П., Шиффелерс Р.М. Современный дизайн и технологии быстрого смешивания липидных наночастиц для доставки нуклеиновых кислот. Малые методы. 2018;2(9):1700375.
Артикул
КАС
Google Scholar
Belliveau NM, Huft J, Lin PJ, Chen S, Leung AK, Leaver TJ, et al. Микрожидкостный синтез сильнодействующих липидных наночастиц предельного размера для доставки миРНК in vivo.Молекулярные нуклеиновые кислоты. 2012;1:e37.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar
Kaczmarek JC, Patel AK, Kauffman KJ, Fenton OS, Webber MJ, Heartlein MW, et al. Полимерно-липидные наночастицы для системной доставки мРНК в легкие. Angew Chem Int Ed Eng. 2016;55(44):13808–12.
КАС
Статья
Google Scholar
Патель А. К., Качмарек Дж.К., Бозе С., Кауфман К.Дж., Мир Ф., Хартлейн М.В. и соавт.Ингаляционные наноформулированные полиплексы мРНК для производства белка в эпителии легких. Adv Mater. 2019;31(8):e1805116.
К., Качмарек Дж.К., Бозе С., Кауфман К.Дж., Мир Ф., Хартлейн М.В. и соавт.Ингаляционные наноформулированные полиплексы мРНК для производства белка в эпителии легких. Adv Mater. 2019;31(8):e1805116.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar
Ковальски П.С., Капассо Пальмьеро У., Хуанг И., Рудра А., Лангер Р., Андерсон Д.Г. Ионизируемые аминополиэфиры, синтезированные посредством полимеризации третичных аминоспиртов с раскрытием кольца, для селективной доставки мРНК в ткани. Adv Mater. 2018;30(34):1801151.
Артикул
КАС
Google Scholar
Dahlman JE, Barnes C, Khan O, Thiriot A, Jhunjunwala S, Shaw TE, et al. Доставка эндотелиальной миРНК in vivo с использованием полимерных наночастиц с низкой молекулярной массой. Нац Нанотехнолог. 2014;9(8):648–55.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Хан О.Ф., Ковальски П.С., Долофф Дж.С., Цосие Дж.К., Бактаватчалу В., Винн С.Б. и др.Доставка эндотелиальной миРНК у нечеловеческих приматов с использованием ионизируемых низкомолекулярных полимерных наночастиц. Научная реклама 2018;4(6):eaar8409.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar
McCullough KC, Milona P, Thomann-Harwood L, Demoulins T, Englezou P, Suter R, et al. Доставка самоамплифицирующейся репликон-РНК-вакцины в дендритные клетки с помощью синтетических наночастиц. Вакцины (Базель). 2014;2(4):735–54.
Артикул
Google Scholar
Чахал Дж.С., Хан О.Ф., Купер К.Л., МакПартлан Дж.С., Цосие Дж.К., Тилли Л.Д. и др. Наночастицы дендример-РНК создают защитный иммунитет против летального вируса Эбола, гриппа h2N1 и токсоплазмы гондии при однократном введении. Proc Natl Acad Sci U S A. 2016;113(29):E4133–42.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Ислам М.А., Сюй Ю., Тао В., Убеллакер Дж.М., Лим М., Аум Д. и др. Восстановление подавления роста опухоли in vivo посредством системной доставки мРНК PTEN, опосредованной наночастицами. Нат Биомед Инж. 2018;2(11):850–64.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Kaczmarek JC, Kauffman KJ, Fenton OS, Sadtler K, Patel AK, Heartlein MW, et al. Оптимизация разлагаемых полимерно-липидных наночастиц для мощной системной доставки мРНК в эндотелий легких и иммунные клетки.Нано Летт. 2018;18(10):6449–54.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
McKinlay CJ, Benner NL, Haabeth OA, Waymouth RM, Wender PA. Улучшенная доставка мРНК в лимфоциты благодаря разнообразным липидным библиотекам высвобождаемых переносчиков, изменяющих заряд. Proc Natl Acad Sci U S A. 2018;115(26):E5859–E66.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar

Haabeth OAW, Blake TR, McKinlay CJ, Waymouth RM, Wender PA, Levy R. Вакцинация мРНК высвобождаемыми переносчиками, изменяющими заряд, вызывает реакции человеческих Т-клеток и излечивает установленные опухоли у мышей. Proc Natl Acad Sci U S A. 2018;115(39):E9153–E61.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
McKinlay CJ, Vargas JR, Blake TR, Hardy JW, Canada M, Contag CH, et al. Высвобождаемые транспортеры с изменением заряда (CART) для доставки и высвобождения мРНК у живых животных.Proc Natl Acad Sci U S A. 2017;114(4):E448–E56.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Scheel B, Teufel R, Probst J, Carralot JP, Geginat J, Radsak M, et al. Толл-подобная рецептор-зависимая активация нескольких типов клеток крови человека протамин-конденсированной мРНК. Евр Дж Иммунол. 2005;35(5):1557–66.
КАС
пабмед
Статья
Google Scholar

Себастьян М., Папахристофилу А., Вайс С., Фру М., Катомас Р., Хильбе В. и др. Исследование фазы Ib по оценке самоадъювантной мРНК-вакцины против рака (RNAactive (R)) в сочетании с местным облучением в качестве консолидирующего и поддерживающего лечения для пациентов с немелкоклеточным раком легкого IV стадии. БМК Рак. 2014;14:748.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar
Sebastian M, Schroder A, Scheel B, Hong HS, Muth A, von Boehmer L, et al.Фаза I / IIa исследования иммунотерапии рака на основе мРНК CV9201 у пациентов с немелкоклеточным раком легкого стадии IIIB / IV. Рак Иммунол Иммунотер. 2019;68(5):799–812.
КАС
пабмед
Статья
Google Scholar
Кублер Х., Шил Б., Гнад-Фогт У., Миллер К., Шульце-Земанн В., фон Дорп Ф. и др. Самоадъювантная вакцинация мРНК у пациентов с распространенным раком простаты: первое исследование фазы I/IIa на людях. J Иммунный рак.2015;3:26.
J Иммунный рак.2015;3:26.
ПабМед
ПабМед Центральный
Статья
Google Scholar
McCarthy HO, McCaffrey J, McCrudden CM, Zholobenko A, Ali AA, McBride JW, et al. Разработка и характеристика самособирающихся наночастиц с использованием биоинспирированного амфипатического пептида для доставки генов. J Управление выпуском. 2014; 189:141–9.
КАС
пабмед
Статья
Google Scholar
Udhayakumar VK, De Beuckelaer A, McCaffrey J, McCrudden CM, Kirschman JL, Vanover D, et al. Нанокомплексы мРНК на основе богатых аргинином пептидов эффективно стимулируют цитотоксический Т-клеточный иммунитет, зависящий от амфипатической организации пептида. Adv Healthc Mater. 2017;6(13). https://doi.org/10.1002/adhm.201601412.
Белл Г.Д., Ян И., Леунг Э., Криссансен Г.В. Трансфекция мРНК проникающим в клетку пептидом Xentry-protamine усиливается антагонистом TLR E6446. ПЛОС Один.2018;13(7):e0201464.
ПЛОС Один.2018;13(7):e0201464.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar
Zhang R, Tang L, Tian Y, Ji X, Hu Q, Zhou B, et al. Липосомы, модифицированные DP7-C, усиливают иммунный ответ и противоопухолевый эффект мРНК-вакцины на основе неоантигена. J Управление выпуском. 2020; 328: 210–21.
КАС
пабмед
Статья
Google Scholar
Татешита Н., Миура Н., Танака Х., Масуда Т., Оцуки С., Танге К. и др.Разработка носителя мРНК липоплексного типа, состоящего из ионизируемого липида с каркасом витамина Е и пептида KALA, для использования в качестве противораковой вакцины ex vivo на основе дендритных клеток. J Управление выпуском. 2019;310:36–46.
КАС
пабмед
Статья
Google Scholar
Lou B, De Koker S, Lau CYJ, Hennink WE, Mastrobattista E. Полиплексы мРНК с постконъюгированными пептидами GALA эффективно нацелены, трансфицируют и активируют антигенпрезентирующие клетки. Биоконьюг Хим. 2019;30(2):461–75.
Биоконьюг Хим. 2019;30(2):461–75.
КАС
пабмед
Статья
Google Scholar
Brito LA, Chan M, Shaw CA, Hekele A, Carsillo T, Schaefer M, et al. Катионная наноэмульсия для доставки РНК-вакцин нового поколения. Мол Тер. 2014;22(12):2118–29.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Liu L, Wang Y, Miao L, Liu Q, Musetti S, Li J, et al.Комбинированная иммунотерапия нановакциной мРНК MUC1 и блокадой CTLA-4 эффективно ингибирует рост трижды негативного рака молочной железы. Мол Тер. 2018;26(1):45–55.
КАС
пабмед
Статья
Google Scholar
Le Moignic A, Malard V, Benvegnu T, Lemiegre L, Berchel M, Jaffres PA, et al. Доклиническая оценка триманнозилированных липополиплексов мРНК в качестве терапевтических противораковых вакцин, нацеленных на дендритные клетки. J Управление выпуском.2018; 278:110–21.
J Управление выпуском.2018; 278:110–21.
ПабМед
Статья
КАС
Google Scholar
Персано С., Гевара М.Л., Ли З., Май Дж., Феррари М., Помпа П.П. и др. Липополиплекс потенцирует противоопухолевый иммунитет при вакцинации на основе мРНК. Биоматериалы. 2017;125:81–9.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Ols S, Yang L, Thompson EA, Pushparaj P, Tran K, Liang F, et al.Путь введения вакцины изменяет перенос антигена, но не врожденный или адаптивный иммунитет. Cell Rep. 2020;30(12):3964–71 e7.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Парди Н., Туишиме С., Мурамацу Х., Карико К., Муи Б.Л., Там Ю.К. и др. Кинетика экспрессии мРНК, модифицированной нуклеозидами, доставляемой в липидных наночастицах мышам различными путями. J Управление выпуском. 2015; 217:345–51.
J Управление выпуском. 2015; 217:345–51.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Li M, Zhao M, Fu Y, Li Y, Gong T, Zhang Z, et al. Улучшенная интраназальная доставка мРНК-вакцины за счет преодоления назального эпителиального барьера внутри- и парацеллюлярным путями. J Управление выпуском. 2016; 228:9–19.
КАС
пабмед
Статья
Google Scholar
Guo Y, Lei K, Tang L. Доставка неоантигенной вакцины для персонализированной противоопухолевой иммунотерапии. Фронт Иммунол. 2018;9:1499.
ПабМед
ПабМед Центральный
Статья
КАС
Google Scholar
Ван Линт С., Ренманс Д., Броос К., Геталс Л., Менхаут С., Бентейн Д. и другие. Внутриопухолевая доставка мРНК TriMix приводит к активации Т-клеток путем перекрестного представления дендритных клеток. Рак Иммунол Рез. 2016;4(2):146–56.
2016;4(2):146–56.
ПабМед
Статья
КАС
Google Scholar
Jansen Y, Kruse V, Corthals J, Schats K, van Dam PJ, Seremet T, et al. Рандомизированное контролируемое клиническое исследование фазы II аутологичных дендритных клеток, полученных из моноцитов, с электропорацией мРНК (TriMixDC-MEL) в качестве адъювантного лечения пациентов с меланомой стадии III/IV, у которых нет признаков заболевания после резекции макрометастазов.Рак Иммунол Иммунотер. 2020;69(12):2589–98.
ПабМед
Статья
КАС
Google Scholar
Wilgenhof S, Van Nuffel AMT, Benteyn D, Corthals J, Aerts C, Heirman C, et al. Фаза IB исследования внутривенной синтетической мРНК-электропорации иммунотерапии дендритных клеток у предварительно леченных пациентов с прогрессирующей меланомой. Энн Онкол. 2013;24(10):2686–93.
КАС
пабмед
Статья
Google Scholar

Huo M, Zhao Y, Satterlee AB, Wang Y, Xu Y, Huang L. Нацеленная на опухоль доставка основания сунитиниба усиливает вакцинную терапию прогрессирующей меланомы за счет ремоделирования микроокружения опухоли. J Управление выпуском. 2017; 245:81–94.
КАС
пабмед
Статья
Google Scholar
Граббе С., Хаас Х., Дикен М., Кранц Л.М., Ланггут П., Сахин У. Преобразование персонализированных противораковых вакцин с наночастицами в клиническое применение: тематическое исследование с РНК-липоплексами для лечения меланомы.Наномедицина (Лондон). 2016;11(20):2723–34.
КАС
Статья
Google Scholar
Ши Ю. Клинический перевод наномедицины и биоматериалов для иммунотерапии рака: прогресс и перспективы. Adv Ther. 2020;3(9):9.
Google Scholar
Батич К.А., Митчелл Д.А., Хили П., Херндон Дж. Э. 2-й, Сэмпсон Дж.Х. Один, два, три раза вывод: воспроизводимость испытаний вакцины на дендритных клетках, направленных против цитомегаловируса при глиобластоме.Клин Рак Рез. 2020;26(20):5297–303.
Э. 2-й, Сэмпсон Дж.Х. Один, два, три раза вывод: воспроизводимость испытаний вакцины на дендритных клетках, направленных против цитомегаловируса при глиобластоме.Клин Рак Рез. 2020;26(20):5297–303.
КАС
пабмед
Статья
Google Scholar
Chen X, Yang J, Wang L, Liu B. Персонализированная неоантигенная вакцинация синтетическими длинными пептидами: последние достижения и перспективы на будущее. Тераностика. 2020;10(13):6011–23.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Cafri G, Gartner JJ, Zaks T, Hopson K, Levin N, Paria BC, et al.мРНК-вакцино-индуцированный неоантиген-специфический Т-клеточный иммунитет у пациентов с раком желудочно-кишечного тракта. Джей Клин Инвест. 2020;130(11):5976–88.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Ott PA, Hu ZT, Keskin DB, Shukla SA, Sun J, Bozym DJ, et al. Иммуногенная персональная неоантигенная вакцина для больных меланомой. Природа. 2017;547(7662):217-+.
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Сахин У., Дерхованесян Э., Миллер М., Клоке Б.П., Саймон П., Лоуэр М. и др. Персонализированные РНК-мутаномные вакцины мобилизуют полиспецифический терапевтический иммунитет против рака. Природа. 2017;547(7662):222–6.
КАС
пабмед
Статья
Google Scholar
Беррис Х.А. Многоцентровое исследование фазы I для оценки безопасности, переносимости и иммуногенности мРНК-4157 отдельно у пациентов с резецированными солидными опухолями и в комбинации с пембролизумабом у пациентов с неоперабельными солидными опухолями; 2019.
Книга
Google Scholar
Опосредованное светом открытие наноразмерной организации поверхности и сетей взаимодействия межклеточных рецепторов
Введение
Клеточная функция регулируется обменом информацией с внешним миром. Протеотип клеточной поверхности (поверхностный) является сигнальным шлюзом к клетке, который обеспечивает и ограничивает коммуникацию 1 . Сборки наноразмерных рецепторов соединяют внешние стимулы с путями внутриклеточного ответа внутри поверхностных сигнальных доменов 2 .Эти субструктуры играют решающую роль в действии лекарств, взаимодействии хозяин-патоген и межклеточной коммуникации 3 . Таким образом, картирование поверхностных ландшафтов, нацеленных на лиганды, с наноразмерной точностью должно дать фундаментальное представление о функции клеточной передачи сигналов в норме и болезни с широкими последствиями для разработки терапевтических стратегий. В частности, распутывание сигнальных взаимодействий поверхностиома в пространстве и времени значительно продвинет разработку лекарств (деконволюция мишеней лекарств и механизма действия) и сделает возможными открытия в широких областях исследований, включая клеточную биологию (рассечение сигналов рецепторов) 4 , микробиологию (выявление участков связывания патогенов).
Протеотип клеточной поверхности (поверхностный) является сигнальным шлюзом к клетке, который обеспечивает и ограничивает коммуникацию 1 . Сборки наноразмерных рецепторов соединяют внешние стимулы с путями внутриклеточного ответа внутри поверхностных сигнальных доменов 2 .Эти субструктуры играют решающую роль в действии лекарств, взаимодействии хозяин-патоген и межклеточной коммуникации 3 . Таким образом, картирование поверхностных ландшафтов, нацеленных на лиганды, с наноразмерной точностью должно дать фундаментальное представление о функции клеточной передачи сигналов в норме и болезни с широкими последствиями для разработки терапевтических стратегий. В частности, распутывание сигнальных взаимодействий поверхностиома в пространстве и времени значительно продвинет разработку лекарств (деконволюция мишеней лекарств и механизма действия) и сделает возможными открытия в широких областях исследований, включая клеточную биологию (рассечение сигналов рецепторов) 4 , микробиологию (выявление участков связывания патогенов). 5 и иммунология (выяснение иммуносинаптической связи) 6 .
5 и иммунология (выяснение иммуносинаптической связи) 6 .
По сей день сигнальный ландшафт поверхности остается terra incognita , который не может быть выведен из клеточных протеомов и транскриптомов. Были установлены специальные стратегии для профилирования клеточных поверхностемов 7 . Например, широкое применение технологии захвата клеточной поверхности (CSC) 8,9 установило N-гликозилированные поверхности для многочисленных типов клеток (совместно сообщается в Атласе белков клеточной поверхности, CSPA https://wlab.ethz.ch/cspa/) 10 и позволили in silico охарактеризовать всю поверхность тела человека 11 . В то время как репортерные анализы на основе флуоресценции 12–14 и платформы генетически модифицированных рецепторов 15–17 установили ассоциации между рецепторами путем высокопроизводительного тестирования бинарных взаимодействий внутри и между поверхностями, технология Ligand-Receptor Capture (LRC) выявила прямые лигандные взаимодействия N-гликозилированных рецепторов 18,19 . Недавно основанные на ферментах стратегии маркировки близости, такие как APEX 20 , HRP или PUP-iT 21 , позволили маркировать и на основе МС идентифицировать липидные рафты 22 , сигнальные домены фактора роста 23 , окрестности рецепторов В-клеток 24 и интактные синапсы нейронов 25 на живых клетках и в фиксированных тканях 26 . Таким образом, основанная на фотокатализаторах стратегия маркировки близостью продемонстрировала улучшенную пространственную точность 27 . Однако в таких методах меченые зонды одновременно назначаются всем поверхностно-доступным рецепторам с помощью антител или генетического слияния, что приводит к отсутствию гибкости и специфичности для обнаружения лиганд-мишеней сигнальных доменов поверхности, оставляя функциональные горячие точки клеточного поверхностного ландшафта неизученными. .
Недавно основанные на ферментах стратегии маркировки близости, такие как APEX 20 , HRP или PUP-iT 21 , позволили маркировать и на основе МС идентифицировать липидные рафты 22 , сигнальные домены фактора роста 23 , окрестности рецепторов В-клеток 24 и интактные синапсы нейронов 25 на живых клетках и в фиксированных тканях 26 . Таким образом, основанная на фотокатализаторах стратегия маркировки близостью продемонстрировала улучшенную пространственную точность 27 . Однако в таких методах меченые зонды одновременно назначаются всем поверхностно-доступным рецепторам с помощью антител или генетического слияния, что приводит к отсутствию гибкости и специфичности для обнаружения лиганд-мишеней сигнальных доменов поверхности, оставляя функциональные горячие точки клеточного поверхностного ландшафта неизученными. .
Генераторы синглетного кислорода (SOG), как известно, проявляют способность маркировать близость посредством активируемого светом образования активных форм синглетного кислорода, которые окисляют биомолекулы в нанометровом диапазоне 28,29 . Малая молекула SOG 30 и генетически кодируемый SOG широко используются в науках о жизни для фотодинамической инактивации белков 31–34 и клеток 35,36 , для корреляционной светоэлектронной микроскопии 37 и для обнаружения внутриклеточного белка -белковые взаимодействия 38,39 в различных организмах.Способность SOG декодировать взаимодействия клеточной поверхности недавно была продемонстрирована путем идентификации мишени связывания флуоресцентного пептида, выделяющего нервы, в тканях млекопитающих 40 . Благодаря небольшому размеру и возможностям мечения in situ под контролем света, SOG представляют собой идеальные зонды для мечения, управляемого лигандом, и обнаружения сигнальных доменов поверхности.
Малая молекула SOG 30 и генетически кодируемый SOG широко используются в науках о жизни для фотодинамической инактивации белков 31–34 и клеток 35,36 , для корреляционной светоэлектронной микроскопии 37 и для обнаружения внутриклеточного белка -белковые взаимодействия 38,39 в различных организмах.Способность SOG декодировать взаимодействия клеточной поверхности недавно была продемонстрирована путем идентификации мишени связывания флуоресцентного пептида, выделяющего нервы, в тканях млекопитающих 40 . Благодаря небольшому размеру и возможностям мечения in situ под контролем света, SOG представляют собой идеальные зонды для мечения, управляемого лигандом, и обнаружения сигнальных доменов поверхности.
Здесь мы разработали и применили управляемую светом технологию обнаружения приближения, называемую LUX-MS, для систематического изучения ландшафта передачи сигналов поверхности на живых клетках.В этом подходе используется малая молекула SOG, связанная с лигандом, для специфического и легко настраиваемого биотинилирования лиганд-проксимальных белков, что позволяет идентифицировать их с использованием высокопроизводительного роботизированного рабочего процесса прототипирования. Мы демонстрируем пространственную специфичность и широкую применимость нашего подхода к декодированию поверхностных сигнальных доменов малых молекул, биомолекул и интактных патогенов в системах млекопитающих и бактерий на основе открытий и без необходимости генетических манипуляций.Наконец, мы продемонстрировали полезность LUX-MS для выяснения сложных межклеточных сигнальных доменов поверхности, картируя молекулярную архитектуру функциональных иммуносинапсов в беспрецедентных деталях.
Мы демонстрируем пространственную специфичность и широкую применимость нашего подхода к декодированию поверхностных сигнальных доменов малых молекул, биомолекул и интактных патогенов в системах млекопитающих и бактерий на основе открытий и без необходимости генетических манипуляций.Наконец, мы продемонстрировали полезность LUX-MS для выяснения сложных межклеточных сигнальных доменов поверхности, картируя молекулярную архитектуру функциональных иммуносинапсов в беспрецедентных деталях.
Результаты
Разработка технологии LUX-MS
Сначала мы протестировали способность низкомолекулярного тиородамина SOG фотоокислять трансферриновые белки in vitro . Количественный МС-анализ и открытый поиск модификаций выявили светозависимое превращение аминокислот гистидина, цистеина, триптофана и метионина в ранее описанные продукты фотоокисления 28 (, дополнительная рис.1 ). Среди прочего, наиболее часто образующаяся модификация 2-оксогистидина (His+14 Да) несет светоиндуцированную кетоновую группу, которая обеспечивает функционализацию посредством образования гидразона с использованием гидразидсодержащих линкеров. Действительно, инкубация фотоокисленного трансферрина с биотин-гидразидным (BH) линкером привела к значительному потреблению 2-оксо-гистидина (, дополнительная рис. 2, ), что указывает на биотинилирование светомодифицированных белков. Затем мы объединили контролируемое светом биотинилирование белка с роботизированной обработкой с жидкостной хроматографией и тандемной масс-спектрометрией с высокоэффективной жидкостной хроматографией (ЖХ-МС/МС) для создания оптопротеомной технологии LUX-MS (, рис.1 ). SOG-проксимальные белки, таким образом, биотинилируются в живых клетках в физиологических условиях светом и захватываются после лизиса клеток с помощью наконечников, функционализированных стрептавидином. Расщепление на гранулах с использованием протеолитических ферментов высвобождает пептиды на N- и C-концах исходного сайта мечения, которые впоследствии без метки количественно определяют с помощью масс-спектрометрии. SOG-проксимальные белки в конечном итоге идентифицируются без гипотез путем светозависимого обогащения по сравнению с неосвещенным контролем.
Действительно, инкубация фотоокисленного трансферрина с биотин-гидразидным (BH) линкером привела к значительному потреблению 2-оксо-гистидина (, дополнительная рис. 2, ), что указывает на биотинилирование светомодифицированных белков. Затем мы объединили контролируемое светом биотинилирование белка с роботизированной обработкой с жидкостной хроматографией и тандемной масс-спектрометрией с высокоэффективной жидкостной хроматографией (ЖХ-МС/МС) для создания оптопротеомной технологии LUX-MS (, рис.1 ). SOG-проксимальные белки, таким образом, биотинилируются в живых клетках в физиологических условиях светом и захватываются после лизиса клеток с помощью наконечников, функционализированных стрептавидином. Расщепление на гранулах с использованием протеолитических ферментов высвобождает пептиды на N- и C-концах исходного сайта мечения, которые впоследствии без метки количественно определяют с помощью масс-спектрометрии. SOG-проксимальные белки в конечном итоге идентифицируются без гипотез путем светозависимого обогащения по сравнению с неосвещенным контролем.
Рис. 1 Стратегия LUX-MS для активируемой светом идентификации сигнальных доменов поверхности.
Освещение генераторов синглетного кислорода (SOG), связанных с антителами или лигандами, способствует пространственно ограниченному окислению целевых поверхностных ландшафтов с пространственно-временной специфичностью. Таким образом, регулируемая светом активация аминокислот, таких как гистидин (His), позволяет химическую маркировку белков в физиологических условиях с использованием гидразидных линкеров, функционализированных биотином. Меченые белки автоматически захватываются и обрабатываются с помощью роботизированной платформы, работающей с жидкостями, что обеспечивает быструю и воспроизводимую количественную оценку на основе масс-спектрометрии для большого количества образцов.Относительная количественная оценка между освещенными и неосвещенными образцами в конечном итоге позволяет обнаруживать в масштабах всего протеома острые сигнальные домены поверхности, нацеленные на антитела и лиганды, на живых клетках.
LUX-MS позволяет картировать мишени связывания антител в масштабах всего протеома и наноразмерную организацию поверхностных молекул
Взаимодействия поверхностных молекул определяют специфичность и сигнальную способность антител, и их выяснение имеет решающее значение для разработки терапевтических антител. В контексте очень успешного терапевтического антитела ритуксимаба 41 десятилетия интенсивных исследований выявили организацию клеточной поверхности его первичного рецептора CD20 на клетках-мишенях В-лимфомы человека.Чтобы оценить LUX-MS для получения дополнительной информации о действии ритуксимаба, мы соединили антитела против CD20 с низкомолекулярным SOG тиородамином (, рис. 2a, ) и инкубировали полученные конструкции Ab-SOG с мононуклеарными клетками периферической крови человека (PBMC), которые были изолированы от здоровых доноров. Проточный цитометрический анализ выявил связывание антител с 8% всех обнаруженных клеток (, рис. 2b ), что соответствует типичной фракции CD20-положительных клеток в РВМС человека (5-10%) 42 .Последующее освещение в течение 5 минут в присутствии BH с последующей проточной цитометрией продемонстрировало значительное биотинилирование клеточной поверхности большинства (96 %) Ab-SOG, связанных, но не несвязанных клеток, что указывает на эффективную близкую маркировку живых клеток в сложных биологических средах с пространственной специфичностью. Затем мы исследовали динамику LUX-мечения с использованием Ab-SOG на клеточной линии В-лимфомы человека и наблюдали, что зависящее от света биотинилирование клеточной поверхности можно точно настроить в зависимости от продолжительности освещения (рис.2c и Дополнительный рис. 3 ). Известно, что буферные условия модулируют производство и время жизни синглетных форм кислорода 43 . Действительно, мы достигли 5-кратного повышения эффективности фотоокисления путем замены H 2 O на D 2 O в буферной системе и двукратного увеличения биотинилирования клеточной поверхности при использовании 2-(аминометил)имидазола в качестве катализатора для повышения образование гидразона вместо ранее применявшейся 5-метоксиантраниловой кислоты 19,43,44 ( Дополнительный рис.
2b ), что соответствует типичной фракции CD20-положительных клеток в РВМС человека (5-10%) 42 .Последующее освещение в течение 5 минут в присутствии BH с последующей проточной цитометрией продемонстрировало значительное биотинилирование клеточной поверхности большинства (96 %) Ab-SOG, связанных, но не несвязанных клеток, что указывает на эффективную близкую маркировку живых клеток в сложных биологических средах с пространственной специфичностью. Затем мы исследовали динамику LUX-мечения с использованием Ab-SOG на клеточной линии В-лимфомы человека и наблюдали, что зависящее от света биотинилирование клеточной поверхности можно точно настроить в зависимости от продолжительности освещения (рис.2c и Дополнительный рис. 3 ). Известно, что буферные условия модулируют производство и время жизни синглетных форм кислорода 43 . Действительно, мы достигли 5-кратного повышения эффективности фотоокисления путем замены H 2 O на D 2 O в буферной системе и двукратного увеличения биотинилирования клеточной поверхности при использовании 2-(аминометил)имидазола в качестве катализатора для повышения образование гидразона вместо ранее применявшейся 5-метоксиантраниловой кислоты 19,43,44 ( Дополнительный рис. 4 ). Затем мы провели эксперимент LUX-MS, нацеленный на CD20, обработав культивированные клетки B-лимфомы человека Ab-SOG перед освещением в течение 0-5 минут. Всего с помощью LUX-MS было количественно определено 1674 белка, по крайней мере, с двумя пептидами на белок (, дополнительная таблица 1, ). В то время как большинство белков были одинаково распространены в условиях без освещения, 63 белка, включая 55 (87%) поверхностных белков, показали поразительное обогащение, которое коррелировало с продолжительностью освещения и достигло кульминации в 5-кратном увеличении содержания после 5 минут освещения (). Инжир.2d и Дополнительный рис. 5 ). Таким образом, идентификация LUX-MS была ограничена 43 из 215 членов поверхности, о которых сообщалось CSPA, и включала негликозилированные поверхностные белки, которые обычно избегают опросов поверхностейома на основе CSC или LRC (, дополнительная рис. 6, ). Таким образом, обнаружение близости с помощью LUX-MS позволяет проводить пространственное прототипирование с субповерхностным разрешением.
4 ). Затем мы провели эксперимент LUX-MS, нацеленный на CD20, обработав культивированные клетки B-лимфомы человека Ab-SOG перед освещением в течение 0-5 минут. Всего с помощью LUX-MS было количественно определено 1674 белка, по крайней мере, с двумя пептидами на белок (, дополнительная таблица 1, ). В то время как большинство белков были одинаково распространены в условиях без освещения, 63 белка, включая 55 (87%) поверхностных белков, показали поразительное обогащение, которое коррелировало с продолжительностью освещения и достигло кульминации в 5-кратном увеличении содержания после 5 минут освещения (). Инжир.2d и Дополнительный рис. 5 ). Таким образом, идентификация LUX-MS была ограничена 43 из 215 членов поверхности, о которых сообщалось CSPA, и включала негликозилированные поверхностные белки, которые обычно избегают опросов поверхностейома на основе CSC или LRC (, дополнительная рис. 6, ). Таким образом, обнаружение близости с помощью LUX-MS позволяет проводить пространственное прототипирование с субповерхностным разрешением. Во втором эксперименте с использованием повторов мы использовали статистическое тестирование для выявления достоверных SOG-проксимальных кандидатов (кратность изменения обогащения> 1.5 и значение p <0,05, Рис. 2e, Дополнительная таблица 1 и Дополнительные данные 1 ). Среди лучших совпадений мы обнаружили изоформы иммуноглобулина G2 (IgG2) Ab-SOG, основную мишень связывания CD20, человеческий лейкоцитарный антиген класса II, тетраспанин CD37, CD298 и CD71, которые, как было показано, физически или функционально связаны с
Во втором эксперименте с использованием повторов мы использовали статистическое тестирование для выявления достоверных SOG-проксимальных кандидатов (кратность изменения обогащения> 1.5 и значение p <0,05, Рис. 2e, Дополнительная таблица 1 и Дополнительные данные 1 ). Среди лучших совпадений мы обнаружили изоформы иммуноглобулина G2 (IgG2) Ab-SOG, основную мишень связывания CD20, человеческий лейкоцитарный антиген класса II, тетраспанин CD37, CD298 и CD71, которые, как было показано, физически или функционально связаны с
Рис. 2 Открытие протеомных мишеней для связывания антител и наноразмерной организации поверхности.
a Схема декорирования антитела (Ab) генератором синглетного кислорода (SOG) тиородамином. b , Проточный цитометрический анализ мононуклеарных клеток периферической крови человека (PBMC), окрашенных на биотин клеточной поверхности и иммуноглобулин, до и после LUX-мечения анти-CD20 Ab-SOG. c , График гистограммы, показывающий светозависимое биотинилирование клеточной поверхности LUX-меченых клеток B-лимфомы SUDHL6. d , Диаграмма рассеяния, показывающая зависящее от света количество белков, определенных с помощью LUX-MS, из клеток B-лимфомы SUDHL6, обработанных Ab-SOG. Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно.Серый цвет указывает на 5-кратное количественное обогащение. e , слева, график Volcano, показывающий относительное количество белков, определенных с помощью LUX-MS, в клетках B-лимфомы SUDHL6, обработанных анти-CD20 Ab-SOG, с освещением и без него в течение 5 мин. Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Красные и серые точки представляют известные и ранее неизвестные белки, ассоциированные с CD20, соответственно. Выделены первичная мишень CD20 и цепи используемого антитела.Справа, литература и сеть взаимодействия CD20 с клеточной поверхностью на основе LUX-MS. f , PCA-анализ участков поверхности CD38, CD54, CD166 и CD220 на живых клетках B-лимфомы SUDHL6, идентифицированных с помощью LUX-MS под контролем антител.
d , Диаграмма рассеяния, показывающая зависящее от света количество белков, определенных с помощью LUX-MS, из клеток B-лимфомы SUDHL6, обработанных Ab-SOG. Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно.Серый цвет указывает на 5-кратное количественное обогащение. e , слева, график Volcano, показывающий относительное количество белков, определенных с помощью LUX-MS, в клетках B-лимфомы SUDHL6, обработанных анти-CD20 Ab-SOG, с освещением и без него в течение 5 мин. Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Красные и серые точки представляют известные и ранее неизвестные белки, ассоциированные с CD20, соответственно. Выделены первичная мишень CD20 и цепи используемого антитела.Справа, литература и сеть взаимодействия CD20 с клеточной поверхностью на основе LUX-MS. f , PCA-анализ участков поверхности CD38, CD54, CD166 и CD220 на живых клетках B-лимфомы SUDHL6, идентифицированных с помощью LUX-MS под контролем антител. Список MS-количественных белков ( дополнительная таблица 1 ) и полностью интерактивный график вулкана ( дополнительные данные 1 ) предоставляются для облегчения исследования данных.
Список MS-количественных белков ( дополнительная таблица 1 ) и полностью интерактивный график вулкана ( дополнительные данные 1 ) предоставляются для облегчения исследования данных.
CD20 на поверхности клеток В-лимфомы 45–53 . Кроме того, LUX-MS выявил близость CD20 к комплексу IgM В-клеточного рецептора (BCR) (CD79A, CD79B, тяжелой и легкой цепи IgM), включая BCR-ассоциированные CD19, CD38 и ITGA4, подтверждая модель, в которой связывание ритуксимаба с CD20 способно эрадикировать Клетки B-лимфомы путем изменения передачи сигналов BCR, способствующих выживанию 54,55 .Таким образом, выполнение высокопроизводительной LUX-MS на той же клеточной линии B-лимфомы с использованием различных SOG-связанных антител (против CD38, CD54, CD166 и CD220) выявило специфические для рецептора окрестности поверхности, которые охватывают различные функции белков ( Fig. 2f и , дополнительная таблица 1 ). В совокупности пространственно-временная специфичность обнаружения близости LUX-MS позволяет выявить взаимодействия антитело-поверхностный комплекс в масштабах всего протеома, а также обеспечивает пропускную способность, необходимую для систематического картирования наноразмерной организации поверхностного слоя.
В совокупности пространственно-временная специфичность обнаружения близости LUX-MS позволяет выявить взаимодействия антитело-поверхностный комплекс в масштабах всего протеома, а также обеспечивает пропускную способность, необходимую для систематического картирования наноразмерной организации поверхностного слоя.
LUX-MS расшифровывает поверхностные сигнальные домены низкомолекулярных препаратов и биомолекул биспецифические антитела (DBA)
56 и технологии внеклеточных лекарственных конъюгатов (EDC) 47 . Таким образом, EDC использует близость клеточной поверхности между антителами-мишенями, имеющими отношение к заболеванию, и низкомолекулярными рецепторами полезной нагрузки для эффективной абляции клеток.Тем не менее, идентичность низкомолекулярных рецепторов и их организация в поверхностном слое живых клеток остаются ограниченными, поскольку основанные на ферментах стратегии обнаружения близости не приспособлены к использованию низкомолекулярных лигандов. Чтобы проверить, может ли LUX-MS выявлять низкомолекулярные поверхностные домены, тиородамин SOG был ковалентно конъюгирован с CG1, низкомолекулярным лекарственным средством, которое избирательно ингибирует активную субъединицу перекачки ионов (ATP1A1) повсеместно экспрессируемого человеческого Na + / K + -АТФазный комплекс (NKA) ( рис.3а ) 47 . Культивированные промиелоцитарные лейкозные клетки инкубировали с увеличивающимися концентрациями конъюгата и выполняли LUX-мечение, как описано выше. Анализ FACS показал индуцированное светом и зависящее от концентрации биотинилирование клеточной поверхности, которое было насыщено при 5 мкМ конъюгата, демонстрируя связывание и мечение поверхностных молекул (, рис. 3b, ). Затем меченые клетки подвергли рабочему процессу LUX-MS, что привело к значительному обогащению 48 белками, локализованными на поверхности, при освещении (рис.3c, дополнительная таблица 2 и , дополнительные данные 2 ), включая субъединицу бета3 NKA (ATP1B3), что указывает на то, что LUX-MS позволяет обнаруживать взаимодействия малых молекул с рецепторами на основе открытий.
Чтобы проверить, может ли LUX-MS выявлять низкомолекулярные поверхностные домены, тиородамин SOG был ковалентно конъюгирован с CG1, низкомолекулярным лекарственным средством, которое избирательно ингибирует активную субъединицу перекачки ионов (ATP1A1) повсеместно экспрессируемого человеческого Na + / K + -АТФазный комплекс (NKA) ( рис.3а ) 47 . Культивированные промиелоцитарные лейкозные клетки инкубировали с увеличивающимися концентрациями конъюгата и выполняли LUX-мечение, как описано выше. Анализ FACS показал индуцированное светом и зависящее от концентрации биотинилирование клеточной поверхности, которое было насыщено при 5 мкМ конъюгата, демонстрируя связывание и мечение поверхностных молекул (, рис. 3b, ). Затем меченые клетки подвергли рабочему процессу LUX-MS, что привело к значительному обогащению 48 белками, локализованными на поверхности, при освещении (рис.3c, дополнительная таблица 2 и , дополнительные данные 2 ), включая субъединицу бета3 NKA (ATP1B3), что указывает на то, что LUX-MS позволяет обнаруживать взаимодействия малых молекул с рецепторами на основе открытий. Кроме того, мы обнаружили рецептор клеточной поверхности Basigin (CD147), который является функциональной мишенью конъюгатов внеклеточных лекарств, нагруженных CG1, на типе клеток, использованном в этом эксперименте 47 . Следовательно, данные свидетельствуют о том, что рецепторы, нацеленные на малые молекулы, и связанные с ними сигнальные домены поверхности могут быть обнаружены с помощью LUX-MS, что в конечном итоге позволит обнаружить мишени для разработки DBA или EDC.
Кроме того, мы обнаружили рецептор клеточной поверхности Basigin (CD147), который является функциональной мишенью конъюгатов внеклеточных лекарств, нагруженных CG1, на типе клеток, использованном в этом эксперименте 47 . Следовательно, данные свидетельствуют о том, что рецепторы, нацеленные на малые молекулы, и связанные с ними сигнальные домены поверхности могут быть обнаружены с помощью LUX-MS, что в конечном итоге позволит обнаружить мишени для разработки DBA или EDC.
Рис. 3 Расшифровка поверхностных сигнальных доменов низкомолекулярных препаратов и биомолекул.
a Схема соединения низкомолекулярных препаратов сердечного гликозида CG1 с генератором синглетного кислорода (SOG) тиородамином. b , График гистограммы, показывающий светозависимое биотинилирование клеточной поверхности LUX-меченых клеток промиелоцитарного лейкоза HL60. c , График вулкана, показывающий относительное количество белков, определенных с помощью LUX-MS, из клеток промиелоцитарного лейкоза HL60, обработанных CG1-SOG, с освещением и без него в течение 5 мин. Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Красные, зеленые и синие точки представляют собой мишень связывания CG1, известных и ранее неизвестных взаимодействующих поверхностей, соответственно. Первые два выделены. d , Схема соединения биомолекул инсулина и трансферрина с генератором синглетного кислорода (SOG) тиородамином. e , График вулкана, показывающий относительное количество белков, определенных с помощью LUX-MS, из обработанных инсулином-SOG и трансферрином-SOG клеток B-лимфомы SUDHL6, освещенных в течение 5 минут.Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Коричневые, красные и синие точки обозначают лиганд, первичную мишень связывания и потенциальные взаимодействующие поверхности, соответственно. Первые два выделены вместе с известными взаимодействующими поверхностями. f , Схематическое изображение сети взаимодействия поверхности, идентифицированной с помощью LUX-MS.
Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Красные, зеленые и синие точки представляют собой мишень связывания CG1, известных и ранее неизвестных взаимодействующих поверхностей, соответственно. Первые два выделены. d , Схема соединения биомолекул инсулина и трансферрина с генератором синглетного кислорода (SOG) тиородамином. e , График вулкана, показывающий относительное количество белков, определенных с помощью LUX-MS, из обработанных инсулином-SOG и трансферрином-SOG клеток B-лимфомы SUDHL6, освещенных в течение 5 минут.Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Коричневые, красные и синие точки обозначают лиганд, первичную мишень связывания и потенциальные взаимодействующие поверхности, соответственно. Первые два выделены вместе с известными взаимодействующими поверхностями. f , Схематическое изображение сети взаимодействия поверхности, идентифицированной с помощью LUX-MS. Список MS-количественных белков ( дополнительная таблица 2 ) и полностью интерактивные графики вулканов ( дополнительные данные 2 и 3 ) предоставляются для облегчения исследования данных.
Список MS-количественных белков ( дополнительная таблица 2 ) и полностью интерактивные графики вулканов ( дополнительные данные 2 и 3 ) предоставляются для облегчения исследования данных.
Затем мы спросили, можно ли применять LUX-MS для выявления поверхностных взаимодействий более сложных лигандов, то есть секретируемых биомолекул, которые являются ключевыми элементами межклеточной коммуникации. Поэтому мы соединили тиородамин SOG с пептидным гормоном инсулином и гликопротеином трансферрином плазмы, которые регулируют клеточный гомеостаз посредством взаимодействия с гомо- или гетеродимерами рецептора инсулина (INR) или рецептора инсулиноподобного фактора роста 1 (IGF1R) и рецептора трансферрина (TFRC) , соответственно ( Рис.3д ). LUX-MS проводили на клетках В-лимфомы человека, которые обрабатывали любым конъюгатом и освещали в течение 5 мин. Результаты продемонстрировали специфическое обогащение инсулина или трансферрина в сочетании с их родственными рецепторами INSR и IGF1R или TFRC, соответственно (, рис. 3e, дополнительная таблица 2 и , дополнительные данные 3 ). LUX-MS, управляемая трансферрином, также выявила CD22, специфичный для В-клеток лектин, который, как было показано, слабо связывается с сиалилированными N-гликопротеинами, включая трансферрин 57 , и катион-независимый маннозо-6-фосфатный рецептор M6PR, сортирующий фермент, участвующий в клатрин-опосредованный эндоцитоз и рециркуляция трансферрин-связанных рецепторов, которые, как было показано, совместно локализуются с TFRC перед интернализацией в сортирующие эндосомы 58 .Таким образом, в дополнение к малым молекулам, LUX-MS можно использовать для раскрытия сигнальных доменов поверхностных биомолекул, обеспечивая молекулярное понимание событий межклеточной коммуникации на большие расстояния.
3e, дополнительная таблица 2 и , дополнительные данные 3 ). LUX-MS, управляемая трансферрином, также выявила CD22, специфичный для В-клеток лектин, который, как было показано, слабо связывается с сиалилированными N-гликопротеинами, включая трансферрин 57 , и катион-независимый маннозо-6-фосфатный рецептор M6PR, сортирующий фермент, участвующий в клатрин-опосредованный эндоцитоз и рециркуляция трансферрин-связанных рецепторов, которые, как было показано, совместно локализуются с TFRC перед интернализацией в сортирующие эндосомы 58 .Таким образом, в дополнение к малым молекулам, LUX-MS можно использовать для раскрытия сигнальных доменов поверхностных биомолекул, обеспечивая молекулярное понимание событий межклеточной коммуникации на большие расстояния.
Сигнальные взаимодействия Surfaceome могут быть обнаружены в эукариотических и прокариотических системах
Маркировка близости белков и обнаружение с помощью LUX-MS в основном основаны на индуцированных светом модификациях гистидина и триптофана, чей клеточный биосинтез эволюционно хорошо законсервирован 59,60 . Таким образом, LUX-MS может обнаруживать взаимодействия лиганд-поверхностный сигнал за пределами эукариотических систем, таких как бактерии, где LRC-подобные подходы неприменимы, решая критический шаг в разработке противоинфекционных мер против растущей угрозы лекарственно-устойчивых бактерий. Недавно было показано, что встречающийся в природе пептид Танатин проявляет противомикробную активность, связываясь с LptA и в конечном итоге блокируя комплекс биогенеза наружной мембраны Lpt в Escherichia coli ( E.coli ) 61 . Чтобы проверить способность LUX-MS идентифицировать критические взаимодействия поверхности у бактерий, мы соединили танатин с генератором синглетного кислорода метиленовым синим (, рис. 4a, ) и добавили его к клеткам E. coli с конкуренцией или без нее путем избытка неконъюгированный танатин. Затем все клетки освещали в течение 5 минут красным светом с длиной волны 656 нм и подвергали LUX-MS для идентификации белков с близкой меткой.
Таким образом, LUX-MS может обнаруживать взаимодействия лиганд-поверхностный сигнал за пределами эукариотических систем, таких как бактерии, где LRC-подобные подходы неприменимы, решая критический шаг в разработке противоинфекционных мер против растущей угрозы лекарственно-устойчивых бактерий. Недавно было показано, что встречающийся в природе пептид Танатин проявляет противомикробную активность, связываясь с LptA и в конечном итоге блокируя комплекс биогенеза наружной мембраны Lpt в Escherichia coli ( E.coli ) 61 . Чтобы проверить способность LUX-MS идентифицировать критические взаимодействия поверхности у бактерий, мы соединили танатин с генератором синглетного кислорода метиленовым синим (, рис. 4a, ) и добавили его к клеткам E. coli с конкуренцией или без нее путем избытка неконъюгированный танатин. Затем все клетки освещали в течение 5 минут красным светом с длиной волны 656 нм и подвергали LUX-MS для идентификации белков с близкой меткой. Многочисленные белки внешней мембраны и периплазмы были значительно обогащены (рис.4b, дополнительная таблица 3 и , дополнительные данные 4 ), что указывает на локализованное в мембране мечение танатином, связанным с SOG. Таким образом, LUX-MS выявила функциональные партнеры по связыванию Thanatin LptA и LptD как максимально обогащенные хиты и предположила близость к белкам внешней мембраны, включая BamA и BamC (, рис. 4b, ). В самом деле, комплекс Bam, как было показано, катализирует сворачивание и встраивание белков наружной мембраны, включая LptD, образующих надмолекулярные поверхностные кластеры на живых бактериях -62-.Таким образом, LUX-MS позволяет проводить деконволюцию мишени на поверхности и картировать биомолекулярные сигнальные взаимодействия в доменах жизни.
Многочисленные белки внешней мембраны и периплазмы были значительно обогащены (рис.4b, дополнительная таблица 3 и , дополнительные данные 4 ), что указывает на локализованное в мембране мечение танатином, связанным с SOG. Таким образом, LUX-MS выявила функциональные партнеры по связыванию Thanatin LptA и LptD как максимально обогащенные хиты и предположила близость к белкам внешней мембраны, включая BamA и BamC (, рис. 4b, ). В самом деле, комплекс Bam, как было показано, катализирует сворачивание и встраивание белков наружной мембраны, включая LptD, образующих надмолекулярные поверхностные кластеры на живых бактериях -62-.Таким образом, LUX-MS позволяет проводить деконволюцию мишени на поверхности и картировать биомолекулярные сигнальные взаимодействия в доменах жизни.
Рис. 4 Безгипотезная идентификация сигнальных взаимодействий поверхностиома в прокариотических системах.
a Схема соединения пептидомиметического антибиотика Танатина с генератором синглетного кислорода (СОГ) тиородамином. b , график Volcano, показывающий относительное количество белков, определенных LUX-MS, из обработанных Thanatin-SOG Escherichia coli , освещенных в течение 15 минут с конкуренцией Thanatin и без нее.Синие точки представляют собой значительно обогащенные белки, а красные точки представляют собой мишени прямого связывания танатина, ранее обнаруженные в экспериментах по фотосшиванию. Выделены основные цели и выбранные взаимодействующие поверхности. c , схематическое изображение, описывающее молекулярный механизм действия танатина в клетках E.coli в пространственном контексте. Список MS-количественных белков ( дополнительная таблица 3 ) и полностью интерактивный график вулкана ( дополнительные данные 4 ) предоставляются для облегчения исследования данных.
b , график Volcano, показывающий относительное количество белков, определенных LUX-MS, из обработанных Thanatin-SOG Escherichia coli , освещенных в течение 15 минут с конкуренцией Thanatin и без нее.Синие точки представляют собой значительно обогащенные белки, а красные точки представляют собой мишени прямого связывания танатина, ранее обнаруженные в экспериментах по фотосшиванию. Выделены основные цели и выбранные взаимодействующие поверхности. c , схематическое изображение, описывающее молекулярный механизм действия танатина в клетках E.coli в пространственном контексте. Список MS-количественных белков ( дополнительная таблица 3 ) и полностью интерактивный график вулкана ( дополнительные данные 4 ) предоставляются для облегчения исследования данных.
Распутывание сети поверхностных взаимодействий интактных вирусных частиц с помощью LUX-MS
Внеклеточные взаимодействия хозяина и патогена являются ключевыми сигнальными событиями, которые опосредуют клеточный тропизм и инфекцию, и, как таковые, являются основными объектами исследований для классификации и диагностики инфекционных заболеваний и вакцин открытие. Поэтому мы объединили тиородамин SOG с бактериофагами, которые распознают вездесущий компонент клеточной стенки тейхоевую кислоту грамположительных бактерий Listeria monocytogenes ( Lm ), высокоинфекционного пищевого патогена, вызывающего листериоз 63 ( Fig.5а ). Lm инкубировали с фагами, связанными с SOG, при множественности заражения (MOI) 1 или 10 и метили LUX при освещении от 0 до 15 мин. Анализ проточной цитометрии выявил светозависимое биотинилирование клеточной поверхности отдельных бактерий, которое достигло насыщения после 15 минут освещения при MOI 10 (, рис. 5b, ). Последующий анализ LUX-MS выявил индуцированное светом значительное обогащение 146 белками, включая 19 фаговых и 127 белков Lm (рис.5c, дополнительная таблица 4 и дополнительные данные 5 ). Обнаружены компоненты капсида бактериофага (Cps, Gp5, Gp9), оболочки (Gp3), базовой пластинки (Gp20) и хвоста (Gp17, Gp18, Gp19, Gp23, Tmp, Tsh), а также ферменты, участвующие в связывании ДНК (Gp37), репликации.
Поэтому мы объединили тиородамин SOG с бактериофагами, которые распознают вездесущий компонент клеточной стенки тейхоевую кислоту грамположительных бактерий Listeria monocytogenes ( Lm ), высокоинфекционного пищевого патогена, вызывающего листериоз 63 ( Fig.5а ). Lm инкубировали с фагами, связанными с SOG, при множественности заражения (MOI) 1 или 10 и метили LUX при освещении от 0 до 15 мин. Анализ проточной цитометрии выявил светозависимое биотинилирование клеточной поверхности отдельных бактерий, которое достигло насыщения после 15 минут освещения при MOI 10 (, рис. 5b, ). Последующий анализ LUX-MS выявил индуцированное светом значительное обогащение 146 белками, включая 19 фаговых и 127 белков Lm (рис.5c, дополнительная таблица 4 и дополнительные данные 5 ). Обнаружены компоненты капсида бактериофага (Cps, Gp5, Gp9), оболочки (Gp3), базовой пластинки (Gp20) и хвоста (Gp17, Gp18, Gp19, Gp23, Tmp, Tsh), а также ферменты, участвующие в связывании ДНК (Gp37), репликации. (Gp45), рекомбинации (Gp44), транскрипции (Gp7) и иммунозащитных функций (Gp32), что указывает на тщательную маркировку LUX бактериофагов, прикрепленных к клетке-хозяину. Для белков Lm онтологический анализ генов выявил 81 внеклеточный и только 8 внутриклеточных белков (рис.5d ), включая основные факторы вирулентности, такие как интерналины A и B, связанные с клеточной стенкой, которые опосредуют инвазию клеток-хозяев, локализованный в мембране ActA, который регулирует межклеточное распространение, и переносчики АТФ-связывающей кассеты (ABC), которые опосредуют множественную лекарственную устойчивость 64 ( рис. 5e ). Используя HA-метку и иммунофлуоресцентную микроскопию, мы дополнительно подтвердили поверхностную экспрессию Lm предполагаемого белка клеточной стенки и транспортера ABC, которые были сильно обогащены LUX-MS ( Fig.5f ). В целом, наши результаты демонстрируют способность LUX-MS распутывать взаимодействия интактных вирусов с поверхностью хозяина в масштабе всего протеома, что позволяет изучать внеклеточные взаимодействия хозяина и патогена, что в конечном итоге облегчает разработку противоинфекционных стратегий.
(Gp45), рекомбинации (Gp44), транскрипции (Gp7) и иммунозащитных функций (Gp32), что указывает на тщательную маркировку LUX бактериофагов, прикрепленных к клетке-хозяину. Для белков Lm онтологический анализ генов выявил 81 внеклеточный и только 8 внутриклеточных белков (рис.5d ), включая основные факторы вирулентности, такие как интерналины A и B, связанные с клеточной стенкой, которые опосредуют инвазию клеток-хозяев, локализованный в мембране ActA, который регулирует межклеточное распространение, и переносчики АТФ-связывающей кассеты (ABC), которые опосредуют множественную лекарственную устойчивость 64 ( рис. 5e ). Используя HA-метку и иммунофлуоресцентную микроскопию, мы дополнительно подтвердили поверхностную экспрессию Lm предполагаемого белка клеточной стенки и транспортера ABC, которые были сильно обогащены LUX-MS ( Fig.5f ). В целом, наши результаты демонстрируют способность LUX-MS распутывать взаимодействия интактных вирусов с поверхностью хозяина в масштабе всего протеома, что позволяет изучать внеклеточные взаимодействия хозяина и патогена, что в конечном итоге облегчает разработку противоинфекционных стратегий.
Рис. 5. Картирование нацеленных на вирусы поверхностных клеток живых хозяев с использованием LUX-MS под контролем бактериофагов.
a . Схема соединения Listeria monocytogenes ( Lm )-специфического бактериофага с генератором синглетного кислорода (SOG) тиородамином. b , График гистограммы, показывающий светозависимое биотинилирование клеточной поверхности клеток Lm после LUX-мечения бактериофагом-SOG. c , График вулкана, показывающий относительное количество белков, определенных LUX-MS, из обработанных бактериофагом-SOG Lm с освещением и без него в течение 15 мин. Синие и зеленые точки представляют собой значительно обогащенные Lm — или фаговые белки соответственно, в то время как серые точки указывают на необогащенные белки. Кандидаты, отобранные для микроскопической характеристики, выделены. d , GO-term анализ обогащения значительно обогащенных белков Lm . e , Схематическое представление пространственного охвата LUX-MS на основе субклеточных аннотаций значительно обогащенных белков. Кандидаты, отобранные для микроскопии, отмечены стрелками. f , Репрезентативные изображения конфокальной микроскопии Lm (1042), оставленного необработанным и окрашенным для определения клеточной поверхности Internalin-A (зеленый), или трансфицированным плазмидой, не содержащей (pPL2) или меченных HA белков-кандидатов, и окрашенным для определения HA клеточной поверхности (Красный).Список MS-количественных белков ( дополнительная таблица 4 ) и полностью интерактивный график вулкана ( дополнительные данные 5 ) предоставляются для облегчения исследования данных.
Выяснение межклеточных сигнальных доменов поверхности в функциональных иммуносинапсах
Контакт-зависимая межклеточная коммуникация является важным регулятором функции иммунной системы и опирается на очень сложные межклеточные сигнальные архитектуры поверхности. «иммунологический синапс» 6 .Контекстное взаимодействие иммуносинаптических рецепторных взаимодействий между APC и CD8 + Т-клетками приводит к активации, пролиферации и дифференцировке Т-клеток, позволяя этим клеткам проявлять эффекторные функции против инфицированных, онкогенных или чужеродных клеток 65 . Таким образом, способность разрешать функциональные иммуносинапсы на молекулярном уровне с пространственно-временной специфичностью позволит сделать открытия в области иммунологии и иммуноонкологии. Таким образом, мы соединили тиородамин SOG с пептидом gp33-41, полученным из вируса лимфоцитарного хориоменингита (LCMV) (gp33, последовательность: KAVYNFATM) ( Fig.6a ), иммунодоминантный эпитоп гликопотеина LCMV (GP), который представлен MHC класса I H-2D b и H-2K b 66 и который распознается трансгенным Т-клеточным рецептором (TCR) CD8 + Т-клетки мышей P14 67 . В двухклеточной системе мы показали, что gp33, связанный с SOG, все еще представлен дендритными клетками мыши и способен сравнительно активировать свежевыделенные P14 CD8 + T-клетки, чем его неконъюгированный аналог ( Supplementary Fig.
«иммунологический синапс» 6 .Контекстное взаимодействие иммуносинаптических рецепторных взаимодействий между APC и CD8 + Т-клетками приводит к активации, пролиферации и дифференцировке Т-клеток, позволяя этим клеткам проявлять эффекторные функции против инфицированных, онкогенных или чужеродных клеток 65 . Таким образом, способность разрешать функциональные иммуносинапсы на молекулярном уровне с пространственно-временной специфичностью позволит сделать открытия в области иммунологии и иммуноонкологии. Таким образом, мы соединили тиородамин SOG с пептидом gp33-41, полученным из вируса лимфоцитарного хориоменингита (LCMV) (gp33, последовательность: KAVYNFATM) ( Fig.6a ), иммунодоминантный эпитоп гликопотеина LCMV (GP), который представлен MHC класса I H-2D b и H-2K b 66 и который распознается трансгенным Т-клеточным рецептором (TCR) CD8 + Т-клетки мышей P14 67 . В двухклеточной системе мы показали, что gp33, связанный с SOG, все еще представлен дендритными клетками мыши и способен сравнительно активировать свежевыделенные P14 CD8 + T-клетки, чем его неконъюгированный аналог ( Supplementary Fig. 8 ). Применение света привело к биотинилированию клеточной поверхности обоих типов клеток, подтверждая фотокаталитическую функцию gp33, связанного с SOG, и нашу способность эффективно выполнять трансклеточную маркировку поверхности во время иммуносинаптических межклеточных контактов (, дополнительная рис. 9 ). Затем мы внедрили мечение стабильным изотопом аминокислотами в клеточной культуре (SILAC) для изотопного штрих-кодирования взаимодействующих дендритных клеток (DC) и Т-клеток (TC) и выполнили LUX-MS в режиме независимого от данных (DIA) после 30 минут совместной работы. -культура для анализа зрелых иммунологических синапсов ( Fig.6б ). В целом, мы идентифицировали 4811 белковых групп, которые могут быть четко отнесены к TC (например, TCR, CD3 и CD8), DC (например, MHC класса I H-2D b и H-2K b ) или обоим на основе наблюдаемых данных. Соотношения SILAC (, рис. 6c, и , дополнительная таблица 5, ).
8 ). Применение света привело к биотинилированию клеточной поверхности обоих типов клеток, подтверждая фотокаталитическую функцию gp33, связанного с SOG, и нашу способность эффективно выполнять трансклеточную маркировку поверхности во время иммуносинаптических межклеточных контактов (, дополнительная рис. 9 ). Затем мы внедрили мечение стабильным изотопом аминокислотами в клеточной культуре (SILAC) для изотопного штрих-кодирования взаимодействующих дендритных клеток (DC) и Т-клеток (TC) и выполнили LUX-MS в режиме независимого от данных (DIA) после 30 минут совместной работы. -культура для анализа зрелых иммунологических синапсов ( Fig.6б ). В целом, мы идентифицировали 4811 белковых групп, которые могут быть четко отнесены к TC (например, TCR, CD3 и CD8), DC (например, MHC класса I H-2D b и H-2K b ) или обоим на основе наблюдаемых данных. Соотношения SILAC (, рис. 6c, и , дополнительная таблица 5, ). Мы наблюдали светозависимое обогащение многочисленных белков клеточной поверхности, включая компоненты первичной сигнальной оси, такие как MHC класса I на DC и gp33-специфический TCR, а также CD8 на TC (, рис. 6d-e, дополнительная таблица 5, дополнительная Данные 6 и 7 ).Кроме того, мы обнаружили костимулирующие сигнальные компоненты, необходимые для раннего праймирования Т-клеток, такие как CD86 оси CD86-CD28 и всей оси CD2-CD48 с сообщенной клеточной направленностью 6 . Наши результаты также указывают на иммуносинаптическую локализацию CD5 и CD6 на TC. Действительно, было показано, что CD5 взаимодействует с TCR, CD8 и CD2 в иммунных синапсах для точной настройки опосредованной TCR передачи сигналов 68 , в то время как структурно родственный CD6 физически связывается с CD5 и его лигандом CD166 (также известным как ALCAM) 69 , который также был захвачен в округе Колумбия.LUX-MS дополнительно выявил иммуносинаптические адгезивные комплексы, включая LFA-1 (TC), ICAM-1 (DC) и VLA-4 (оба), которые усиливают межклеточную близость и активацию Т-клеток 65 , и идентифицировал тетраспанины CD9, CD81, сильно встроенные в мембрану.
Мы наблюдали светозависимое обогащение многочисленных белков клеточной поверхности, включая компоненты первичной сигнальной оси, такие как MHC класса I на DC и gp33-специфический TCR, а также CD8 на TC (, рис. 6d-e, дополнительная таблица 5, дополнительная Данные 6 и 7 ).Кроме того, мы обнаружили костимулирующие сигнальные компоненты, необходимые для раннего праймирования Т-клеток, такие как CD86 оси CD86-CD28 и всей оси CD2-CD48 с сообщенной клеточной направленностью 6 . Наши результаты также указывают на иммуносинаптическую локализацию CD5 и CD6 на TC. Действительно, было показано, что CD5 взаимодействует с TCR, CD8 и CD2 в иммунных синапсах для точной настройки опосредованной TCR передачи сигналов 68 , в то время как структурно родственный CD6 физически связывается с CD5 и его лигандом CD166 (также известным как ALCAM) 69 , который также был захвачен в округе Колумбия.LUX-MS дополнительно выявил иммуносинаптические адгезивные комплексы, включая LFA-1 (TC), ICAM-1 (DC) и VLA-4 (оба), которые усиливают межклеточную близость и активацию Т-клеток 65 , и идентифицировал тетраспанины CD9, CD81, сильно встроенные в мембрану. , CD82 и CD151 на DC, которые организуют иммуносинаптические сигнальные домены на антигенпрезентирующих клетках 70 . Напротив, вездесущий рецептор протеинтирозинфосфатазы типа C (PTPRC, CD45) не был идентифицирован на TC в соответствии с его иммуносинаптическим исключением, позволяющим фосфорилировать TCR и передавать сигналы 65 ( Fig.6е ). В совокупности наши результаты демонстрируют полезность LUX-MS для всестороннего разрешения межклеточных сигнальных архитектур поверхностных функциональных иммунных синапсов с клеточной специфичностью и пространственно-временным контролем света, обеспечивая молекулярную основу для открытия лекарств в иммунологии и иммуноонкологии.
, CD82 и CD151 на DC, которые организуют иммуносинаптические сигнальные домены на антигенпрезентирующих клетках 70 . Напротив, вездесущий рецептор протеинтирозинфосфатазы типа C (PTPRC, CD45) не был идентифицирован на TC в соответствии с его иммуносинаптическим исключением, позволяющим фосфорилировать TCR и передавать сигналы 65 ( Fig.6е ). В совокупности наши результаты демонстрируют полезность LUX-MS для всестороннего разрешения межклеточных сигнальных архитектур поверхностных функциональных иммунных синапсов с клеточной специфичностью и пространственно-временным контролем света, обеспечивая молекулярную основу для открытия лекарств в иммунологии и иммуноонкологии.
Рис. 6 Выяснение межклеточных сигнальных доменов поверхности в функциональных иммуносинапсах.
a , схема соединения иммуногенного пептида gp33 с генератором синглетного кислорода (SOG) тиородамином.Показана кристаллическая структура gp33 (темно-серый), представленная альфа-цепью MHC класса I H-2D B (красный) в комплексе с бета-2-микроглобулином (темно-красный) (идентификатор PDB: 1FG2) b , Схема рабочий процесс LUX-MS на основе SILAC-DIA. Меченые изотопами (тяжелые) мышиные дендритные клетки, представляющие gp33-SOG2, устанавливают синхронизированные иммуносинапсы с изолированными (легкими), gp33-специфичными CD8 + Т-клетками, что позволяет активировать световое мечение in situ и молекулярный анализ межклеточных сигнальных взаимодействий поверхностома в синапсах с использованием масс-спектрометрии с независимым сбором данных (DIA). c , График гистограммы, показывающий распределение соотношений содержания тяжелых и легких белков, идентифицированных с помощью LUX-MS. Показаны границы соотношений для определения типа клеток и репрезентативных белков. d , График вулкана, показывающий относительное количество белков, определенных LUX-MS, из двухклеточных систем с освещением и без него в течение 15 минут. Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Красные и синие точки представляют собой белки, полученные из дендритных клеток (DC) и Т-клеток (TC), соответственно.
Меченые изотопами (тяжелые) мышиные дендритные клетки, представляющие gp33-SOG2, устанавливают синхронизированные иммуносинапсы с изолированными (легкими), gp33-специфичными CD8 + Т-клетками, что позволяет активировать световое мечение in situ и молекулярный анализ межклеточных сигнальных взаимодействий поверхностома в синапсах с использованием масс-спектрометрии с независимым сбором данных (DIA). c , График гистограммы, показывающий распределение соотношений содержания тяжелых и легких белков, идентифицированных с помощью LUX-MS. Показаны границы соотношений для определения типа клеток и репрезентативных белков. d , График вулкана, показывающий относительное количество белков, определенных LUX-MS, из двухклеточных систем с освещением и без него в течение 15 минут. Точки и крестики обозначают клеточную поверхность и другие аннотированные белки соответственно. Красные и синие точки представляют собой белки, полученные из дендритных клеток (DC) и Т-клеток (TC), соответственно. Известные иммуносинаптические компоненты выделены прямыми взаимодействующими элементами gp33, показанными в прямоугольниках. e . Схема LUX-MS, идентифицированная сетью иммуносинаптических поверхностных взаимодействий. f . Схема, подчеркивающая взаимосвязь иммунологического синапса с эндолизосомными путями в Т-клетках и показывающая белки на межклеточном интерфейсе с помощью LUX-MS. Список MS-количественных белков ( дополнительная таблица 5 ) и полностью интерактивный график вулкана ( дополнительные данные 6 и 7 ) предоставляются для облегчения исследования данных.
Известные иммуносинаптические компоненты выделены прямыми взаимодействующими элементами gp33, показанными в прямоугольниках. e . Схема LUX-MS, идентифицированная сетью иммуносинаптических поверхностных взаимодействий. f . Схема, подчеркивающая взаимосвязь иммунологического синапса с эндолизосомными путями в Т-клетках и показывающая белки на межклеточном интерфейсе с помощью LUX-MS. Список MS-количественных белков ( дополнительная таблица 5 ) и полностью интерактивный график вулкана ( дополнительные данные 6 и 7 ) предоставляются для облегчения исследования данных.
Обсуждение
Наноразмерная организация и сеть внеклеточных взаимодействий клеточного поверхностного слоя является ключевым сигнальным медиатором в действии лекарств, вирусной инфекции и межклеточной коммуникации. В этом исследовании мы разработали и применили основанную на SOG и управляемую светом технологию обнаружения близости, называемую LUX-MS, которая позволяет выяснить наноразмерную организацию поверхности и ее сигнальные доамины поверхности, нацеленные на лиганд, на живых клетках. В отличие от более крупных ферментов (APEX2: 28 кДа; miniTurbo: 27 кДа) 71 использование низкомолекулярных SOG (< 1 кДа) в качестве меченых зондов дает следующие преимущества: профилирование поверхностного взаимодействия лигандов практически любого описания, начиная от малых молекул, антител и вирусных частиц и заканчивая целыми клетками.ii) Ковалентное связывание SOG с совместимой с лигандом химией, сохраняющей структурно-функциональные отношения и исключающей необходимость генетических манипуляций. iii) Контролируемая светом генерация синглетного кислорода, позволяющая точно настраивать маркировку на месте с жестким пространственно-временным контролем. iv) Мечение белков с помощью консервативных аминокислотных остатков, позволяющее исследовать поверхности всего протеома в широком диапазоне эволюционно различных организмов. Недавно фотокатализаторы, продуцирующие короткоживущий карбен с наносекундным периодом полураспада, были использованы для картирования белковых микродоменов под контролем антител, которые не могли быть разрешены с помощью стратегий на основе ферментов (промежуточные продукты с периодом полураспада 0.
В отличие от более крупных ферментов (APEX2: 28 кДа; miniTurbo: 27 кДа) 71 использование низкомолекулярных SOG (< 1 кДа) в качестве меченых зондов дает следующие преимущества: профилирование поверхностного взаимодействия лигандов практически любого описания, начиная от малых молекул, антител и вирусных частиц и заканчивая целыми клетками.ii) Ковалентное связывание SOG с совместимой с лигандом химией, сохраняющей структурно-функциональные отношения и исключающей необходимость генетических манипуляций. iii) Контролируемая светом генерация синглетного кислорода, позволяющая точно настраивать маркировку на месте с жестким пространственно-временным контролем. iv) Мечение белков с помощью консервативных аминокислотных остатков, позволяющее исследовать поверхности всего протеома в широком диапазоне эволюционно различных организмов. Недавно фотокатализаторы, продуцирующие короткоживущий карбен с наносекундным периодом полураспада, были использованы для картирования белковых микродоменов под контролем антител, которые не могли быть разрешены с помощью стратегий на основе ферментов (промежуточные продукты с периодом полураспада 0.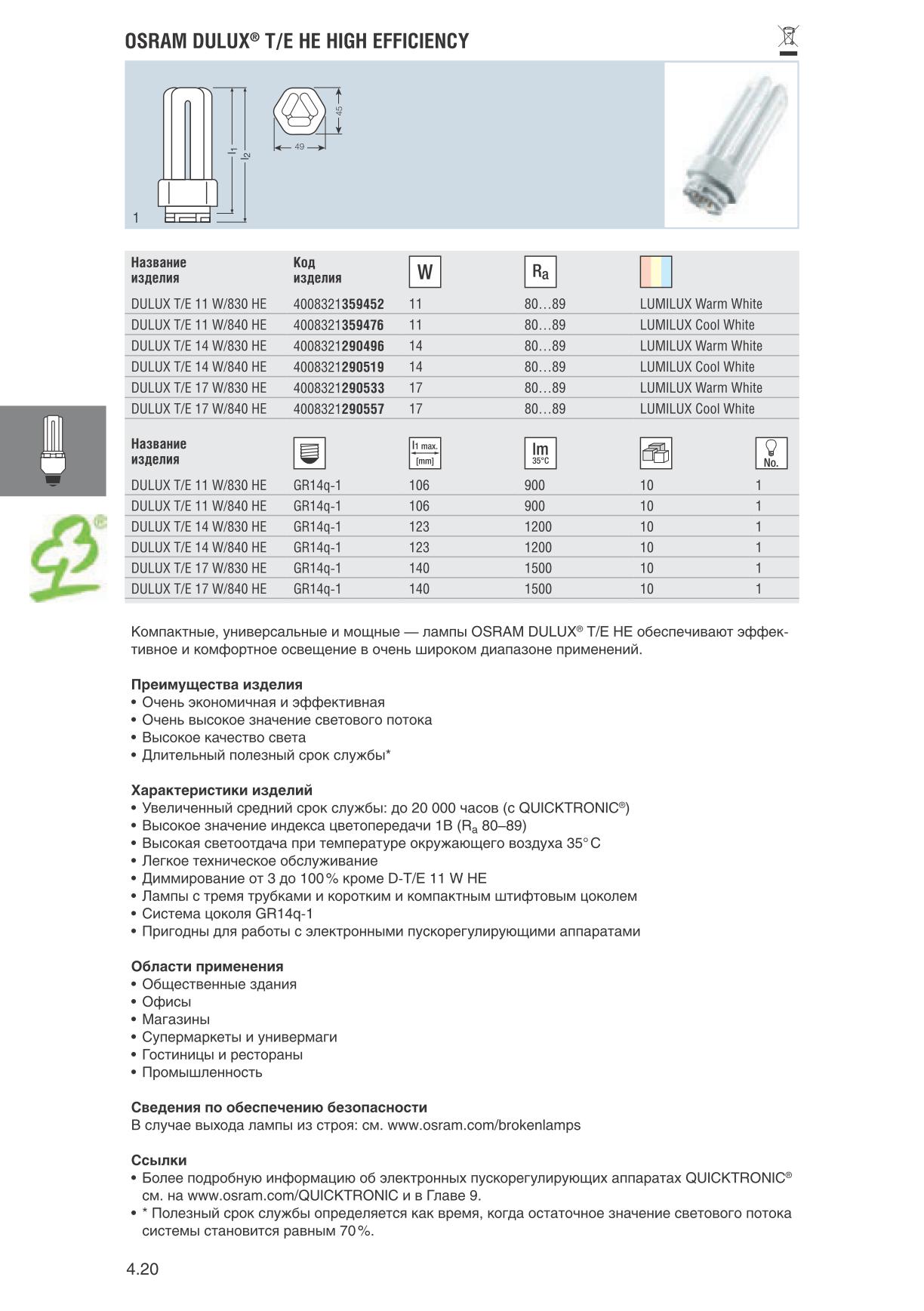 от 1 мс до 60 с) 27 . Маркировка близости с помощью LUX-MS опосредована фотосенсибилизированными синглетными формами кислорода со временем жизни 25–36 мкс 43 , что указывает на пространственное разрешение между подходами на основе ферментов и карбенов. Тем не менее, прямое связывание SOG с сигнальными компетентными лигандами ограничивает мечение сигнальными релевантными поверхностными доменами и позволяет избежать использования антител, которые влекут за собой структурную гибкость. Таким образом, LUX-MS дополняет доступные в настоящее время технологии и обеспечивает аналитическую ступеньку для общесистемного выяснения ландшафта клеточной поверхности.
от 1 мс до 60 с) 27 . Маркировка близости с помощью LUX-MS опосредована фотосенсибилизированными синглетными формами кислорода со временем жизни 25–36 мкс 43 , что указывает на пространственное разрешение между подходами на основе ферментов и карбенов. Тем не менее, прямое связывание SOG с сигнальными компетентными лигандами ограничивает мечение сигнальными релевантными поверхностными доменами и позволяет избежать использования антител, которые влекут за собой структурную гибкость. Таким образом, LUX-MS дополняет доступные в настоящее время технологии и обеспечивает аналитическую ступеньку для общесистемного выяснения ландшафта клеточной поверхности.
Используя антитела для маркировки близости, мы продемонстрировали способность LUX-MS специфически выявлять различные области близости поверхности на живых клетках, которые формируют молекулярный каркас для реконструкции организации поверхности в наномасштабе. Учитывая широкую доступность высококачественных антител для установленных поверхностных мишеней и высокую пропускную способность рабочего процесса LUX-MS, возможно глубокое картирование взаимодействия поверхности, предоставляя информацию на уровне клеточной поверхности, которой в настоящее время не хватает в текущих усилиях системной биологии по картированию и понимать структуру и функции интерактома человека 72–75 .
Применяя LUX-MS к малым молекулам, мы картировали поверхностные сигнальные домены низкомолекулярного сердечного гликозида CG1. В отличие от методологий захвата лиганд-рецептор, LUX-MS не только обеспечивает целевую деконволюцию малых молекул в масштабах всего протеома, но также предоставляет информацию о близости поверхностного клеточного типа, которая может быть использована для адресной доставки лекарств, например. с использованием EDC 47 . Хотя ожидается, что этот подход будет применим к другим лекарственным средствам, накопление знаний о клеточных поверхностомах, нацеленных на лекарственные средства, в конечном итоге позволит идентифицировать связанные с заболеванием проксимиты поверхностных веществ, что позволит создать прецизионную медицину следующего поколения, которая выходит далеко за рамки нацеливания на отдельные рецепторы.
С помощью LUX-MS мы расшифровали взаимодействия вирусной поверхности в бактериальных хозяевах, которые невозможно обнаружить с помощью современных технологий, таких как CSC, LRC и HATRIC, из-за структурной сложности клеточной оболочки и общего отсутствия гликозилирование белков. Используя бактериофаги, которые распознают вездесущий компонент клеточной стенки Listeria monocytogenes , мы нанесли на карту бактериальную поверхность (> 80 белков) со значительным охватом и специфичностью, которые заменили обычные стратегии профилирования, такие как трипсиновое бритье (идентифицировано 11 белков), поверхностно-ограниченное биотинилирование. идентифицировано 27 белков) и фракционирование клеток (идентифицировано 21 белок) 76 .В зависимости от специфичности связывания используемой вирусной частицы, LUX-MS позволяет определять в пространстве и времени сайты прикрепления вирусов, рецепторы-мишени и глобальные поверхностомы живых прокариотических и эукариотических клеток в сложных биологических системах. Это представляет особый интерес, учитывая широкое применение передовых анализов фагового дисплея 77 и соответствующих платформ скрининга 5,78 в фундаментальных и трансляционных исследованиях.
Используя бактериофаги, которые распознают вездесущий компонент клеточной стенки Listeria monocytogenes , мы нанесли на карту бактериальную поверхность (> 80 белков) со значительным охватом и специфичностью, которые заменили обычные стратегии профилирования, такие как трипсиновое бритье (идентифицировано 11 белков), поверхностно-ограниченное биотинилирование. идентифицировано 27 белков) и фракционирование клеток (идентифицировано 21 белок) 76 .В зависимости от специфичности связывания используемой вирусной частицы, LUX-MS позволяет определять в пространстве и времени сайты прикрепления вирусов, рецепторы-мишени и глобальные поверхностомы живых прокариотических и эукариотических клеток в сложных биологических системах. Это представляет особый интерес, учитывая широкое применение передовых анализов фагового дисплея 77 и соответствующих платформ скрининга 5,78 в фундаментальных и трансляционных исследованиях.
Наконец, мы использовали LUX-MS для пространственно-временного анализа межклеточных сигнальных доменов поверхности в иммунологических синапсах. Предыдущие попытки ограничивались молекулярным анализом в модельных системах, в которых использовались растворимые, связанные с планшетами или ассоциированные с мембраной факторы для имитации активации иммуносинаптических Т-клеток 65 , или флуоресцентным отчетом о транзиторных межклеточных взаимодействиях иммунных клеток 27,79 . Здесь мы выполнили LUX-MS, управляемую иммуногеном gp33, в двухклеточной системе с изотопным штрих-кодом, состоящей из миллионов синхронизированных и функциональных иммуносинапсов, и таким образом получили целостное представление о зрелой иммуносинаптической сигнальной архитектуре поверхности, что способствовало формулированию новых биологических гипотез.Например, мы обнаружили несколько лизосомных белков (LAMP-1, LAMP-2, CD68 и катепсины B, D и Z) в иммуносинаптической щели на наивных CD8 + Т-клетках, но не антигенпрезентирующих дендритных клетках (). Дополнительная таблица 5 ). Это указывает на локальное слияние лизосомальных гранул в Т-клетках, которое обычно наблюдается в цитотоксических Т-клетках во время опосредованного дегрануляцией уничтожения клеток-мишеней 80 .
Предыдущие попытки ограничивались молекулярным анализом в модельных системах, в которых использовались растворимые, связанные с планшетами или ассоциированные с мембраной факторы для имитации активации иммуносинаптических Т-клеток 65 , или флуоресцентным отчетом о транзиторных межклеточных взаимодействиях иммунных клеток 27,79 . Здесь мы выполнили LUX-MS, управляемую иммуногеном gp33, в двухклеточной системе с изотопным штрих-кодом, состоящей из миллионов синхронизированных и функциональных иммуносинапсов, и таким образом получили целостное представление о зрелой иммуносинаптической сигнальной архитектуре поверхности, что способствовало формулированию новых биологических гипотез.Например, мы обнаружили несколько лизосомных белков (LAMP-1, LAMP-2, CD68 и катепсины B, D и Z) в иммуносинаптической щели на наивных CD8 + Т-клетках, но не антигенпрезентирующих дендритных клетках (). Дополнительная таблица 5 ). Это указывает на локальное слияние лизосомальных гранул в Т-клетках, которое обычно наблюдается в цитотоксических Т-клетках во время опосредованного дегрануляцией уничтожения клеток-мишеней 80 .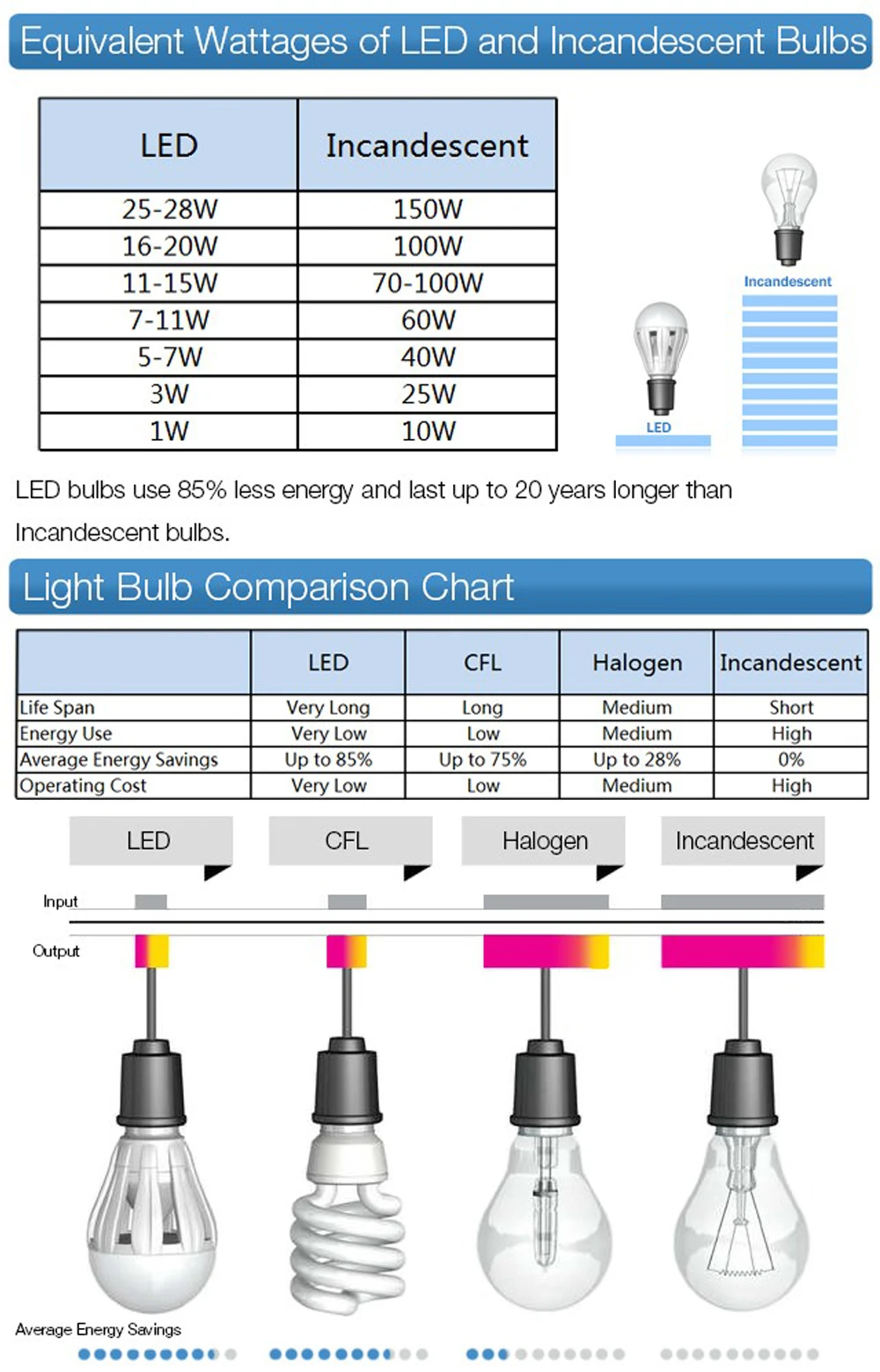 Однако было показано, что, хотя дегрануляция и, следовательно, цитолитическая активность являются признаком клеток, подвергшихся воздействию антигена, они отсутствуют в наивных CD8 + Т-клетках.Действительно, мы не идентифицировали какие-либо цитолитические эффекторные молекулы, такие как перфорин или гранзимы, что указывает на секрецию нелитических гранул в наивных CD8 + Т-клетках в течение 30 минут после первоначального контакта с антигенпрезентирующими клетками (, рис. 6f, ). . Несмотря на то, что клеточный механизм для иммуносинаптически направленного транспорта везикул запускается в течение 6 минут после первоначального клеточного контакта, точный механизм такой нелитической секреции гранул и его функциональная роль в антиген-зависимом праймировании Т-клеток еще предстоит изучить. .
Однако было показано, что, хотя дегрануляция и, следовательно, цитолитическая активность являются признаком клеток, подвергшихся воздействию антигена, они отсутствуют в наивных CD8 + Т-клетках.Действительно, мы не идентифицировали какие-либо цитолитические эффекторные молекулы, такие как перфорин или гранзимы, что указывает на секрецию нелитических гранул в наивных CD8 + Т-клетках в течение 30 минут после первоначального контакта с антигенпрезентирующими клетками (, рис. 6f, ). . Несмотря на то, что клеточный механизм для иммуносинаптически направленного транспорта везикул запускается в течение 6 минут после первоначального клеточного контакта, точный механизм такой нелитической секреции гранул и его функциональная роль в антиген-зависимом праймировании Т-клеток еще предстоит изучить. .
В заключение, мы представляем высокопроизводительную оптопротеомную технологию, называемую LUX-MS, для обнаружения белковых сетей в сигнальных доменах поверхностных клеток на живых клетках, и мы демонстрируем ее полезность и универсальность, распутывая наноразмерную организацию поверхности и сигнальные взаимодействия, лежащие в основе действия лекарств, хозяина и хозяина. Взаимодействие патогенов и межклеточная коммуникация. Учитывая биологическую и терапевтическую важность сигнального ландшафта поверхности, мы предполагаем, что LUX-MS повлияет на фундаментальные и прикладные области исследований, раскрывая пространственно-временную взаимосвязь сигнальных шлюзов клеточной поверхности в норме и болезни.
Взаимодействие патогенов и межклеточная коммуникация. Учитывая биологическую и терапевтическую важность сигнального ландшафта поверхности, мы предполагаем, что LUX-MS повлияет на фундаментальные и прикладные области исследований, раскрывая пространственно-временную взаимосвязь сигнальных шлюзов клеточной поверхности в норме и болезни.
Методы
Реагенты
Все химикаты были приобретены у Merck, если не указано иное. Генераторы синглетного кислорода в виде NHS-функционализированного тиородамина (кат.: AD-Thio12-41) и метиленового синего, функционализированного азидом (кат.: AD-MB2-31), были приобретены у ATTO-TEC GmbH (Зиген, Германия). Биоцитин-гидразид был приобретен у Pitsch Nucleic Acids AG (Stein am Rhein, Швейцария). Для экспериментов с LUX-MS использовали следующие антитела: контрольное антитело изотипа IgG мыши (Invitrogen, кат.: 10400C), моноклональное антитело IgG2 мыши против CD20, клон 2H7 (Invitrogen, кат.: 14-0209-82), антитело против CD38 человека. химерное моноклональное антитело, любезно предоставленное Centrose LLC, моноклональное антитело IgG1 мыши против CD54 человека, клон HCD54 (BioLegend, кат. № 322704), моноклональное антитело IgG1 мыши против CD166 человека, клон 3A6 (BioLegend, кат. № 343902), антитело против CD220 человека мышиное моноклональное антитело IgG2, клон B6.220 (BioLegend, кат.: 352602).
химерное моноклональное антитело, любезно предоставленное Centrose LLC, моноклональное антитело IgG1 мыши против CD54 человека, клон HCD54 (BioLegend, кат. № 322704), моноклональное антитело IgG1 мыши против CD166 человека, клон 3A6 (BioLegend, кат. № 343902), антитело против CD220 человека мышиное моноклональное антитело IgG2, клон B6.220 (BioLegend, кат.: 352602).
Культура клеток
Все химические вещества для культивирования клеток были приобретены у ThermoFisher Scientific, если не указано иное. Клеточные линии были приобретены в ATCC и выращены при 37°C и 5% CO 2 окружающей среды. Полученную от пациента клеточную линию В-лимфомы SU-DHL-6 (ATCC, CRL-2959) выращивали в среде 1640 Мемориального института Розуэлл-Парк (RPMI) с 1,5 мМ GlutaMAX, 1% пенициллина-стрептомицина и 10% эмбриональной телячьей сыворотки. Полученную от пациентов клеточную линию промиелоцитарного лейкоза HL60 (ATCC CCL-240) выращивали в модифицированной среде Дульбекко Искова (IMDM) с 1. 5 мМ GlutaMAX, 1% пенициллин-стрептомицин и 10% эмбриональная бычья сыворотка. Линия дендритных клеток мыши MutuDC1940 81 была любезно предоставлена Хансом Ача-Орбеа (кафедра биохимии, Лозаннский университет, Швейцария) и была культивирована и изотопно помечена в модифицированной среде Дульбекко Искова (IMDM) для SILAC (Thermo, кат.: 88367) с глюкозой (конечная: 4,5 г/л), L-Arg- 13 C 6 — 15 N 4 (конечная: 42 мкг/мл), L-Lys- 13 C 6 — 15 N 2 (конечная: 73 мкг/мл), 1.5 мМ GlutaMAX, 10 мМ HEPES, 50 мкМ β-меркаптоэтанола, 1% пенициллин-стрептомицина, заменимых аминокислот и 10% диализированной эмбриональной бычьей сыворотки.
5 мМ GlutaMAX, 1% пенициллин-стрептомицин и 10% эмбриональная бычья сыворотка. Линия дендритных клеток мыши MutuDC1940 81 была любезно предоставлена Хансом Ача-Орбеа (кафедра биохимии, Лозаннский университет, Швейцария) и была культивирована и изотопно помечена в модифицированной среде Дульбекко Искова (IMDM) для SILAC (Thermo, кат.: 88367) с глюкозой (конечная: 4,5 г/л), L-Arg- 13 C 6 — 15 N 4 (конечная: 42 мкг/мл), L-Lys- 13 C 6 — 15 N 2 (конечная: 73 мкг/мл), 1.5 мМ GlutaMAX, 10 мМ HEPES, 50 мкМ β-меркаптоэтанола, 1% пенициллин-стрептомицина, заменимых аминокислот и 10% диализированной эмбриональной бычьей сыворотки.
Мыши
Самок или самцов мышей в возрасте от шести до шестнадцати недель использовали для экспериментов на животных, описанных в этом исследовании. Мыши CD45.1 P14 с Т-клеточными рецепторами на CD8 + Т-клетки, специфичные к эпитопу гликопротеина GP33-41 вируса лимфоцитарного хориоменингита (LCMV) 67 и CD45. 1 P14, скрещенные с мышами Nur77-GFP 82 , были выведены в Центре Феномики ETH.Всех животных перед использованием разводили и содержали в условиях, свободных от специфических патогенов. Все эксперименты на животных проводились в соответствии с институциональной политикой и швейцарскими федеральными нормами, в соответствии с руководящими принципами и были одобрены ветеринарным управлением кантона Цюрих (разрешение на эксперименты на животных: 115/2017).
1 P14, скрещенные с мышами Nur77-GFP 82 , были выведены в Центре Феномики ETH.Всех животных перед использованием разводили и содержали в условиях, свободных от специфических патогенов. Все эксперименты на животных проводились в соответствии с институциональной политикой и швейцарскими федеральными нормами, в соответствии с руководящими принципами и были одобрены ветеринарным управлением кантона Цюрих (разрешение на эксперименты на животных: 115/2017).
Светоконтролируемое производство синглетного кислорода
Генератор синглетного кислорода (SOG) тиородамин смешивали с 1 мкМ Singlet Oxygen Sensor Green (Invitrogen, кат.: S36002) в фосфатно-солевом буфере при повышении D 2 O/H 2 O и освещены на рабочем расстоянии 30 см прожекторами Precision LED с длиной волны 590 нм, управляемыми модулем управления освещением BioLED (Mightex Systems) в течение от 0 до 15 минут.Флуоресценцию фотоокисленного SOSG контролировали с помощью многорежимного планшетного ридера Synergy HT (BioTek) при возбуждении 485 ± 20 нм и эмиссии 528 ± 20 нм. Количественные значения интенсивности были стерты и нормализованы к неосвещенному контролю с использованием индивидуального R-скрипта.
Количественные значения интенсивности были стерты и нормализованы к неосвещенному контролю с использованием индивидуального R-скрипта.
Открытие индуцированных светом модификаций белка
Голотрансферрин человека (Sigma, кат. : 89882) в соответствии с рекомендациями производителя.SOG-связанный трансферрин в D 2 PBS на основе O, содержащий 0 или 5 мМ биоцитин-гидразида, освещали прожекторами Precision LED с длиной волны 590 нм, управляемыми через модуль управления источником света BioLED (Mightex Systems, Плезантон, США) в течение 15 мин. или оставить в темноте. Буфер дополняли 50 мМ катализатором 2-(аминометил)имидазолдигидрохлорида в течение 50 мин. Обработанные белки расщепляли в течение ночи при 37 °C с использованием трипсина для секвенирования (Promega, кат.: V511C) при соотношении фермента к белку 1:100.Пептиды очищали по C18 с использованием колонок UltraMicroSpin 5–60 мкг (The Nest Group, кат.: SEMSS18V) в соответствии с инструкциями производителя и подвергали масс-спектрометрическому анализу с использованием масс-спектрометра Orbitrap Fusion Tribrid (Thermo Scientific) в режиме сбора данных (DDA). режим с массовым разрешением 120 000 и 30 000 на уровне предшественника и фрагмента соответственно (подробности см. Ниже). Полученные необработанные файлы освещенных образцов были подвергнуты открытому поиску модификаций с помощью MSfragger (v.201
режим с массовым разрешением 120 000 и 30 000 на уровне предшественника и фрагмента соответственно (подробности см. Ниже). Полученные необработанные файлы освещенных образцов были подвергнуты открытому поиску модификаций с помощью MSfragger (v.201
 Пептиды были количественно определены путем интеграции хроматографических следов с использованием Progenesis QI v.4.0 (Nonlinear Dynamics) и отфильтрованы до уровня ложных обнаружений <1%. Пептиды трансферрина были отнесены к фиктивным белкам, представляющим немодифицированные типы или типы с одной модификацией, которые были статистически проверены на дифференциальную распространенность с использованием пакета R MSstats (v.3.8.6) 85 . Типы модификаций с кратностью изменения численности > 1.5 при освещении и скорректированном значении p <0,05 считались продуктами фотоокисления, индуцированного светом. Аналогичным образом модификации аминокислот со значительно сниженной светозависимой продукцией в присутствии биотин-гидразида (p-значение <0,1) считались модификациями аминокислот, реагирующими с гидразидами.
Пептиды были количественно определены путем интеграции хроматографических следов с использованием Progenesis QI v.4.0 (Nonlinear Dynamics) и отфильтрованы до уровня ложных обнаружений <1%. Пептиды трансферрина были отнесены к фиктивным белкам, представляющим немодифицированные типы или типы с одной модификацией, которые были статистически проверены на дифференциальную распространенность с использованием пакета R MSstats (v.3.8.6) 85 . Типы модификаций с кратностью изменения численности > 1.5 при освещении и скорректированном значении p <0,05 считались продуктами фотоокисления, индуцированного светом. Аналогичным образом модификации аминокислот со значительно сниженной светозависимой продукцией в присутствии биотин-гидразида (p-значение <0,1) считались модификациями аминокислот, реагирующими с гидразидами.Сбор и очистка мононуклеарных клеток периферической крови человека (МКПК)
Лейкоцитарная пленка была получена от здоровых доноров, предоставленных Службой переливания крови Цюриха. Для выделения PBMC образцы лейкоцитарной пленки человека разводили 1:1 в PBS (Gibco) и выделяли мононуклеарные клетки с градиентом плотности Histopaque-1077 (Sigma-Aldrich) в соответствии с инструкциями производителя. РВМС на границе раздела собирали, один раз промывали в PBS и ресуспендировали в среде до дальнейшей обработки.
Для выделения PBMC образцы лейкоцитарной пленки человека разводили 1:1 в PBS (Gibco) и выделяли мононуклеарные клетки с градиентом плотности Histopaque-1077 (Sigma-Aldrich) в соответствии с инструкциями производителя. РВМС на границе раздела собирали, один раз промывали в PBS и ресуспендировали в среде до дальнейшей обработки.
Поверхностное наномасштабное картирование под контролем антител
На образец 10 мкг антител связывали с тиородамином SOG при молярном соотношении антитело:SOG 1:5 и очищали с использованием 7-кДа колонок ZebaSpin (Thermo, кат.: 89882) в соответствии с инструкцией производителя. рекомендации.Конструкции антител, связанных с SOG, хранили при 4°C и немедленно использовали. Двадцать миллионов клеток инкубировали с 10 мкг SOG-связанного антитела в течение 30 минут при 4 °C в темноте, чтобы свести к минимуму вызванное фоновым светом окисление. Клетки промывали ледяным PBS, ресуспендировали в охлажденном буфере для фотоокисления (5 мМ биоцитин-гидразид в PBS на основе D 2 O, pH 7,5) и освещали на рабочем расстоянии 20 см прецизионным светодиодом с длиной волны 590 нм. прожекторы или оставлены в темноте. Для временной кривой LUX-MS анти-CD20 анализировали один и три биологически независимых образца для каждой временной точки освещения и контроля без освещения соответственно.Для анти-CD20 LUX-MS анализировали два и три биологически независимых образца в условиях меченого и немеченого соответственно. Для всех других LUX-MS, управляемых антителами, были проанализированы три биологически независимых образца как для меченых, так и для немеченых условий. Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения (5 мМ биоцитин-гидразид и 50 мМ 2-(аминометил)имидазолдигидрохлорид в PBS, pH 6,0) в течение 50 мин при 4 °C в темноте. Для проточного цитометрического анализа биотинилирования клеточной поверхности клетки тщательно промывали PBS и окрашивали NeutrAvidin Protein DyLight 488 (Invitrogen, кат. в.10.07). Таким образом, паттерн FSC/SSC немеченого состояния использовали для селекции живых клеток. Для анализа LUX-MS клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при температуре -80 °C до дальнейшей обработки.
прожекторы или оставлены в темноте. Для временной кривой LUX-MS анти-CD20 анализировали один и три биологически независимых образца для каждой временной точки освещения и контроля без освещения соответственно.Для анти-CD20 LUX-MS анализировали два и три биологически независимых образца в условиях меченого и немеченого соответственно. Для всех других LUX-MS, управляемых антителами, были проанализированы три биологически независимых образца как для меченых, так и для немеченых условий. Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения (5 мМ биоцитин-гидразид и 50 мМ 2-(аминометил)имидазолдигидрохлорид в PBS, pH 6,0) в течение 50 мин при 4 °C в темноте. Для проточного цитометрического анализа биотинилирования клеточной поверхности клетки тщательно промывали PBS и окрашивали NeutrAvidin Protein DyLight 488 (Invitrogen, кат. в.10.07). Таким образом, паттерн FSC/SSC немеченого состояния использовали для селекции живых клеток. Для анализа LUX-MS клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при температуре -80 °C до дальнейшей обработки.
Синтез сердечного гликозида CG1, связанного с генератором синглетного кислорода
Сердечный гликозид CG1, содержащий первичный амин, был любезно предоставлен Centrose LLC (Madison, Wisconsin). К раствору CG1 2 (5 мг, 0,0081 ммоль) и эфира тиородамин-NHS 1 (5.7 мг, 0,0081 ммоль) в N,N-диметилформамиде (ДМФ) (1 мл) добавляли Et 3 N (0,011 мл, 0,08 ммоль). Реакционную смесь перемешивали при комнатной температуре в течение 1 часа, и образование продукта подтверждали с помощью ЖХ-МС. Конечный продукт CG1-SOG 3 очищали препаративной ОФ-ВЭЖХ на колонке Waters Eclipse XDB (C18, 250 x 21,2 мм, 7 мкм) с градиентом 5 – 50% ACN/H 2 O + 0,1 % TFA в 5 объемах колонки. Продукт имел чистоту> 95%, судя по обращенно-фазовой ВЭЖХ и HR-MS (, дополнительная рис.10 ). HR-ESI-MS: m/z (M+H + ) 999,4938 (вычисленная масса = 999,4936).
Идентификация низкомолекулярных структур поверхности, нацеленных на лекарственное средство
Пятьдесят миллионов клеток инкубировали с 0–10 мкМ CG1-SOG (10 мкМ для эксперимента LUX-MS) в течение 30 мин при 4 °C в темноте, чтобы свести к минимуму фоновую засветку. индуцированное окисление. Клетки промывали ледяным PBS, ресуспендировали в охлажденном буфере для фотоокисления и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм или оставляли в темноте (по три биологически независимых образца в каждом).Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Для проточного цитометрического анализа биотинилирования клеточной поверхности клетки тщательно промывали PBS и окрашивали NeutrAvidin Protein DyLight 488 (Invitrogen, кат.: 22832) 1:200 в течение 20 минут перед анализом с использованием проточного цитометра Accuri C6 (BD Biosciences) и FlowJo ( v.10.07). Таким образом, паттерн FSC/SSC немеченого состояния использовали для селекции живых клеток. Для анализа LUX-MS клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при температуре -80 °C до дальнейшей обработки.
индуцированное окисление. Клетки промывали ледяным PBS, ресуспендировали в охлажденном буфере для фотоокисления и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм или оставляли в темноте (по три биологически независимых образца в каждом).Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Для проточного цитометрического анализа биотинилирования клеточной поверхности клетки тщательно промывали PBS и окрашивали NeutrAvidin Protein DyLight 488 (Invitrogen, кат.: 22832) 1:200 в течение 20 минут перед анализом с использованием проточного цитометра Accuri C6 (BD Biosciences) и FlowJo ( v.10.07). Таким образом, паттерн FSC/SSC немеченого состояния использовали для селекции живых клеток. Для анализа LUX-MS клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при температуре -80 °C до дальнейшей обработки.
Открытие межклеточных поверхностных сигнальных доменов с помощью биомолекул
Человеческий пептидный гормон инсулин и голотрансферрин гликопротеина были соединены с тиородамином SOG при молярном соотношении лиганд:SOG 1:1 и 1:13, соответственно, в соответствии с рекомендациями производителя. Конструкции лиганда, связанного с SOG, хранили при 4°C и немедленно использовали. Двадцать миллионов клеток инкубировали с 1 мкМ инсулина-SOG или 75 нМ трансферрина-SOG в течение 10 минут при 4°C в темноте, чтобы свести к минимуму вызванное фоновым светом окисление.Клетки промывали ледяным PBS, ресуспендировали в охлажденном фотоокислительном буфере и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм в течение 5 мин или оставляли в темноте (каждый по три биологически независимых образца). Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при -80 °C до дальнейшей обработки.
Конструкции лиганда, связанного с SOG, хранили при 4°C и немедленно использовали. Двадцать миллионов клеток инкубировали с 1 мкМ инсулина-SOG или 75 нМ трансферрина-SOG в течение 10 минут при 4°C в темноте, чтобы свести к минимуму вызванное фоновым светом окисление.Клетки промывали ледяным PBS, ресуспендировали в охлажденном фотоокислительном буфере и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм в течение 5 мин или оставляли в темноте (каждый по три биологически независимых образца). Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при -80 °C до дальнейшей обработки.
Синтез и характеристика противомикробного препарата танатина, связанного с генератором синглетного кислорода
Производное танатина было любезно предоставлено группой Джона Робинсона (Цюрихский университет, Швейцария) и (6. 0 мг, 0,0023 ммоль) растворяли в 500 мкл H 2 O. Аскорбат натрия (0,1 М, 30 мкл, 0,0022 ммоль) и азид-функционализированный метиленовый синий в качестве генератора синглетного кислорода (0,5 мг, 0,0009 ммоль) в 100 мкл Добавляли ДМСО и CuSO 4 (0,1 М, 30 мкл, 0,003 ммоль). Через 15 мин реакционную смесь вводили непосредственно в колонку препаративной ВЭЖХ C18 с градиентом 10-40% ACN/H 2 O (0,1% TFA). Чистота соединения танатин-SOG была подтверждена анализом UPLC, а идентичность подтверждена HR-ESI с расчетной массой 652.1353 (M+6H) 6+ и измеренное значение m/z 652,1360 (M+6H) 6+ ( Дополнительный рисунок 11 ).
0 мг, 0,0023 ммоль) растворяли в 500 мкл H 2 O. Аскорбат натрия (0,1 М, 30 мкл, 0,0022 ммоль) и азид-функционализированный метиленовый синий в качестве генератора синглетного кислорода (0,5 мг, 0,0009 ммоль) в 100 мкл Добавляли ДМСО и CuSO 4 (0,1 М, 30 мкл, 0,003 ммоль). Через 15 мин реакционную смесь вводили непосредственно в колонку препаративной ВЭЖХ C18 с градиентом 10-40% ACN/H 2 O (0,1% TFA). Чистота соединения танатин-SOG была подтверждена анализом UPLC, а идентичность подтверждена HR-ESI с расчетной массой 652.1353 (M+6H) 6+ и измеренное значение m/z 652,1360 (M+6H) 6+ ( Дополнительный рисунок 11 ).
Деконволюция всего протеома и активируемая светом мишень в прокариотических системах
Клетки Escherichia coli (Migula) Castellani and Chalmers (ATCC: 25922) инкубировали с 3 мкМ Thanatin-SOG в присутствии или в отсутствие 30 мкM неконъюгированного Thanatin производное в течение 30 минут при 37 ° C в темноте, чтобы свести к минимуму окисление, вызванное фоновым светом. Клетки промывали ледяным PBS, ресуспендировали в охлажденном фотоокислительном буфере и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 659 нм в течение 5 мин или оставляли в темноте (каждый по три биологически независимых образца). Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при -80 °C до дальнейшей обработки.
Клетки промывали ледяным PBS, ресуспендировали в охлажденном фотоокислительном буфере и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 659 нм в течение 5 мин или оставляли в темноте (каждый по три биологически независимых образца). Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при -80 °C до дальнейшей обработки.
Генерация и характеристика бактериофагов, связанных с генератором синглетного кислорода
Грамположительные L.monocytogenes специфические бактериофаги связывали с тиородамином SOG путем смешивания 1,2 мл раствора фага Listeria A500 (титр 10 12 бляшкообразующих единиц/мл) с 60 мкл 0,2 М NaHCO 3 и 30 мкл 0,2 М NaHCO 3 и 30 мкл. Тиородамин, функционализированный NHS. Смесь инкубировали при комнатной температуре в темноте в течение 1 часа. Комплекс фаг-SOG осаждали добавлением 1/5 объема 20% ПЭГ/2,5 М NaCl и инкубировали в течение 2 часов при 4°С. Супернатант отбрасывали для удаления непрореагировавших молекул SOG, а осадок красителя М13 ресуспендировали в 1 мл буфера SM и подвергали определению бляшкообразующих единиц (БОЕ).Для анализа БОЕ 990 мкл ночной культуры клеток L. monocytogenes смешивали с 10 мкл серийно разведенного исходного раствора фага (в буфере SM). Затем культуру смешивали и встряхивали с 5 мл расплавленного мягкого 1/2 BHI-агара (50°C) и выливали на чашки с твердым агаром. Чашки с затвердевшим агаром инкубировали при 30 °C в течение дня и определяли титр фага на основе количества наблюдаемых бляшек.
Комплекс фаг-SOG осаждали добавлением 1/5 объема 20% ПЭГ/2,5 М NaCl и инкубировали в течение 2 часов при 4°С. Супернатант отбрасывали для удаления непрореагировавших молекул SOG, а осадок красителя М13 ресуспендировали в 1 мл буфера SM и подвергали определению бляшкообразующих единиц (БОЕ).Для анализа БОЕ 990 мкл ночной культуры клеток L. monocytogenes смешивали с 10 мкл серийно разведенного исходного раствора фага (в буфере SM). Затем культуру смешивали и встряхивали с 5 мл расплавленного мягкого 1/2 BHI-агара (50°C) и выливали на чашки с твердым агаром. Чашки с затвердевшим агаром инкубировали при 30 °C в течение дня и определяли титр фага на основе количества наблюдаемых бляшек.
Изучение поверхностных клеток-мишеней хозяина под контролем бактериофагов
Клетки Listeria monocytogenes (штамм WSLC 1042) обрабатывали 40 мкг/мл гентамицина в течение 1 часа при комнатной температуре перед инкубацией с SOG-связанными Listeria
8 при aphages A5 множественность заражения (MOI) 10 в течение 30 мин при 4°С в темноте. На образец бактерии, соответствующие 35 x 10 9 колониеобразующих единиц (КОЕ), промывали ледяным PBS, ресуспендировали в охлажденном буфере для фотоокисления и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм. на 15 мин или оставляли в темноте (по три биологически независимых образца). Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Для проточного цитометрического анализа биотинилирования клеточной поверхности клетки тщательно промывали PBS и окрашивали NeutrAvidin Protein DyLight 488 1:50 в течение 20 минут перед анализом с использованием проточного цитометра Accuri C6 (BD Biosciences) с боковым рассеянием (высота), установленным на 10. ‘000 в качестве предела обнаружения и FlowJo (v.10.07). Таким образом, паттерн FSC/SSC немеченого состояния использовали для селекции живых клеток. Для анализа LUX-MS клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при температуре -80 °C до дальнейшей обработки.
На образец бактерии, соответствующие 35 x 10 9 колониеобразующих единиц (КОЕ), промывали ледяным PBS, ресуспендировали в охлажденном буфере для фотоокисления и освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм. на 15 мин или оставляли в темноте (по три биологически независимых образца). Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Для проточного цитометрического анализа биотинилирования клеточной поверхности клетки тщательно промывали PBS и окрашивали NeutrAvidin Protein DyLight 488 1:50 в течение 20 минут перед анализом с использованием проточного цитометра Accuri C6 (BD Biosciences) с боковым рассеянием (высота), установленным на 10. ‘000 в качестве предела обнаружения и FlowJo (v.10.07). Таким образом, паттерн FSC/SSC немеченого состояния использовали для селекции живых клеток. Для анализа LUX-MS клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при температуре -80 °C до дальнейшей обработки.
Генная инженерия и проверка под микроскопом идентифицированных поверхностных белков
L. monocytogenes
Гены-кандидаты, кодирующие предполагаемые поверхностные белки A0A0E1RAE1 (белок якорного домена клеточной стенки с мотивом LPXTG) и A0A0E1R383 (переносчик ABC), а также их нативные промоторы были клонированы в pPL2 векторы интеграции 63 с меткой HA с использованием метода Гибсона, как описано ранее 86 .Пустой вектор pPL2, два вектора, содержащих вставку, pPL2( 05780 -HA) и pPL2( 06470 -HA), трансформировали в электрокомпетентные клетки L. monocytogenes WSLC 1042. Праймеры 5′-GTCAAAACATACGCTCTTATC-3′ и 5′-ACATAATCAGTCCAAAGTAGATGC-3′ использовали для проверки интеграции pPL2 в бактериальный геном. Для поверхностного иммунного окрашивания штаммов L. monocytogenes WT и HA, меченных HA, соответствующие клетки выращивали до средней логарифмической фазы, промывали PBS и обрабатывали 100 мкг/мл лизоцима в течение 30 минут, чтобы сделать поверхностные белки более доступными для иммунного окрашивания, а затем блокируется в 1% BSA/PBS. Клетки далее промывали и окрашивали поликлональным антителом InlA (LSBio, Inc) или моноклональным антителом HA, конъюгированным с Alexa Fluor 555 (ThermoFisher Scientific) в концентрации 1:100 в течение 30 мин перед визуализацией с помощью конфокальной микроскопии. Для окрашивания InlA клетки снова промывали в PBS и дополнительно инкубировали с конъюгированным с Alexa Fluor 488 козьим антикроличьим вторичным антителом. После последней стадии промывки клеточные суспензии наносили на предметное стекло и визуализировали с помощью конфокальной микроскопии.
Клетки далее промывали и окрашивали поликлональным антителом InlA (LSBio, Inc) или моноклональным антителом HA, конъюгированным с Alexa Fluor 555 (ThermoFisher Scientific) в концентрации 1:100 в течение 30 мин перед визуализацией с помощью конфокальной микроскопии. Для окрашивания InlA клетки снова промывали в PBS и дополнительно инкубировали с конъюгированным с Alexa Fluor 488 козьим антикроличьим вторичным антителом. После последней стадии промывки клеточные суспензии наносили на предметное стекло и визуализировали с помощью конфокальной микроскопии.
Синтез и характеристика иммуногенного пептида gp33, связанного с генератором синглетного кислорода
Пептид gp33-41 LCMV (gp33; KAVYNFATM) был приобретен у NeoMPS.Пептид gp33 (1,0 мг, 1,0 мкмоль, 1 экв.) и NHS-тиородамин (1,4 мг, 2,0 мкмоль, 2 экв.) инкубировали при комнатной температуре в течение 90 мин в PBS при pH 8,3. Реакционную смесь очищали непосредственно с помощью препаративной обращенно-фазовой высокоэффективной жидкостной хроматографии (ОФ-ВЭЖХ) на приборе Jasco с использованием колонки YMC C18 (5 мкм, 20 мм ID x 250 мм) при комнатной температуре при скорости потока 10 мл. /мин, с одновременным контролем элюента при 220, 254 и 301 нм. Вода Milli-Q с 0,1% TFA (растворитель A) и градиент от 5 до 15% ACN с 0.В качестве элюента использовали 1% ТФУ (растворитель В) в течение 5 минут и от 15 до 75% растворителя В в течение 30 минут. Фракции, содержащие чистый продукт, объединяли и лиофилизировали с получением gp33-SOG 2 в виде фиолетового твердого вещества. Чистота и идентичность были проверены с помощью аналитической ОФ-ВЭЖХ и ВР-МС (, дополнительная рис. 12, ). HR-MS (ESI): рассчитан для C 106 N 133 N 17 O 17 S 3 9 3 3 2+ [M] 2+ : 1005.9608, найдено: 1005.9589.
/мин, с одновременным контролем элюента при 220, 254 и 301 нм. Вода Milli-Q с 0,1% TFA (растворитель A) и градиент от 5 до 15% ACN с 0.В качестве элюента использовали 1% ТФУ (растворитель В) в течение 5 минут и от 15 до 75% растворителя В в течение 30 минут. Фракции, содержащие чистый продукт, объединяли и лиофилизировали с получением gp33-SOG 2 в виде фиолетового твердого вещества. Чистота и идентичность были проверены с помощью аналитической ОФ-ВЭЖХ и ВР-МС (, дополнительная рис. 12, ). HR-MS (ESI): рассчитан для C 106 N 133 N 17 O 17 S 3 9 3 3 2+ [M] 2+ : 1005.9608, найдено: 1005.9589.
Функциональная оценка генератора синглетного кислорода gp33
CD8 + Т-клетки были выделены из разрушенных селезенки и паховых лимфатических узлов CD45.1 х мышей Nur77-GFP с использованием набора для выделения Т-клеток EasySep(tm), не подвергавшихся воздействию мышей CD8 + (StemCell, Гренобль, Франция), в соответствии с инструкциями производителя. Селезенки и лимфатические узлы разрушали с помощью 70-микрометрового клеточного фильтра. Меченные изотопами мышиные дендритные клетки инкубировали без соединения или с 1 мкг/мл очищенного gp33, не несущего (gp33), одного (gp33-SOG) или двух (gp33-SOG 2 ) генераторов синглетного кислорода в течение 1 часа. Центрифугированием добавляли 10-кратный избыток остро выделенных CD8 + Т-клеток и коинкубировали клетки в течение 3 ч в присутствии интерлейкина-2.Клетки осторожно промывали ледяным PBS, добавляли охлажденный буфер для фотоокисления и взаимодействующие клетки освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм в течение 15 мин. Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS и оценивали на активацию Т-клеток и биотинилирование специфической поверхности клеток с использованием проточной цитометрии.
Селезенки и лимфатические узлы разрушали с помощью 70-микрометрового клеточного фильтра. Меченные изотопами мышиные дендритные клетки инкубировали без соединения или с 1 мкг/мл очищенного gp33, не несущего (gp33), одного (gp33-SOG) или двух (gp33-SOG 2 ) генераторов синглетного кислорода в течение 1 часа. Центрифугированием добавляли 10-кратный избыток остро выделенных CD8 + Т-клеток и коинкубировали клетки в течение 3 ч в присутствии интерлейкина-2.Клетки осторожно промывали ледяным PBS, добавляли охлажденный буфер для фотоокисления и взаимодействующие клетки освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм в течение 15 мин. Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS и оценивали на активацию Т-клеток и биотинилирование специфической поверхности клеток с использованием проточной цитометрии.
Активация светом
in situ Маркировка функциональных иммуносинапсов
На образец десять миллионов меченных изотопами дендритных клеток мыши были предварительно активированы 0. 5 мкг/мл CpG (Microsynth, Balgach, Швейцария) в течение ночи. Промытые PBS дендритные клетки затем инкубировали с 1 мкг/мл gp33-SOG 2 в течение 1 ч в темноте, чтобы свести к минимуму вызванное фоновым светом окисление. Центрифугированием добавляли 1,5-кратный избыток свежевыделенных CD8 + Т-клеток и коинкубировали клетки в течение 30 мин в присутствии интерлейкина-2. Клетки осторожно промывали ледяным PBS, добавляли охлажденный фотоокислительный буфер и взаимодействующие клетки освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм в течение 15 мин или оставляли в темноте (два и три биологически независимых образцы соответственно).Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при -80 °C до дальнейшей обработки.
5 мкг/мл CpG (Microsynth, Balgach, Швейцария) в течение ночи. Промытые PBS дендритные клетки затем инкубировали с 1 мкг/мл gp33-SOG 2 в течение 1 ч в темноте, чтобы свести к минимуму вызванное фоновым светом окисление. Центрифугированием добавляли 1,5-кратный избыток свежевыделенных CD8 + Т-клеток и коинкубировали клетки в течение 30 мин в присутствии интерлейкина-2. Клетки осторожно промывали ледяным PBS, добавляли охлажденный фотоокислительный буфер и взаимодействующие клетки освещали на рабочем расстоянии 20 см прожекторами Precision LED с длиной волны 590 нм в течение 15 мин или оставляли в темноте (два и три биологически независимых образцы соответственно).Клетки осаждали центрифугированием и ресуспендировали в охлажденном буфере для мечения в течение 50 мин при 4°С в темноте. Клетки тщательно промывали PBS, быстро замораживали в виде осадка клеток в жидком азоте и хранили при -80 °C до дальнейшей обработки.
Анализ мышиных иммунных клеток проточной цитометрией
Собранные клетки окрашивали для проточной цитометрии в PBS в течение 20 мин при комнатной температуре. Следующие антитела были приобретены у BioLegend (Сан-Диего, США) и использованы для проточного цитометрического анализа: CD8a-BV510 (53-6.7), CD45.1-PacificBlue (A20), CD44-APC (IM7), CD11c-PerCP (N418), CD69-PeCy7 (h2.2F3), CD25-FITC (3C7). Жизнеспособность клеток определяли с помощью фиксируемого окрашивания мертвых клеток в ближней ИК-области (LifeTechnologies, Карлсбад, США). Биотинилирование оценивали окрашиванием стрептавидином-PE (BioLegend) в течение 10 мин при комнатной температуре. Активацию Т-клеток определяли по экспрессии экспрессии GFP под контролем промотора Nur77. Данные были получены на проточном цитометре Canto ™ (BD Bioscience, Allschwil, Швейцария) и проанализированы с использованием программного обеспечения FlowJo (Treestar, Ashland, OR, USA).
Следующие антитела были приобретены у BioLegend (Сан-Диего, США) и использованы для проточного цитометрического анализа: CD8a-BV510 (53-6.7), CD45.1-PacificBlue (A20), CD44-APC (IM7), CD11c-PerCP (N418), CD69-PeCy7 (h2.2F3), CD25-FITC (3C7). Жизнеспособность клеток определяли с помощью фиксируемого окрашивания мертвых клеток в ближней ИК-области (LifeTechnologies, Карлсбад, США). Биотинилирование оценивали окрашиванием стрептавидином-PE (BioLegend) в течение 10 мин при комнатной температуре. Активацию Т-клеток определяли по экспрессии экспрессии GFP под контролем промотора Nur77. Данные были получены на проточном цитометре Canto ™ (BD Bioscience, Allschwil, Швейцария) и проанализированы с использованием программного обеспечения FlowJo (Treestar, Ashland, OR, USA).
Автоматический захват и обработка белков
Осадок фотомеченых клеток лизировали в 500 мкл лизирующего буфера (100 мМ Трис, 1% дезоксихолат натрия, 10 мМ Трис(2-карбоксиэтил)фосфин (TCEP), 15 мМ 2-хлорацетамид). обработкой ультразвуком в течение четырех интервалов по 30 секунд в VialTweeter (Hielscher Ultrasonics) при мощности 170 Вт при 80% времени цикла и последующем нагреве до 100 °С в течение 5 мин. Буфер для лизиса бактериальных клеток дополнительно содержал 75 мкг эндолизинов Ply511 Listeria для снижения целостности клеточной стенки.Концентрацию белка определяли с помощью спектрофотометра Nanodrop 2000 (Thermo Fisher Scientific), и равные количества между образцом и контрольным состоянием (обычно 3 мг белка) подвергали автоматическому захвату и обработке фотомеченых белков с использованием робота для работы с жидкостями. В частности, упакованные наконечники собственного производства, содержащие 80 мкл смолы Streptavidin Plus UltraLink (Thermo Fisher Scientific), были подключены к системе обработки жидкости Versette (Thermo Fisher Scientific) для автоматического смешивания с клеточным лизатом для 2.5 ч и последующая промывка 5 М NaCl, буфер StimLys (50 мМ Трис pH 7,8, 137 мМ NaCl, 150 мМ глицерин, 0,5 мМ ЭДТА, 0,1% Тритон Х-100), 100 мМ NaHCO 3 и 50 мМ (NH 4 ) HCO 3 .
обработкой ультразвуком в течение четырех интервалов по 30 секунд в VialTweeter (Hielscher Ultrasonics) при мощности 170 Вт при 80% времени цикла и последующем нагреве до 100 °С в течение 5 мин. Буфер для лизиса бактериальных клеток дополнительно содержал 75 мкг эндолизинов Ply511 Listeria для снижения целостности клеточной стенки.Концентрацию белка определяли с помощью спектрофотометра Nanodrop 2000 (Thermo Fisher Scientific), и равные количества между образцом и контрольным состоянием (обычно 3 мг белка) подвергали автоматическому захвату и обработке фотомеченых белков с использованием робота для работы с жидкостями. В частности, упакованные наконечники собственного производства, содержащие 80 мкл смолы Streptavidin Plus UltraLink (Thermo Fisher Scientific), были подключены к системе обработки жидкости Versette (Thermo Fisher Scientific) для автоматического смешивания с клеточным лизатом для 2.5 ч и последующая промывка 5 М NaCl, буфер StimLys (50 мМ Трис pH 7,8, 137 мМ NaCl, 150 мМ глицерин, 0,5 мМ ЭДТА, 0,1% Тритон Х-100), 100 мМ NaHCO 3 и 50 мМ (NH 4 ) HCO 3 . Белки, связанные с шариками, ферментативно расщепляли 0,5 мкг лизил-эндопептидазы Lys-C (Wako, кат. № 125-05061) в течение 2 ч при 37°C в 3 М мочевины/50 мМ бикарбоната аммония, а затем разбавляли до 1,5 М мочевины/50 мкл. мМ бикарбонат аммония для расщепления в течение ночи с 0,8 мкг трипсина для секвенирования при 37°C.Элюирующие пептиды очищали по C18 с использованием колонок UltraMicroSpin 5–60 мкг в соответствии с инструкциями производителя и ресуспендировали в 3% ацетонитриле (ACN), 0,1% муравьиной кислоте (FA), содержащем iRT-пептиды (Biognosys AG, Шлирен, Швейцария) для масс-спектрометрического анализа.
Белки, связанные с шариками, ферментативно расщепляли 0,5 мкг лизил-эндопептидазы Lys-C (Wako, кат. № 125-05061) в течение 2 ч при 37°C в 3 М мочевины/50 мМ бикарбоната аммония, а затем разбавляли до 1,5 М мочевины/50 мкл. мМ бикарбонат аммония для расщепления в течение ночи с 0,8 мкг трипсина для секвенирования при 37°C.Элюирующие пептиды очищали по C18 с использованием колонок UltraMicroSpin 5–60 мкг в соответствии с инструкциями производителя и ресуспендировали в 3% ацетонитриле (ACN), 0,1% муравьиной кислоте (FA), содержащем iRT-пептиды (Biognosys AG, Шлирен, Швейцария) для масс-спектрометрического анализа.
Жидкостная хроматография-тандемная масс-спектрометрия (ЖХ-МС/МС) обращенно-фазовая хроматография на колонке для ВЭЖХ (внутренний диаметр 75 мкм, New Objective), которая была заполнена неподвижной фазой диаметром 15 см (ReproSil-Pur C18-AQ, 1.9 мкм) и подключен к нанопоточной ВЭЖХ с автоматическим пробоотборником (EASY-nLC 1000, Thermo Scientific). ВЭЖХ соединяли с масс-спектрометром Q-Exactive Plus (Thermo Scientific), оснащенным источником ионов с наноэлектрическим распылением (Thermo Scientific).
 Пептиды наносили на колонку со 100% буфером А (99,9% H 2 O, 0,1% FA) и элюировали при постоянной скорости потока 300 нл/мин 70-минутным линейным градиентом от 6–28% буфера B. (99,9% ACN, 0,1% FA) с последующим 4-минутным переходом от 28 до 50% буфера B.После градиента колонку промывали в течение 10 минут 98% буфером B, 4 минуты 10% буфером B и 8 минут 98% буфером B. Напряжение электрораспыления устанавливали на 2,2 кВ, а температуру капилляра — на 250 °C. Обзорный масс-спектр высокого разрешения (от 300 до 1700 m/z), полученный в Orbitrap с разрешением 70 000 при m/z 200 (целевое значение автоматической регулировки усиления 3 × 10 6 ), сопровождался спектрами МС/МС в Orbitrap с разрешением 35 000 (целевое значение автоматической регулировки усиления 1 × 10 6 ) из 12 наиболее распространенных пептидных ионов с минимальной интенсивностью 2.5 × 10 4 , которые были отобраны для фрагментации с помощью высокоэнергетической диссоциации, вызванной столкновением, с энергией столкновения 28% и окном изоляции 1,4 Да.
Пептиды наносили на колонку со 100% буфером А (99,9% H 2 O, 0,1% FA) и элюировали при постоянной скорости потока 300 нл/мин 70-минутным линейным градиентом от 6–28% буфера B. (99,9% ACN, 0,1% FA) с последующим 4-минутным переходом от 28 до 50% буфера B.После градиента колонку промывали в течение 10 минут 98% буфером B, 4 минуты 10% буфером B и 8 минут 98% буфером B. Напряжение электрораспыления устанавливали на 2,2 кВ, а температуру капилляра — на 250 °C. Обзорный масс-спектр высокого разрешения (от 300 до 1700 m/z), полученный в Orbitrap с разрешением 70 000 при m/z 200 (целевое значение автоматической регулировки усиления 3 × 10 6 ), сопровождался спектрами МС/МС в Orbitrap с разрешением 35 000 (целевое значение автоматической регулировки усиления 1 × 10 6 ) из 12 наиболее распространенных пептидных ионов с минимальной интенсивностью 2.5 × 10 4 , которые были отобраны для фрагментации с помощью высокоэнергетической диссоциации, вызванной столкновением, с энергией столкновения 28% и окном изоляции 1,4 Да. Фрагментированные предшественники динамически исключались в течение 30 с.
Фрагментированные предшественники динамически исключались в течение 30 с.
Для образцов LUX-MS под контролем бактериофагов, полученных в режиме сбора данных (DDA), пептиды разделяли с помощью обращенно-фазовой хроматографии на колонке для ВЭЖХ (внутренний диаметр 75 мкм, New Objective), которая была упакована собственными силами с стационарная фаза 15 см (ReproSil-Pur C18-AQ, 1.9 мкм) и подключен к нанопоточной ВЭЖХ с автоматическим пробоотборником (EASY-nLC 1000, Thermo Scientific). ВЭЖХ соединяли с масс-спектрометром Orbitrap Fusion Tribrid (Thermo Scientific), оснащенным источником ионов с наноэлектрическим распылением (Thermo Scientific). Пептиды наносили на колонку со 100% буфером А (99,9% H 2 O, 0,1% FA) и элюировали при постоянной скорости потока 300 нл/мин 70-минутным линейным градиентом от 6–28% буфера B. (99,9% ACN, 0,1% FA) с последующим 4-минутным переходом от 28 до 50% буфера B.После градиента колонку промывали в течение 10 мин 98%, 4 мин 10% и снова 8 мин 98% буфером B. Напряжение электрораспыления устанавливали на 1,8 кВ, а температуру капилляра на 275. Масс-спектр высокого разрешения. (от 395 до 1500 м/z), полученных в Orbitrap с разрешением 120 000 при m/z 200 (целевое значение автоматической регулировки усиления 2 × 10 5 и максимальное время ввода 50 мс), сопровождались спектрами МС/МС большинства -обильные пептидные ионы с минимальной интенсивностью 5 × 10 3 , которые были отобраны для последующей высокоэнергетической диссоциационной фрагментации, вызванной столкновением, с энергией столкновения 35% и окном изоляции 1.6 Да. Фрагменты были обнаружены с помощью МС/МС в ионной ловушке со скоростью сканирования, установленной на «Быстрое» (целевое значение автоматической регулировки усиления 1 × 10 4 и максимальное время ввода 250 мс). Фрагментированные предшественники динамически исключались в течение 30 с.
Напряжение электрораспыления устанавливали на 1,8 кВ, а температуру капилляра на 275. Масс-спектр высокого разрешения. (от 395 до 1500 м/z), полученных в Orbitrap с разрешением 120 000 при m/z 200 (целевое значение автоматической регулировки усиления 2 × 10 5 и максимальное время ввода 50 мс), сопровождались спектрами МС/МС большинства -обильные пептидные ионы с минимальной интенсивностью 5 × 10 3 , которые были отобраны для последующей высокоэнергетической диссоциационной фрагментации, вызванной столкновением, с энергией столкновения 35% и окном изоляции 1.6 Да. Фрагменты были обнаружены с помощью МС/МС в ионной ловушке со скоростью сканирования, установленной на «Быстрое» (целевое значение автоматической регулировки усиления 1 × 10 4 и максимальное время ввода 250 мс). Фрагментированные предшественники динамически исключались в течение 30 с.
Для образцов LUX-MS с контролем антител, полученных в режиме независимого сбора данных (DIA), пептиды разделяли с помощью обращенно-фазовой хроматографии на колонке для ВЭЖХ (внутренний диаметр 75 мкм, New Objective), которая была упакована собственными силами стационарная фаза 15 см (ReproSil-Pur C18-AQ, 1. 9 мкм) и подключен к нанопоточной ВЭЖХ с автоматическим пробоотборником (EASY-nLC 1200, Thermo Scientific). ВЭЖХ соединяли с масс-спектрометром Orbitrap Fusion Tribrid (Thermo Scientific), оснащенным источником ионов с наноэлектрическим распылением (Thermo Scientific). Пептиды наносили на колонку со 100% буфером А (99,9% H 2 O, 0,1% FA) и элюировали при постоянной скорости потока 250 нл/мин с 70-минутным линейным градиентом от 6–28% буфера B. (99,9% ACN, 0,1% FA) с последующим 4-минутным переходом от 28 до 50% буфера B.После градиента колонку промывали в течение 10 минут 98% буфером B, 4 минуты 10% буфером B и 8 минут 98% буфером B. Напряжение электрораспыления устанавливали на 4,0 кВ, а температуру капилляра — на 320 °C. Обзорное сканирование, полученное на орбитальной ловушке с разрешением 120 000, максимальным временем инжекции 50 мс и целью АРУ 3×10 6 , сопровождалось 24 предварительными окнами с фрагментацией при энергии столкновения 28% и получением спектров МС/МС в орбитальная ловушка с разрешением 30 000.
9 мкм) и подключен к нанопоточной ВЭЖХ с автоматическим пробоотборником (EASY-nLC 1200, Thermo Scientific). ВЭЖХ соединяли с масс-спектрометром Orbitrap Fusion Tribrid (Thermo Scientific), оснащенным источником ионов с наноэлектрическим распылением (Thermo Scientific). Пептиды наносили на колонку со 100% буфером А (99,9% H 2 O, 0,1% FA) и элюировали при постоянной скорости потока 250 нл/мин с 70-минутным линейным градиентом от 6–28% буфера B. (99,9% ACN, 0,1% FA) с последующим 4-минутным переходом от 28 до 50% буфера B.После градиента колонку промывали в течение 10 минут 98% буфером B, 4 минуты 10% буфером B и 8 минут 98% буфером B. Напряжение электрораспыления устанавливали на 4,0 кВ, а температуру капилляра — на 320 °C. Обзорное сканирование, полученное на орбитальной ловушке с разрешением 120 000, максимальным временем инжекции 50 мс и целью АРУ 3×10 6 , сопровождалось 24 предварительными окнами с фрагментацией при энергии столкновения 28% и получением спектров МС/МС в орбитальная ловушка с разрешением 30 000. Время впрыска было установлено на автоматический режим, цель АРУ на 1×10 6 и диапазон масс на 300-1700 м/з.Для спектральной библиотеки образцы были дополнительно измерены на платформе Q-Exactive Plus в режиме сбора данных (DDA), как описано выше.
Время впрыска было установлено на автоматический режим, цель АРУ на 1×10 6 и диапазон масс на 300-1700 м/з.Для спектральной библиотеки образцы были дополнительно измерены на платформе Q-Exactive Plus в режиме сбора данных (DDA), как описано выше.
пептиды разделяли с помощью обращенно-фазовой хроматографии на колонке для ВЭЖХ (внутренний диаметр 75 мкм, New Objective), которая была заполнена 30-см неподвижной фазой (ReproSil-Pur C18-AQ, 1,9 мкм) и соединена с нанопоточная ВЭЖХ с автоматическим пробоотборником (EASY-nLC 1200, Thermo Scientific).ВЭЖХ соединяли с масс-спектрометром Orbitrap Fusion Lumos Tribrid (Thermo Scientific), оснащенным источником ионов с наноэлектрическим распылением (Thermo Scientific). Пептиды наносили на колонку со 100% буфером А (99,9% H 2 O, 0,1% FA) и элюировали при постоянной скорости потока 250 нл/мин с 240-минутным нелинейным градиентом от 4 до 58% буфер B (80% ACN, 0,1% FA). После градиента колонку промывали в течение 10 минут 98% буфером B, 4 минуты 1% буфером B. Напряжение электрораспыления устанавливали на 3.6 кВ и капиллярной температуре до 320 °С. Обзорное сканирование, полученное на орбитальной ловушке с разрешением 120 000, максимальным временем инжекции 50 мс и целью АРУ 4×10 5 , сопровождалось 80 предварительными окнами с фрагментацией при энергии столкновения 27% и получением спектров МС/МС в орбитальная ловушка с разрешением 30 000. Время впрыска было установлено на автоматический режим, цель АРУ на 1×10 6 и диапазон масс на 350-2000 м/з. Для спектральной библиотеки повторы освещенного и неосвещенного состояния были объединены и проанализированы на одной и той же приборной установке в режиме сбора данных (DDA) с использованием универсального метода с одинаковым градиентом и энергией столкновения 27% и 30%.
Напряжение электрораспыления устанавливали на 3.6 кВ и капиллярной температуре до 320 °С. Обзорное сканирование, полученное на орбитальной ловушке с разрешением 120 000, максимальным временем инжекции 50 мс и целью АРУ 4×10 5 , сопровождалось 80 предварительными окнами с фрагментацией при энергии столкновения 27% и получением спектров МС/МС в орбитальная ловушка с разрешением 30 000. Время впрыска было установлено на автоматический режим, цель АРУ на 1×10 6 и диапазон масс на 350-2000 м/з. Для спектральной библиотеки повторы освещенного и неосвещенного состояния были объединены и проанализированы на одной и той же приборной установке в режиме сбора данных (DDA) с использованием универсального метода с одинаковым градиентом и энергией столкновения 27% и 30%.
Влияние фотопериода и спектров света на рост и пигментный состав зеленой макроводоросли Codium tomentosum
Abreu MH, Pereira R, Sousa-Pinto I, Yarish C (2011) Экофизиологические исследования неаборигенных видов Gracilaria vermiculophylla (Rhodophyta) и закономерности их численности в лагуне Риа-де-Авейру, Португалия.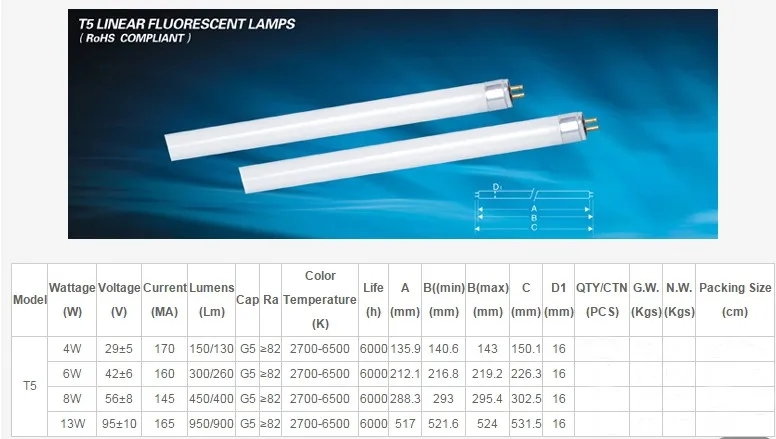 Eur J Phycol 46:453–464
Eur J Phycol 46:453–464
Статья
Google Scholar
Abreu MH, Pereira R, Sassi J-F (2014) Морские водоросли и глобальная пищевая промышленность.В: Перейра Л., Нето Дж. М. (ред.) Морские водоросли — биоразнообразие, таксономия, оценка окружающей среды и биотехнология. CRC Press, Бока-Ратон, стр. 300–319
Google Scholar
Акимото С., Томо Т., Найтох Ю., Отомо А., Мураками А., Мимуро М. (2007) Идентификация нового возбужденного состояния, ответственного за уникальную полосу поглощения сифонаксантина in vivo в зеленых водорослях Кодиум хрупкий . J Phys Chem B 111:9179–9181
CAS
пабмед
Статья
Google Scholar
Anderson JM (1983) Хлорофилл-белковые комплексы видов Codium , включая светособирающий комплекс a/b-белков сифонаксантин-хлорофилл, эволюционный реликт некоторых Chlorophyta. Биохим Биофиз Акта 724:370–380
Биохим Биофиз Акта 724:370–380
CAS
Статья
Google Scholar
Anderson JM, Chow WS, Goodchild DJ (1988) Организация тилакоидной мембраны при акклиматизации к солнцу/тени. Aust J Plant Physiol 15:11–26
Google Scholar
Benson EE, Cobb AH (1983) Пигментно-белковые комплексы литоральной водоросли Codium хрупкий (Suringar) Hariot. Новый фитол 95: 581–594
CAS
Статья
Google Scholar
Benson EE, Cobb AH (1981) Разделение, идентификация и количественное определение фотопигментов из сифоновых морских водорослей Codium хрупкий.Новый фитол 88: 627–632
CAS
Статья
Google Scholar
Бунгард Р.А., Рубан А.В., Хибберд Дж.М., Пресс М.С., Хортон П., Шоулз Дж.Д. (1999) Необычный состав каротиноидов и новый тип ксантофиллового цикла в растениях. Proc Nat Acad Sci USA 96:1135–1139
Proc Nat Acad Sci USA 96:1135–1139
CAS
Статья
Google Scholar
Cartaxana P, Morelli L, Quintaneiro C, Calado G, Calado R, Cruz S (2018)Состояние фотоакклиматизации клептопластов модулирует фотоповедение морского слизня, работающего на солнечной энергии Elysia viridis .J Exp Biol 221:jeb180463
PubMed
Статья
Google Scholar
Christa G, Cruz S, Jahns P, de Vries J, Cartaxana P, Esteves AC, Serôdio J, Gould SB (2017)Фотозащита в монофилетической ветви хлорофитных водорослей не зависит от энергозависимого тушения (qE). Новый фитол 214:1132–1144
CAS
пабмед
Статья
ПабМед Центральный
Google Scholar
Круз С., Картаксана П., Новичок Р., Дионисио Г., Каладо Р., Серодио Дж., Пеллетро К.Н., Румфо М.Е. (2015) Фотозащита в изолированных пластидах морских слизней и соответствующих источниках водорослей. Научный представитель 5:7904
Научный представитель 5:7904
CAS
пабмед
ПабМед Центральный
Статья
Google Scholar
Costa E, Melo T, Moreira ASP, Alves E, Domingues P, Calado R, Abreu MH, Domingues MR (2015) Расшифровка профиля биоактивных полярных липидов макроводорослей Codium tomentosum из устойчивой системы IMTA с использованием липидомного анализа подход. Algal Res 12:388–397
Статья
Google Scholar
Figueroa FL, Aguilera J, Niell X (1995) Регуляция роста и фотосинтетического метаболизма красным и синим светом у Porphyra umbilicalis (Bangiales, Rhodophyta).Eur J Phycol 30:11–18
Статья
Google Scholar
Франклин Л.А., Ларкум А.В.Д. (1997) Множественные стратегии существования тропических морских макроводорослей при ярком свете. Photosynth Res 53:149–159
CAS
Статья
Google Scholar
Ганесан П. , Мацубара К., Окубо Т., Танака Й., Нодаа К., Сугавараа Т., Хирата Т. (2010) Антиангиогенный эффект сифонаксантина из зеленых водорослей, Кодиум хрупкий .Фитомедицина 17:1140–1144
, Мацубара К., Окубо Т., Танака Й., Нодаа К., Сугавараа Т., Хирата Т. (2010) Антиангиогенный эффект сифонаксантина из зеленых водорослей, Кодиум хрупкий .Фитомедицина 17:1140–1144
CAS
пабмед
Статья
ПабМед Центральный
Google Scholar
Ganesan P, Noda K, Manabe Y, Ohkubo T, Tanaka Y, Maoka T, Sugawaraa T, Hirata T (2011) Сифонаксантин, морской каротиноид из зеленых водорослей, эффективно индуцирует апоптоз при лейкемии человека (HL-60) клетки. Биохим Биофиз Акта 1810:497–503
CAS
пабмед
Статья
ПабМед Центральный
Google Scholar
Джовагнетти В., Хан Г., Уэр М.А., Унгерер П., Цинь С., Ван В.Д., Куанг Т., Шен Дж.Р., Рубан А.В. (2018) Сифонная морфология влияет на светособирающую модуляцию у литоральных зеленых макроводорослей Bryopsis corticulans (ульвофитовые).Планта 247:1293–1306
CAS
пабмед
ПабМед Центральный
Статья
Google Scholar
Genty B, Briantais J-M, Baker NR (1989) Взаимосвязь между квантовым выходом фотосинтетического переноса электронов и гашением флуоресценции хлорофилла.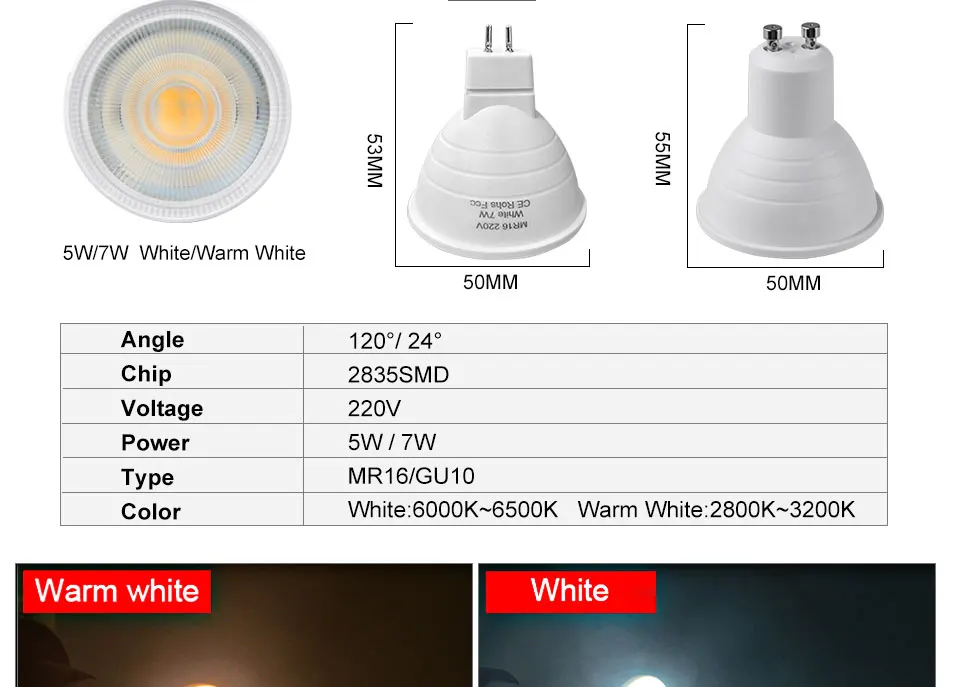 Биохим Биофиз Акта 990:87–92
Биохим Биофиз Акта 990:87–92
CAS
Статья
Google Scholar
Гранада Л., Соуза Н., Лопес С., Лемос М.Ф. (2016) Является ли интегрированная мультитрофическая аквакультура решением основных проблем отрасли? — Обзор.Rev Aquac 8:283–300
Статья
Google Scholar
Госс Р., Лепетит Б. (2015) Биоразнообразие NPQ. J Plant Physiol 172:13–32
CAS
пабмед
Статья
Google Scholar
Hanisak MD (1979) Модели роста Codium хрупкий подвид. tomentosoides в зависимости от температуры, освещенности, солености и источника азота. Мар Биол 50:319–332
CAS
Статья
Google Scholar
Hwang EK, Baek JM, Parket CS (2008) Выращивание зеленых водорослей, Кодиум хрупкий (Suringar) Hariot, выращивание искусственных семян в Корее. J Appl Phycol 20:469–475
J Appl Phycol 20:469–475
Статья
Google Scholar
Jahns P, Holzwarth AR (2012)Роль ксантофиллового цикла и лютеина в фотозащите фотосистемы II. Biochim Biophys Acta Bioenerg 1817:182–193
CAS
Статья
Google Scholar
Джеффри С. (1980) Пигментные системы водорослей. В: Фальковский П.Г. (ред.) Первичная продуктивность в море.Спрингер, Бостон, стр. 33–58
Глава
Google Scholar
Юнгандеас А., Коста Б.С., Якоб Т., фон Берген М., Бауманн С., Вильгельм С. (2014) Акклиматизация Phaeodactylum tricornutum к синему и красному свету не влияет на реакцию фотосинтеза, но сильно нарушает выделение углерода шаблон. PLoS One 9:e99727
PubMed
ПабМед Центральный
Статья
КАС
Google Scholar
Kraay GW, Zapata M, Veldhuis MJW (1992) Разделение хлорофиллов c 1 , c 2 и c 3 морского фитопланктона с помощью обращенно-фазовой C18-высокоэффективной жидкостной хроматографии. J Phycol 28:708–712
J Phycol 28:708–712
CAS
Статья
Google Scholar
Kirk JTO (2011) Свет и фотосинтез в водных экосистемах. Издательство Кембриджского университета, Кембридж
Google Scholar
Лоббан К.С., Харрисон П.Дж. (1994) Экология и физиология морских водорослей. Издательство Кембриджского университета, Кембридж
Книга
Google Scholar
Люнинг К. (1990) Морские водоросли – среда их обитания, биогеография и экофизиология.Джон Вили и сыновья, Нью-Йорк
Google Scholar
Мацубара К., Мацуура Ю., Хори К., Миядзава К. (2000) Протеогликан-антикоагулянт из морских зеленых водорослей, Codium pugniformis . J Appl Phycol 12:9–14
CAS
Статья
Google Scholar
Mendes CR, Cartaxana P, Brotas V (2007) Определение пигментов фитопланктона и микрофитобентоса методом ВЭЖХ: сравнение разрешения и чувствительности методов C18 и C8.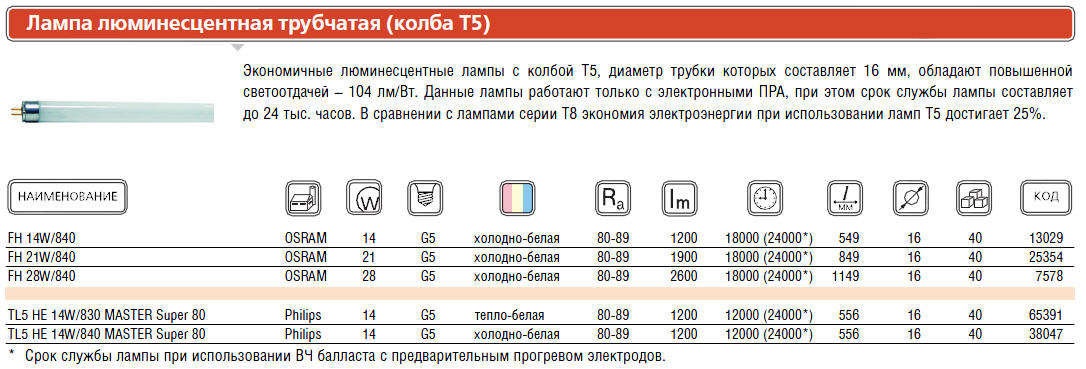 Limnol Oceanogr Methods 5:363–370
Limnol Oceanogr Methods 5:363–370
CAS
Статья
Google Scholar
Murchie EH, Lawson T (2013) Анализ флуоресценции хлорофилла: руководство по передовой практике и понимание некоторых новых приложений. J Exp Bot 64: 3983–3998
CAS
Статья
пабмед
Google Scholar
Nanba N, Kado R, Ogawa H, Nakagawa T, Sugiura Y (2005) Влияние освещенности и потока воды на формирование и рост губчатых и нитевидных талломов Codium хрупкого .Aquat Bot 81:315–325
Артикул
Google Scholar
Ohta Y, Lee J-B, Hayashi K, Hayashi T (2009) Выделение сульфатированного галактана из Codium хрупкого и его противовирусный эффект. Биол Фарм Булл 32:892–898
CAS
пабмед
Статья
ПабМед Центральный
Google Scholar
Pereira R, Sousa-Pinto I, Yarish C (2004) Полевые и культуральные исследования истории жизни Porphyra dioica (Bangiales, Rhodophyta) из Португалии. Phycologia 43:756–767
Phycologia 43:756–767
Статья
Google Scholar
Перейра Л. (2018) Морские водоросли как источник биоактивных веществ и уход за кожей – космецевтика, альготерапия и талассотерапия. Косметика 5:68
CAS
Статья
Google Scholar
Перес-Льоренс Х., Эрнандес I, Вергара Х. (2018) Эти любопытные и вкусные водоросли. Увлекательное путешествие от биологии к гастрономии.UCA Press, Кадис
Pillay TVR (2004) Аквакультура и окружающая среда. Издательство Блэквелл, Оксфорд
Книга
Google Scholar
Platt T, Gallegos CL, Harrison WG (1980) Фотоингибирование фотосинтеза в естественных сообществах морского фитопланктона. Дж. Мар Рес 38: 687–701
Google Scholar
Радулович Р., Уманзор С., Кабрера Р., Мата Р. (2015) Тропические водоросли в пищу человеку, их выращивание и влияние на обогащение биоразнообразия. Аквакультура 436:40–46
Аквакультура 436:40–46
Статья
Google Scholar
Rey F, Cartaxana P, Melo T, Calado R, Pereira R, Abreu H, Domingues P, Cruz S, Domingues MR (2020) Домашние популяции Codium tomentosum демонстрируют экстракты липидов с более низкими сезонными сдвигами, чем сородичи из дикий — актуальность для биотехнологических применений этой зеленой водоросли. Mar Drugs 18:188
CAS
ПабМед Центральный
Статья
пабмед
Google Scholar
Самаравира А.М., Виданараччи Дж.К., Курукуласурия М.С. (2011) Промышленное применение макроводорослей.В: Ким С.К. (ред.) Справочник по морским макроводорослям: биотехнология и прикладная психология. John Wiley & Sons, Чичестер, стр. 500–521
Глава
Google Scholar
Santos SAO, Vilela C, Freire CSR, Abreu MH, Rocha SM, Silvestre AJD (2015) Макроводоросли Chlorophyta и Rhodophyta: источник полезных для здоровья фитохимических веществ. Food Chem 183:122–128
Food Chem 183:122–128
CAS
пабмед
Статья
Google Scholar
Senge M, Senger H (1991) Адаптация фотосинтетического аппарата Chlorella и Ankistrodesmus к синему и красному свету.Бот Acta 104:139–143
CAS
Статья
Google Scholar
Троелл М., Холлинг С., Неори А., Шопен Т., Бушманн А.Х., Каутский Н., Яриш С. (2003) Интегрированная марикультура: задавайте правильные вопросы. Аквакультура 226:69–80
Статья
Google Scholar
Урагами С., Галзерано Д., Галл А., Шигемацу Ю., Мейстерханс М., Ока Н., Иха М., Фуджи Р., Роберт Б., Хасимото Х. (2014) Светозависимое конформационное изменение неоксантина в сифонной зеленой водоросли, Codium intricatum , выявленный с помощью рамановской спектроскопии.Photosynth Res 121:69–77
CAS
пабмед
Статья
Google Scholar
Wang H-MD, Chen C-C, Huynh P, Chang J-S (2015) Изучение потенциала использования водорослей в косметике. Биоресурс Технол 184:355–362
Биоресурс Технол 184:355–362
CAS
пабмед
Статья
Google Scholar
Wang L, Wang X, Wu H, Liu R (2014) Обзор биологической активности и молекулярных характеристик сульфатированных полисахаридов морских зеленых водорослей за последние годы.Mar Drugs 12:4984–5020
CAS
пабмед
ПабМед Центральный
Статья
Google Scholar
Wang W, Qin X, Sang M, Chen D, Wang K, Lin R, Lu C, Shen JR, Kuang T (2013) Спектральные и функциональные исследования светособирающего комплекса сифонаксантинового типа фотосистемы II из Бриопсис кортикальный . Photosynth Res 117: 267–279
CAS
пабмед
Статья
Google Scholar
Yang MH, Blunden G, Huang FL, Fletcher RL (1997) Рост диссоциированной нитевидной стадии видов Codium в лабораторной культуре.J Appl Phycol 9:1–3
CAS
Статья
Google Scholar
Транслатом тел нейронов, дендритов и аксонов
Значение
Белки являются ключевыми факторами синаптической функции нейронов. Регуляция экспрессии генов важна для формирования и модификации синапсов на протяжении всей жизни. Сложность дендритов и аксонов создает уникальные проблемы для снабжения белком в удаленных местах.Открытие матричных РНК (мРНК) и рибосом вблизи синапсов показало, что локальный синтез белка представляет собой важное решение этой проблемы. Здесь мы использовали секвенирование РНК и секвенирование рибосом для непосредственного определения популяции мРНК, которая присутствует и находится в процессе трансляции в телах нейронов, дендритах и аксонах. Тысячи транскриптов были по-разному транслированы между телом клетки и синаптическими областями, причем более 800 мРНК демонстрировали большую трансляцию в дендритно-аксональном компартменте.
Регуляция экспрессии генов важна для формирования и модификации синапсов на протяжении всей жизни. Сложность дендритов и аксонов создает уникальные проблемы для снабжения белком в удаленных местах.Открытие матричных РНК (мРНК) и рибосом вблизи синапсов показало, что локальный синтез белка представляет собой важное решение этой проблемы. Здесь мы использовали секвенирование РНК и секвенирование рибосом для непосредственного определения популяции мРНК, которая присутствует и находится в процессе трансляции в телах нейронов, дендритах и аксонах. Тысячи транскриптов были по-разному транслированы между телом клетки и синаптическими областями, причем более 800 мРНК демонстрировали большую трансляцию в дендритно-аксональном компартменте.
Abstract
Для формирования синаптических связей и хранения информации нейроны постоянно реконструируют свои протеомы. Впечатляющая длина дендритов и аксонов создает логистические проблемы для поддержания синаптических белков в местах, удаленных от источника транскрипции (ядра). Открытие тысяч матричных РНК (мРНК) вблизи синапсов показало, что нейроны преодолевают расстояние и обретают автономию, производя белки локально. Однако в целом не известно, транслируются ли, как и когда локализованные мРНК в белок.Чтобы исследовать ландшафт трансляции в субрегионах нейронов, мы провели одновременное секвенирование РНК (RNA-seq) и секвенирование рибосом (Ribo-seq) из микрорассеченных срезов мозга грызунов, чтобы идентифицировать и количественно определить транскриптом и транслатом в клеточных телах (соматах), а также в дендритах. и аксоны (нейропиль). Тысячи транскриптов по-разному транслировались между соматическими и синаптическими областями, при этом многие каркасные и сигнальные молекулы демонстрировали повышенные уровни трансляции в нейропиле.Большинство трансляционных изменений между компартментами можно объяснить различиями в изобилии РНК. Проникающая трансляционная регуляция наблюдалась как в соматах, так и в нейропиле под влиянием специфических особенностей мРНК (например, длины нетранслируемой области [UTR], мотивов РНК-связывающего белка [RBP] и открытых рамок считывания выше по течению [uORFs]).
Открытие тысяч матричных РНК (мРНК) вблизи синапсов показало, что нейроны преодолевают расстояние и обретают автономию, производя белки локально. Однако в целом не известно, транслируются ли, как и когда локализованные мРНК в белок.Чтобы исследовать ландшафт трансляции в субрегионах нейронов, мы провели одновременное секвенирование РНК (RNA-seq) и секвенирование рибосом (Ribo-seq) из микрорассеченных срезов мозга грызунов, чтобы идентифицировать и количественно определить транскриптом и транслатом в клеточных телах (соматах), а также в дендритах. и аксоны (нейропиль). Тысячи транскриптов по-разному транслировались между соматическими и синаптическими областями, при этом многие каркасные и сигнальные молекулы демонстрировали повышенные уровни трансляции в нейропиле.Большинство трансляционных изменений между компартментами можно объяснить различиями в изобилии РНК. Проникающая трансляционная регуляция наблюдалась как в соматах, так и в нейропиле под влиянием специфических особенностей мРНК (например, длины нетранслируемой области [UTR], мотивов РНК-связывающего белка [RBP] и открытых рамок считывания выше по течению [uORFs]). Для более чем 800 мРНК доминирующим источником трансляции был нейропиль. Мы создали интерактивную базу данных с возможностью поиска для изучения транскриптов мРНК и уровней их трансляции в сомате и нейропиле [MPI Brain Research, The mRNA Translation landscape in the synaptic neuropil.https://public.brain.mpg.de/dashapps/localseq/. По состоянию на 5 октября 2021 г.]. В целом, наши результаты подчеркивают существенный вклад локальной трансляции в поддержание уровней синаптических белков и указывают на то, что локальный контроль трансляции является важным механизмом контроля синаптической силы.
Для более чем 800 мРНК доминирующим источником трансляции был нейропиль. Мы создали интерактивную базу данных с возможностью поиска для изучения транскриптов мРНК и уровней их трансляции в сомате и нейропиле [MPI Brain Research, The mRNA Translation landscape in the synaptic neuropil.https://public.brain.mpg.de/dashapps/localseq/. По состоянию на 5 октября 2021 г.]. В целом, наши результаты подчеркивают существенный вклад локальной трансляции в поддержание уровней синаптических белков и указывают на то, что локальный контроль трансляции является важным механизмом контроля синаптической силы.
В синапсах нейронов более 2500 белков (1, 2) («синаптический протеом») действуют как сенсоры и эффекторы, контролирующие возбудимость нейронов, синаптическую силу и пластичность. Сложная морфология и функциональная компартментализация отдельного нейрона создает уникальные логистические проблемы для поддержания и модификации синаптического протеома в местах, удаленных от источника транскрипции (т.д., ядро). Чтобы удовлетворить локальный спрос на новый белок, нейроны локализуют матричные РНК (мРНК) и рибосомы вблизи синапсов, чтобы производить белки непосредственно там, где они необходимы (1). Используя высокопроизводительное секвенирование, несколько групп сообщили о локализации тысяч транскриптов в аксонах и дендритах («локальный транскриптом») (3⇓⇓⇓–7). Однако во многих типах клеток было показано, что уровни транскриптов не всегда предсказывают уровни белка (8), что позволяет предположить, что трансляция мРНК является строго регулируемым процессом.Поскольку белки, а не мРНК, управляют клеточной функцией, необходимо непосредственно определить, какие транскрипты транслируются в белки в дендритах и/или аксонах in vivo («локальный транслатом»). Важно, что остается неизвестным, какие транскрипты обнаруживают разные уровни трансляции между соматическими и синаптическими областями.
Уровень трансляции данного транскрипта определяется скоростью набора рибосомы к стартовому кодону во время инициации и скоростью транслокации рибосомы во время удлинения полипептида.Для большинства мРНК инициация трансляции считается ограничивающей скорость (9): инициация регулируется элементами внутри нетранслируемых областей мРНК (UTR), которые связывают РНК-связывающие белки (RBP) или микроРНК (10⇓–12). Кроме того, скорость элонгации также играет регулирующую роль в определении количества белка, продуцируемого из транскрипта (13). Хотя нарушенный трансляционный контроль связан с рядом неврологических расстройств (14), мало что известно о величине и механизмах специфичной для транскрипта регуляции трансляции в нейрональных компартментах.
В этом исследовании мы объединили глубокое секвенирование защищенных рибосомами фрагментов (рибосомное секвенирование [Ribo-seq]) и секвенирование РНК (RNA-seq) микрорассеченных срезов мозга грызунов, чтобы обеспечить всесторонний анализ трансляционного ландшафта мРНК как в соматы (обогащенные телами клеток) и нейропиль (обогащенные дендритами/аксонами нейронов). Тысячи мРНК были транслированы в соматических и синаптических областях. Многие транскрипты обнаруживают различные уровни трансляции между соматическими и синаптическими областями.Многие из этих трансляционных изменений, вероятно, являются результатом различий в уровнях РНК между соматами и нейропилем. Кроме того, мы нашли доказательства всепроникающей трансляционной регуляции синаптических белков в обоих отделах нейронов. Мы предоставляем динамический веб-интерфейс на основе запросов для изучения транскриптов мРНК и их трансляции в нейронных компартментах (15). Вместе наши результаты показывают беспрецедентную способность локального производства белка in vivo для поддержания и модификации пре- и постсинаптического протеома.
Результаты
Измерение транскриптомной трансляции в нейронных компартментах.
Чтобы обнаружить виды мРНК, локализованные и транслируемые в телах клеток, а также в дендритах и аксонах, мы провели полногеномный анализ транскриптома и транслатома соматы и нейропиля из микрорассеченных срезов гиппокампа (16). Следы рибосом были получены из лизатов сомат и нейропилей для оценки количества и положения транслирующих рибосом на транскрипте (Ribo-seq) (17).Параллельно с этим определяли уровни транскриптов, выполняя секвенирование РНК из сомата и нейропиля (рис. 1 –) (16). Библиотеки РНК и рибо-seq из соматы и нейропиля были высоко воспроизводимы среди трех биологических повторов ( SI, Приложение , рис. S1 A и B ). Кроме того, образцы Ribo-seq продемонстрировали ожидаемое снижение плотности считывания следа в UTR и интронах транскриптов ( SI, Приложение , рис. S1, C и D ), а также трехнуклеотидную фазировку ( SI). Приложение , рис.S1 E и F ) (17).
Рис. 1.
Многие транскрипты демонстрируют дифференциальную трансляцию между соматой и нейропилем. ( A ) Экспериментальный рабочий процесс. Микродиссекция области СА1 гиппокампа крысы. РНК-секвенирование и рибо-секвенирование проводили одновременно для слоя сомата (обогащенного телами пирамидных нейронов) и нейропиля (обогащенного дендритами и аксонами). Нейронный фильтр был применен для обогащения транскриптов возбуждающих нейронов в последующих анализах.( B ) График вулкана, сравнивающий уровень трансляции 7850 транскриптов между компартментами (соотношение нейропил: сомата Ribo-seq [log 2 FC]). FDR < 0,05 с использованием DESeq2 ( Экспериментальные процедуры ). Цветными точками выделены транскрипты, значительно более транслируемые в сомате (сомата [smt]-translation-up, n = 2,945, оранжевый) или нейропиле (neuropil [npl]-translation-up, n = 807, бирюзовый). ( C ) Треки покрытия, представляющие среднее нейропилевое ( Верхнее ) или соматное ( Нижнее ) покрытие рибосомного следа кандидата smt-translation-up ( Gria2 , Neurod6 и Hpca ) перевод вверх ( Shank1 , Map2 , и Dgkz ) стенограммы.Ось y указывает количество нормализованных прочтений. ( D ) Схема, изображающая отток рибосом in vivo после инкубации с харрингтонином культур крысиного гиппокампа. ( E ) Коэффициенты элонгации для smt-translation-up (оранжевый), npl-translation-up (бирюзовый) и других (серый) транскриптов, выведенные из наклона линейной аппроксимации, показанной в SI Приложение , рис. S4 нанесены с их SE ( n = 3). P = 0,5738, однофакторный дисперсионный анализ. Хар, харрингтонин; Chx, циклогексимид; нс, не имеет значения.
Мы обнаружили 13 055 и 12 371 транскриптов с числом на миллион (CPM) в двух из трех нейропилей ( SI Приложение , рис. S2 A ) или соматах ( SI Приложение , рис. S2 B ) Рибо -seq повторяется соответственно. Используя наборы данных Ribo-seq, мы обнаружили существенное совпадение между нашими данными транслатома и ранее опубликованным транскриптомом нейропиля ( SI Приложение , рис. S2 A ) и соматы ( SI Приложение , рис. S2 B ) ( 3).Соматы и нейропиль гиппокампа содержат тела возбуждающих нейронов и их отростки, а также глию и интернейроны. Мы создали конвейер, чтобы сосредоточиться на генах возбуждающих нейронов, сводя к минимуму вклад других типов клеток с помощью биоинформационной фильтрации. Чтобы получить полный набор транскриптов, обогащенных глией, мы подготовили культуры нейронов и глии гиппокампа (приложение SI, рис. S2 C и набор данных S1). Поскольку сомата и нейропиль содержат не только глию, но и интернейроны, мы дополнительно составили списки транскриптов, обогащенных невозбуждающими типами нейронов в гиппокампе.Для этого мы идентифицировали значительно деобогащенные транскрипты в гиппокампе двух разных линий мышей RiboTag, которые нацелены в первую очередь на возбуждающие нейроны: мыши Camk2Cre::RiboTag ( SI, Приложение , рис. S2 D ), а также микрорассеченные соматы. ( SI Приложение , рис. S2 E ) и нейропил ( SI Приложение , рис. S2 F ) от мышей Wfs1Cre::RiboTag (16). Объединив эти наборы данных, мы получили список «загрязненных» генов невозбуждающих нейронов ( Приложение SI , рис.S2 G ).
Во многих расшифровках показана дифференциальная трансляция между нейронными компартментами.
Количество рибосом, загруженных в транскрипт, указывает, насколько он транслируется. Чтобы идентифицировать транскрипты, которые демонстрируют дифференциальную трансляцию между соматой и нейропилем, мы рассчитали соотношение нейропиль: сомата Ribo-seq (DESeq2) (18) ( Experimental Procedures ). После вычитания генов-загрязнителей мы обнаружили 7850 нейронных транскриптов ( Приложение SI , рис.S2 H ) (19), которые транслировались как в сомате, так и в нейропиле (рис. 1 B ). Из них 807 транскриптов демонстрировали значительно повышенные уровни трансляции в нейропиле по сравнению с соматами («нейропиль-трансляция вверх») (рис. 1 B и набор данных S2). Транскрипты-трансляции нейропиля включали, например, Shank1 , Map2 и Dgkz (рис. 1 B и C ). Напротив, 2945 транскриптов показали повышенную трансляцию в соматах, в том числе Gria2 , Neurod6 и Hpca («somata-translation-up») (рис.1 B и C и набор данных S2). Транскрипты как нейропиля, так и сомата-трансляции демонстрируют трехнуклеотидную периодичность, возникающую в результате покодонной транслокации рибосом вдоль мРНК во время трансляции в нейропиле и сомате, соответственно ( SI Приложение , рис. S3 A и Б ). В соответствии с предыдущими выводами (12), транскрипты нейропиля с трансляцией вверх демонстрировали значительно более длинные 3′-UTR ( SI Приложение , рис.S3 С ).
Предыдущие исследования предполагали, что мРНК, присутствующие в дендритах и/или аксонах, могут трансляционно молчать посредством «паузы» рибосом на уровне элонгации (13, 20). Чтобы решить эту проблему, мы спросили, демонстрируют ли транскрипты с трансляцией нейропиля и соматы различия в скорости элонгации трансляции. Мы выполнили временной ряд «оттока» рибосом, инкубируя культивируемые нейроны гиппокампа в течение 15, 30, 45 или 90 с с харрингтонином, препаратом, который иммобилизует рибосомы сразу после инициации трансляции, что приводит к прогрессивному оттоку рибосом в течение время (рис.1 D и SI Приложение , рис. S4). Мы проанализировали скорость рибосомной прогрессии (элонгации) от 5′-конца транскриптов нейропиля и сомата-трансляции вверх ( SI Приложение , рис. S4). Подмножества транскриптов нейропиля и сомата-трансляции показали одинаковую скорость элонгации ~4 кодона в секунду (рис. 1 E и SI Приложение , рис. S4), значение, которое находится в пределах диапазона, измеренного в других типы клеток (от 3 до 10 кодонов в секунду) (21⇓⇓–24).В совокупности эти находки указывают на то, что мРНК, повышающие трансляцию нейропиля, глобально не значительно более приостановлены, чем др. транскрипты.
Чтобы выяснить, кодируются ли определенные функциональные группы белков транскриптами, демонстрирующими повышенные уровни трансляции в любом из компартментов, мы провели анализ генной онтологии (GO) (рис. 2 A и B ). Обогащение терминов, связанных с синаптической функцией, было обнаружено как для сомата-транскриптов, так и для транскриптов нейропиля (рис.2 A и B ). Для транскриптов сомата-трансляции мы наблюдали значительное преобладание термина «перикарион», а также многих терминов, связанных с мембранами, таких как «интегральный компонент постсинаптической мембраны плотности», «пресинаптическая мембрана» или «мембрана синаптических пузырьков». (Рис. 2 A ). С другой стороны, в основном постсинаптические функции были в значительной степени связаны с транскриптами нейропиля-трансляции-вверх, включая, например, «дендритный шип» и «постсинаптическую плотность» (рис.2 В ). Чтобы лучше понять синаптическую функцию транскриптов нейропиля и сомата-трансляции, мы проанализировали кратность нейропиля: сомата Ribo-seq изменений возбуждающих синаптических белков (рис. 2 C ). Мы отметили, что субъединицы ионотропных и метаботропных глутаматных рецепторов (AMPAR, NMDAR и mGluR) в основном демонстрировали более высокие уровни трансляции в соматах (рис. 2 C ). Напротив, многие вспомогательные белки, связанные с рецептором глутамата (например, Cnih3 ) или каркасные белки (например,g., Shank1 , Dlg4 и Homer2 ) показали повышенный уровень трансляции в нейропиле (рис. 2 C ). Кроме того, мы обнаружили, что многие пресинаптические белки демонстрируют более высокую скорость синтеза белка в соматах (рис. 2 C ). Интересно, что мы идентифицировали несколько мРНК, кодируемых ядром, связанных с функцией митохондрий, которые демонстрировали повышенные уровни трансляции в нейропиле (например, Timm8a1 и Mrpl40 ) (Fig. 2 C ).
Рис. 2.
Функциональная сегрегация транскриптов, дифференциально транслируемых между соматой и нейропилем. ( A и B ) Термины GO, представляющие первые пять наиболее значительно обогащенных (FDR <0,05) белковых функциональных групп для транскриптов сомата-трансляции ( A ) и нейропиля ( B ). ( C ) Схема, изображающая белки глутаматергических синапсов. Соотношения нейропиля: соматы Ribo-seq (log 2 FC) имеют цветовую кодировку от оранжевого (больше соответствует сомате) до бирюзового (больше соответствует нейропилю).Взаимодействующие белки отображаются ближе друг к другу. Белки со сходными функциями сгруппированы вместе, а цикл синаптических пузырьков указан стрелками.
Профили транскрипции и трансляции мРНК в сомате и нейропиле доступны для загрузки и изучения через веб-интерфейс с возможностью поиска (https://public.brain.mpg.de/dashapps/localseq/). Эта интерактивная база данных позволяет зрителям сравнивать уровни трансляции транскриптов и мРНК между нейронными компартментами.
Большинство трансляционных изменений между соматическими и синаптическими областями можно объяснить различиями в изобилии РНК.
Уровень трансляции данного транскрипта пропорционален его количеству и плотности рибосом. Таким образом, мы задались вопросом, связана ли дифференциальная трансляция транскриптов сомата- и нейропил-трансляции с межкомпартментными изменениями уровней РНК (набор данных S3). В самом деле, транскрипты нейропиля с трансляцией вверх обнаруживают значительно более высокие соотношения нейропил: сомата RNA-seq по сравнению с генами с трансляцией соматы вверх (Fig. 3 A ). Чтобы подтвердить эти наблюдения in situ на срезах гиппокампа, мы провели флуоресцентную гибридизацию in situ с высоким разрешением (FISH) для 14 транскриптов-кандидатов со значительно различающимися уровнями трансляции между соматами и нейропилем (рис.3 B – D ). Обнаруженный сигнал гибридизации in situ был самым высоким в ожидаемом компартменте (т. е. сомата для сомата-трансляции вверх, рис. 3 B и D , и нейропил для нейропиля-трансляции вверх, рис. 3 C и ). Д ). Взятые вместе, анализы RNA-seq и FISH показали, что повышенная трансляция в соматах или нейропиле сопровождается более высокими уровнями РНК в том же самом нейронном компартменте.
Рис. 3.
Дифференциальная трансляция генов нейропиля и сомата-трансляции вверх сопровождается межкомпартментными изменениями уровней РНК.( A ) Коробчатая диаграмма, представляющая соотношение нейропиль: сомата RNA-seq (log 2 FC) для генов сомата (smt)-трансляция-вверх (оранжевый) и нейропиль (npl)-трансляция-вверх (бирюзовый) (DESeq2 ; Экспериментальные процедуры ). ( B и C ) ( Top ) Соотношения Neuropil:somata RNA- и Ribo-seq (log 2 FC) для генов-кандидатов smt-translation-up ( Gria2 , Uchl1118, , SV2B , SYP1 , GRIA1 , Snap25 , и Snap25 ) ( B ) и гены NPL-трансляции ( ACO2 , DLG4 , HPCAL4 , CNIH3 , DDN , Eef2 и Camk2a ) ( C ).( Внизу ) Сигнал FISH в области СА1 срезов гиппокампа крысы с использованием зондов против генов-кандидатов smt- ( B ) и npl-translation-up ( C ). Дендриты были иммуноокрашены антителом против MAP2 (фиолетовый). (Масштабная линейка, 50 мкм.) ( D ) Соотношение нейропилей: соматы точек мРНК относительно среднего соотношения нейропилей: сомата генов, повышающих трансляцию smt (*** P < 2,2e-16, Mann –Whitney U Тест между всеми генами smt-translation-up и всеми npl-translation-up).
Затем мы сравнили эффективность трансляции (TE) на уровне гена между нейропилем и соматой, рассчитав отношение следов рибосом (из Ribo-seq) к фрагментам мРНК (из RNA-seq) (17) в обоих компартментах (рис. 4). A и набор данных S4). Мы наблюдали хорошую корреляцию между значениями TE соматы и нейропиля, что указывает на то, что большинство транскриптов демонстрируют сходную регуляцию трансляции в обоих нейронных компартментах (рис. 4 A , R 2 = 0.92, P < 2.2e-16). Напр., Syngap1 демонстрирует низкое отношение следа к мРНК как в соматах, так и в нейропиле, указывая на относительно низкую трансляционную эффективность этого транскрипта (Fig. 4 A и B ). Напротив, Camk2a транслируется с высокой эффективностью (высокое отношение следа к мРНК) в обоих нейронных компартментах (фиг. 4 A и B ). Мы также идентифицировали несколько мРНК, которые демонстрировали значительно более высокие значения TE в сомах, включая, например, Kif5c (рис.4 A и B ). Таким образом, многие, но не все различия между компартментами в уровнях рибосомного следа могут быть объяснены различиями в количестве присутствующей мРНК.
Рис. 4.
Большинство транскриптов проявляют сходную трансляционную эффективность в сомате и нейропиле. ( A ) Корреляция эффективности трансляции (TE; log 2 Ribo-Seq/RNA-seq) в нейропиле и соматах ( R 2 = 0,92, P <2.2д-16). Выделены гены со значительно более высоким (TE , высокий , желтый) или более низким (TE , низкий , синий) TE, чем log 2 1,5 (FDR <0,05, DESeq2) как в сомате, так и в нейропиле. Гены со значительными различиями TE между соматами и нейропилем показаны красным. DESeq2 с FDR <0,05. Крайний ковер (серый) представляет распределение значений TE в соматах (ось x ) и нейропиле (ось y ). ( B ) Треки покрытия, представляющие средний след рибосомы или покрытие РНК для генов-кандидатов ( Syngap1 , Kif5c и Camk2a ) в нейропиле и сомате.Ось y указывает число считываний на миллион (об/мин). ( C и D ) Термины GO, представляющие значительно обогащенные (FDR <0,05) группы белковых функций для транскриптов TE low ( C ) и TE high ( D ). ( E ) Эмпирическая кумулятивная частота распределения (Ecdf) TE (log 2 FC) SFARI связанных с аутизмом (желтый) и других (черный) генов. P = 2,579e-05, критерий Колмогорова–Смирнова.
Первазивная трансляционная регуляция в соматах и нейропилях.
В обоих нейронных компартментах мы наблюдали широкое распределение эффективности трансляции с более чем 1000-кратной разницей между наиболее и наименее эффективно транслируемыми транскриптами в нейропиле (Fig. 4 A ). Мы идентифицировали 730 и 592 транскрипта, демонстрирующих значительно высокую или низкую эффективность трансляции, соответственно, как в сомате, так и в нейропиле (рис. 4 A и набор данных S4). Мы идентифицировали особенности генов, связанные с этими двумя группами, которые мы назвали TE low и TE high .Анализ GO выявил обогащение таких терминов, как «веретено» и «центр организации микротрубочек» для генов TE low (рис. 4 C ). Напротив, TE high генов были связаны с такими терминами, как «внутренний компонент мембраны синаптических пузырьков» и «внутренний компонент постсинаптической мембраны» (Fig. 4 D ). Как группа, транскрипты TE low имели более длинные кодирующие последовательности (CDS), что согласуется с предыдущими наблюдениями (25⇓–27) ( Приложение SI , рис.S5 А ). Поскольку гены факторов риска аутизма были описаны как исключительно длинные (28⇓–30), мы проанализировали значения TE транскриптов Инициативы по исследованию аутизма Фонда Саймонса (SFARI). Мы обнаружили, что транскрипты SFARI демонстрируют в целом более низкие значения TE по сравнению с другими генами (рис. 4 E ). На эффективность трансляции мРНК также влияют элементы внутри UTR, которые служат платформами для связывания регуляторных RBP (10, 12). Поскольку более длинные UTR содержат больше цис-действующих элементов (10, 12), мы исследовали длину 5′- и 3′-UTR трансляционно регулируемых транскриптов.Мы обнаружили, что гены TE low демонстрируют значительно более длинные 5′- и 3′-UTR (рис. 5 A и B ). Чтобы идентифицировать потенциальные RBP для UTR нейропиля, мы провели поиск известных консенсусных мотивов RBP (31) и определили, связаны ли группы транскриптов, имеющие одинаковые мотивы, с более высокими или более низкими значениями TE в нейропиле ( Experimental Procedures ). В общей сложности 131 мотив 3′-UTR, на которые нацелены 52 RBP (набор данных S5), были связаны с транскриптами, демонстрирующими значительно более высокие значения TE в нейропиле (рис.5 С ; для somata см. SI Приложение , рис. S5 B и набор данных S6). Например, в соответствии с их описанной ролью трансляционных энхансеров (32⇓-34), мотивы HNRNPK и MBNL1 были обнаружены в транскриптах, демонстрирующих значительно более высокие значения TE (Fig. 5 C ). С другой стороны, 155 мотивов 3′-UTR, на которые нацелены 90 RBP (набор данных S5), были связаны с транскриптами, демонстрирующими значительно более низкие значения TE нейропиля в нейропиле (Fig. 5 C ).Среди них мы идентифицировали, например, семейства CPEB, Hu (Elav) и PUF/Pumilio RBP, все известные своим репрессивным действием на трансляцию в нейронных отростках (35). Мы отмечаем, что ни один из мотивов RBP, обнаруженных нами в 5′-UTR нейропиля, не был связан с транскриптами, демонстрирующими значительно более высокий или более низкий TE нейропиля или соматы (наборы данных S7 и S8). Таким образом, наши результаты раскрывают идентичность потенциально новых регуляторов, которые связывают 3′-UTR и контролируют трансляцию прямо или косвенно, например, посредством регуляции полиаденилирования (34) или распада мРНК (35).
Рис. 5.
Особенности трансляционно регулируемых транскриптов в сомате и нейропиле. ( A и B ) Графики длины 5′ UTR ( A ) и 3′ UTR ( B ) (log 10 нуклеотидов (нт) для TE high (желтый), TE ) низкий (синий) и другие (серые) гены. Столбцы указывают 1,5 * IQR. * P < 0,05, **** P < 0,0001; односторонний тест ANOVA с последующим попарным тестом t с Бенджамини –Hochberg P регулировка значения.( C ) Показаны RBP-мотивы в 3′-UTR, связанные со значительно более низкими (синий) или более высокими (желтый) значениями TE нейропиля (значения q <0,05; критерий суммы рангов Уилкоксона) ( Экспериментальные процедуры ). ( D ) Обнаружение транслируемых uORF в нейронах гиппокампа. Сайты инициации трансляции картировали с помощью препарата харрингтонина (har), который накапливает рибосомы на стартовых кодонах. Всего в соматах и нейропиле обнаружено 766 uORF-содержащих нейрональных транскриптов.( E ) Треки покрытия, представляющие средние считывания рибосомного следа вдоль UTR (серый), обнаруженных uORF (оранжевый) или последовательности, кодирующей основной белок (синий) Dlg4 , Gria2 , Taok1 и Ppp1r9b в нейропиле. Ось y указывает число считываний на миллион (об/мин). ( F ) Наблюдаемое и ожидаемое соотношение TE high (бирюзовый), TE low (синий) и других (серых) транскриптов, содержащих uORF. ** P < 0.01, *** P < 0,001, **** P < 0,0001; гипергеометрический тест. ( G ) Neuropil TE (log 2 FC) измерения транскриптов, содержащих или не содержащих транслированные uORF («uORF») («нет uORF»). **** Р < 0,0001; Двухвыборочный тест Welch t . ( H ) Термины GO, представляющие восемь верхних, значительно (FDR <0,05) обогащают функциональные группы белков для uORF-содержащих транскриптов в нейропиле.
Верхние открытые рамки считывания (uORF) также играют важную роль в регуляции трансляции основной последовательности, кодирующей белок (36).В то время как считается, что большинство uORF оказывают негативное влияние на трансляцию нижестоящих ORF (36), сообщалось о нескольких примерах положительного действия uORF (37, 38). Мы идентифицировали транслируемые uORF в нейронных компартментах, используя интегрированный экспериментальный и вычислительный подход. Чтобы картировать восходящие сайты инициации трансляции в нейронных транскриптах, мы выполнили Ribo-seq на нейронах, обработанных препаратом харрингтонином, который вызывает накопление рибосом в стартовых кодонах (21) (рис.5 D и Экспериментальные методики ). Затем мы использовали конвейер ORF-RATER для идентификации и количественной оценки транслируемых uORF в данных нейропиля и соматы Ribo-seq ( Experimental Procedures ) (39). Всего мы идентифицировали 766 uORF-содержащих мРНК в компартментах нейронов (рис. 5 D и набор данных S9), включая новые (например, Gria2 , Taok1 , Dlg4 и Ppp1r9b ) (рис. 5 E и SI Приложение , рис.S5 C ) и ранее описанных (например, Atf4 и Ppp1r15b ) (38, 40) ( SI Приложение , рис. S5 D ). Сравнение транскриптов TE low и TE high выявило чрезмерную представленность транскриптов, содержащих uORF, в группе TE low и недостаточную представленность транскриптов, содержащих uORF, в группе TE high (рис. 5 F ). ). Кроме того, транскрипты, содержащие uORF, показали значительно более низкое значение медианы TE нейропиля по сравнению с мРНК, не содержащими uORF (рис.5 G и SI Приложение , рис. S5 E для соматы). Затем, используя данные нейропиля Ribo-seq, мы рассчитали относительную плотность uORF к плотности рибосом CDS для каждой uORF. Интересно, что относительная плотность рибосом uORF:CDS варьировалась от 0,1 до 1000, что указывает на широкое распространение опосредованной uORF трансляционной репрессии в нейропиле ( SI Annex , Fig. S5 F ). Многие uORF демонстрируют соотношение плотности рибосом uORF:CDS больше 1, что указывает на то, что uORF часто действуют как репрессоры трансляции CDS.Анализ GO показал, что описанные выше uORF-содержащие мРНК нейропиля и сомата были значительно обогащены такими терминами, как «позитивная регуляция сборки синапсов», «регуляция мембранного потенциала» и «поведение» (Fig. 5 H ). Эти находки подчеркивают, что uORFs являются важным трансляционным регуляторным элементом, присутствующим во многих транскриптах в соматических и синаптических областях.
Обсуждение
Используя профилирование рибосом, мы обнаружили тысячи видов мРНК, которые транслируются в синаптических областях, что резко увеличило вклад текущего локального синтеза белка в белковый пул, обнаруживаемый в дендритах, аксонах или синапсах (41⇓⇓–44) .Действительно, среди локально переведенного мРНКа, мы определили большинство семейств белков, в том числе сигнальных молекул (киназы или фосфатазы), ионные каналов, метаботропные и ионотропных рецепторов, молекул клеточной адгезии, каркасные белков, а также регуляторов цитоскелета ремоделирования или перевода.
Многие транскрипты были обнаружены дифференцированно переведены между нейронными отсеках. Открытый вопрос в поле отнесся вклад локального синтеза в общий пул конкретного белка.Наши данные показывают, что большинство белков синтезируется в обоих компартментах. Мы отмечаем, что более 800 мРНК обнаруживают повышенные уровни трансляции в нейропиле, предполагая, что большинство этих белков происходят из местного источника. Для многих транскриптов обилие мРНК было положительно связано с различиями в уровне трансляции между соматами и нейропилем, как это наблюдалось ранее в развивающихся нейронах, происходящих из эмбриональных стволовых клеток мыши (45). Примечательно, что транскрипты neuropil-translation-up часто кодируют сигнальные и каркасные белки, которые играют важную роль в поддержании и модификации синаптической силы.Интересно, что мы обнаружили несколько митохондриальных мРНК, которые демонстрировали усиленную трансляцию нейропиля. Недавно было показано, что эндосомы могут действовать как платформы для локальной трансляции кандидатов в митохондриальные мРНК (46). Таким образом, заманчиво предположить, что локальная трансляция играет роль в поддержании митохондрий, которые, в свою очередь, подпитывают синтез белка вблизи синапсов во время пластичности (47). В совокупности наши результаты предполагают, что повышенные уровни трансляции специфического подмножества транскриптов в нейропиле, вероятно, обеспечивают средства для обеспечения эффективного производства ключевых синаптических белков в очень удаленных местах от тела клетки.
Напротив, транскрипты с повышенным уровнем трансляции в соматах часто кодируют трансмембранные белки. Этот класс белков обычно обрабатывается через несколько связанных с мембраной органелл (включая эндоплазматический ретикулум [ER] и аппарат Гольджи [GA]), где они сворачиваются, собираются и биохимически модифицируются перед доставкой на поверхность нейронов (48). . Однако недавние исследования показали, что сотни белков поверхности нейронов (например, субъединица AMPAR GluA1) обходят созревание GA и, вероятно, перемещаются непосредственно из ER на поверхность нейронов (49, 50).Т.о., хотя основной синтез и посттрансляционная модификация трансмембранных белков могут происходить в соматических ER и GA, небольшая остаточная фракция этого класса белков может подвергаться локальной трансляции «по требованию» для тонкой настройки синаптической силы.
Используя комбинацию микродиссекции с Ribo- и RNA-seq, мы обнаружили, что большинство транскриптов проявляют сходную регуляцию трансляции в сомате и нейропиле. В обоих нейронных компартментах мы обнаружили обширную трансляционную регуляцию с неожиданно высоким динамическим диапазоном эффективности трансляции транскриптов.Среди механизмов, регулирующих синтез белков в соматических и синаптических областях, мы идентифицировали uORF-опосредованный контроль трансляции. Эти данные хорошо согласуются с предыдущими исследованиями, раскрывающими роль uORFs в регуляции трансляции двух транскриптов-кандидатов в нейронных отростках (51, 52). uORF-опосредованный контроль трансляции часто точно регулируется фосфорилированием эукариотического фактора инициации 2α (eIF2α) (53). Фосфорилирование eIF2α ингибирует глобальную трансляцию, одновременно приводя к парадоксальному увеличению трансляции субнабора транскриптов, несущих uORF (54).Многие манипуляции с клеточной и синаптической активностью модулируют статус фосфорилирования eIF2α в нейронах in vivo и in vitro (54⇓⇓–57). Т.о., управляемое активностью фосфорилирование eIF2α может действовать как переключатель для усиления локальной эффективности трансляции uORF-содержащих транскриптов, кодирующих ключевые белки, связанные с пластичностью. Примечательно, что трансляционная регуляция некоторых транскриптов, содержащих uORF, нечувствительна к изменениям статуса фосфорилирования eIF2α (например, регуляторной субъединицы протеинфосфатазы 1 CReP [ Ppp1r15b ]) (40).
Электронно-микроскопические (ЭМ) исследования показали, что распределение рибосом вдоль отростков нейронов неоднородно, с избирательной локализацией белковообразующих машин (т. е. полирибосом, более трех рибосом на мРНК) под синапсами, в то время как лишь несколько полирибосомы могут наблюдаться в стволах дендритов СА1 (58, 59). Стержни дендритов могут быть в основном заселены моносомами (т.е. одной рибосомой на мРНК), которые не могут быть визуализированы с помощью ЭМ, но также представляют собой машины, производящие активный белок в синаптических областях (16).Действительно, недавнее исследование сверхвысокого разрешения, которое, вероятно, обнаруживает как моносомы, так и полисомы, выявило большую плотность рибосом в дендритах по сравнению с исследованиями ЭМ (60). Эти наблюдения поднимают интригующие вопросы об определении локальных компартментов трансляции: синтезируются ли различные виды белков в отдельных субрегионах нейронных отростков (например, шипы или дендритные стержни)? И: может ли эффективность трансляции одного и того же транскрипта варьироваться в зависимости от того, локализован он под синапсами или в других дендритных регионах? Эти вопросы подготовили почву для будущих исследований, характеризующих трансляционный ландшафт в субрегионах нейронов с большим пространственным разрешением с использованием, например, профилирования рибосом, специфичных для близости.
Экспериментальные процедуры
Животные.
Беременных самок крыс, свободных от специфических патогенов (Charles River Laboratories) с отложенным сроком содержания в течение 1 недели содержали в помещении для животных Института исследований мозга им. Макса Планка при 12/12-часовом цикле свет/темнота с едой и водой без ограничений до появления помета был рожден. Культивируемые нейроны были получены от крысят Sprague-Dawley P0 (послеродовой день 0) (как самцов, так и самок, идентификатор исследовательского ресурса: 734476). Детенышей убивали обезглавливанием. Процедуры содержания и умерщвления, связанные с обращением с животными и уходом за ними, проводились в соответствии с институциональными рекомендациями, которые соответствуют национальным и международным законам и политикам (Директива 2010/63/ЕС, немецкий закон о защите животных, руководящие принципы Федерации европейских лабораторных ассоциаций животных). ).Животные были убиты в соответствии с приложением 2 § 2 Abs. 2 Tierschutz-Versichstier-Verordnung. Количество животных сообщалось в местные органы власти (Regierungspräsidium Darmstadt, регистрационные номера: V54-19c20/15-F126/1020 и V54-19c20/15-F126/1023).
Библиотеки Ribo- и RNA-Seq из микропрепарированных крыс Somata и Neuropil.
Библиотеки Total Ribo-seq (включая моносомы и полисомы) и RNA-seq из микродиссекции сомата крысы и нейропиля в трех биологических повторностях были созданы ранее (16) ( SI, Приложение , Таблица S1).Короче говоря, соматы и нейропиль были микродиссекции у 4-недельных самцов крыс. Образцы тканей гомогенизировали в буфере для лизиса полисом (20 мМ трис, рН 7,5, 150 мМ NaCl, 5 мМ MgCl 2 , 24 ед/мл турбоДНазы, 100 мкг/мл циклогексимида, 1 мМ дитиотреитола (ДТТ), 1% тритона X). -100 и смесь ингибиторов протеазы [Roche]) путем обливания в стеклянном гомогенизаторе. После 10-кратного растирания лизата с помощью шприца 23 размера образцы охлаждали на льду в течение 10 минут и очищали путем двух центрифугирований при 16 100 × g в течение 6 минут.Из лизатов соматы и нейропиля одновременно готовили библиотеки Ribo-seq и RNA-seq. Для Ribo-seq лизаты нейропиля и сомата, содержащие одинаковое количество тотальной РНК, расщепляли 0,5 ЕД/мкг РНКазы I (Epicentre) при встряхивании в течение 45 мин при 400 об/мин при 24 °C. Реакции расщепления нуклеазой быстро охлаждали и центрифугировали и добавляли 10 мкл ингибитора SUPERaseIN*RNase. Затем образцы наслаивали на подушку из 34% сахарозы, приготовленную в массе/объеме в градиентном буфере с добавлением 20 ед/мкл ингибитора SUPERaseIN*РНКазы.Частицы 80S осаждали центрифугированием в роторе SW55Ti в течение 3 ч 30 мин при 55000 об/мин при 4 °С. Библиотеки Ribo-seq были приготовлены в соответствии со ссылкой. 61 с модификациями, описанными в ref. 16. Суммарную РНК выделяли из лизатов тканей с использованием набора для микропрепаратов Direct-zol RNA (Zymo). Целостность РНК оценивали с помощью набора Agilent RNA 6000 Nano. Библиотеки тотальной РНК-seq нейропиля и соматы крыс готовили из равного количества тотальной РНК с использованием золотого набора для подготовки библиотеки тотальной РНК TruSeq (Illumina) (16).Библиотеки секвенировали на Illumina NextSeq500 с использованием одноконцевой серии 52 и 75 п.н. для Ribo-seq и RNA-seq соответственно.
Библиотеки RNA-Seq из культур, обогащенных нейронами и глиями.
Культуры, обогащенные нейронами и глиями, были приготовлены из того же помета, как описано ранее (12). Гиппокамп детенышей крысы возраста P0d выделяли и растирали после переваривания папаином. Обе культуры высевали на чашки для культивирования клеток диаметром 60 мм. Для приготовления культур, обогащенных нейронами гиппокампа, клетки высевали на 60-мм чашки для культивирования клеток, покрытые поли-d-лизином, и обрабатывали, как описано выше, с помощью Ara-C (Sigma) в конечной концентрации 5 мкМ в течение 48 часов.Через 48 ч среду заменяли предварительно кондиционированной ростовой средой и клетки культивировали до 21 дня in vitro (DIV). Для получения культур, обогащенных глией, клетки высевали на непокрытые 60-мм чашки для культивирования клеток в кондиционированной минимально необходимой среде (минимальная необходимая среда, 10% лошадиной сыворотки, 0,6% глюкозы [масса/объем]). В 7 DIV среду заменяли предварительно кондиционированной ростовой средой и клетки культивировали до 21 DIV. Были подготовлены четыре независимых биологических повтора. РНК выделяли с использованием набора для микропрепаратов Direct-zol RNA (Zymo).Целостность РНК оценивали с помощью набора Agilent RNA 6000 Nano. Библиотеки mRNA-seq готовили, начиная с примерно 200 нг общей РНК, используя набор для подготовки библиотеки TruSeq stranded mRNA (Illumina). Библиотеки были секвенированы на Illumina NextSeq500 с использованием одностороннего цикла 75 п.н.
Библиотеки RNA-Seq из иммунопреципитации меченых рибосом.
Библиотеки последовательностей ввода и трансляции рибосомной аффинной очистки (TRAP) из гиппокампа Camk2a-Cre-RiboTag или срезов соматы/нейропила мышей Wfs1-Cre-RiboTag были созданы ранее (16) ( SI Приложение , Таблица S1 ).
Библиотеки Ribo-Seq из культивируемых нейронов гиппокампа крыс, обработанных харрингтонином.
Диссоциированные нейроны гиппокампа крыс получали из детенышей крысы возраста P0d, как описано ранее (62). Нейроны гиппокампа высевали с плотностью 31 250 клеток/см 2 на покрытые поли-d-лизином 100-мм чашки и культивировали в предварительно кондиционированной питательной среде (Neurobasal-A, B27, GlutaMAX, 30% супернатант культуры глии, 15 % супернатанта культуры коры) для 21 DIV. В 1 DIV клетки обрабатывали Ara-C (Sigma) в конечной концентрации 5 мкМ для предотвращения избыточного роста ненейрональных клеток.Через 48 ч среду заменяли на предварительно кондиционированную среду для роста и клетки культивировали до 21 DIV. Клетки подкармливали 1 мл прекондиционированной среды каждые 7 дней. Были приготовлены три независимых биологических повтора. За 24 ч до обработки лекарственным средством клеточную среду доводили до 8 мл на чашку. В соответствующих экспериментах харрингтонин (LKT Laboratories) добавляли до конечной концентрации 2 мкг/мл из исходного раствора 5 мг/мл в 100% этаноле. Клетки возвращали в инкубатор при 37°С на 15, 30, 45, 90 или 150 с.Циклогексимид добавляли до конечной концентрации 100 мкг/мл из исходного раствора 50 мг/мл в 100% этаноле. После добавления препарата клетки возвращали в инкубатор при 37°С на 1 мин. После инкубации с циклогексимидом клетки немедленно помещали на лед и дважды промывали ледяным фосфатно-солевым буфером (PBS) плюс 100 мкг/мл циклогексимида и соскребали в буфере для лизиса полисом (20 мМ Трис, рН 7,5, 150 мМ NaCl, 5 мМ MgCl 2 , 24 ед/мл турбоДНКазы, 100 мкг/мл циклогексимида, 1 мМ DTT, 1% Triton-X-100 и смесь ингибиторов протеазы [Roche]) (21).После соскоба лизаты растирали 10 раз с помощью шприца 23G; образцы охлаждали на льду в течение 10 мин, а затем очищали центрифугированием при 16 100 × g в течение 10 мин. Библиотеки Ribo-seq из культур нейронов гиппокампа крыс, обработанных харрингтонином в течение 0, 15, 30, 45, 90 и 150 с, готовили, как описано выше. Наборы данных за 0, 30 и 90 с были ранее опубликованы в ссылке. 16 ( Приложение SI , таблица S1).
Гибридизация in situ в срезах гиппокампа мозга.
Четырехнедельным самцам крыс перфузировали 1× PBS без РНКазы и фиксирующий раствор (4% (об./об.) параформальдегида (PFA), 4% (вес./об.) сахарозы в 1× PBS без РНКазы) . Мозг вырезали и фиксировали еще в течение часа при комнатной температуре. Мозг подвергали криозащите два дня подряд при температуре 4 °C. В 15% (вес/объем) сахарозы в 1× PBS без РНКазы в 1-й день, затем в 30% (вес/объем) сахарозы в 1× PBS без РНКазы на 2-й день. Гиппокампы подвергали криосрезу толщиной 30 мкм.
Флуоресцентную гибридизацию in situ проводили с использованием набора QuantiGene ViewRNA (Thermo Fisher), в основном следуя инструкциям производителя.Вкратце, срезы гиппокампа постфиксировали в течение 10 минут при комнатной температуре в фиксирующем растворе (4% [объем/объем] PFA, 5,4% [вес/объем] глюкозы, 0,01 М метапериодата натрия в 1× лизин-фосфатном буфере). Изготовитель рекомендовал отказаться от обработки протеиназой К, чтобы сохранить целостность дендритов. Срезы пермеабилизировали в течение 20 минут с использованием детергентного буфера из набора. Детектирующие зонды инкубировали в течение ночи при 40 °C. Зонды преамплификации, амплификации и метки инкубировали в течение 60 мин при 40 °C соответственно, промывая три раза по 5 мин между каждым этапом.После завершения гибридизации in situ срезы промывали 1× PBS и инкубировали в блокирующем буфере (4% [об./об.] козьей сыворотки 1× PBS) в течение 1 ч при комнатной температуре. Первичное антитело (gp-анти-MAP2, SYSY 188004, 1:1000) инкубировали в течение ночи в блокирующем буфере при 4 °C. Срезы промывали пять раз по 10 мин в 1× PBS и вторичное антитело (gt-анти-gp Alexa 647, Thermo Fisher A21450, 1:500) инкубировали в блокирующем буфере в течение 5 ч при комнатной температуре. Срезы промывали 1× PBS и окрашивали ядра DAPI в течение 3 мин при комнатной температуре.Срезы монтировались в AquaPolyMount.
Срезы визуализировали с помощью конфокального микроскопа Zeiss LSM780 и масляного объектива 40× (числовая апертура [NA] 1,3). Z-стеки, охватывающие весь объем среза, были получены с использованием соответствующих лазерных линий возбуждения и окон спектрального детектирования. Сигнал мРНК был расширен для лучшей визуализации. Для анализа использовались необработанные нерасширенные изображения.
Для подсчета точек мРНК в соматическом слое и слое нейропиля использовали собственный скрипт Python.В нейропиле точки, солокализованные с сигналом DAPI (возникающие из глии или интернейронов), были исключены из анализа. Подсчеты нормализовали по площади и для каждого среза рассчитывали отношение нейропиля к соматам. Среднее отношение нейропиля к сомате рассчитывали для генов-мишеней, повышающих трансляцию соматы. Все отношения нейропиля к соматам делили на это среднее.
Анализ данных.
Выравнивание генома и транскриптома библиотек профилирования рибосом.
Адаптеры для секвенирования были обрезаны с использованием программного обеспечения Cutadapt версии 1.15 (63) со следующими аргументами: — отрезать 1–минимум -длина 22 –отбросить-необрезать –перекрыть 3 -е 0,2 . Расширенный уникальный молекулярный идентификатор (UMI) был сконструирован из двух случайных нуклеотидов (nts) праймера обратной транскрипции и пяти случайных нуклеотидов линкера и добавлен к строке описания FASTQ с использованием пользовательского сценария Perl. Для удаления прочтений, происходящих из некодирующей РНК (нкРНК, т. е. рРНК), обрезанные прочтения выравнивали с крысиной нкРНК с помощью Bowtie2 версии 2.3.5.1 (–очень чувствительные) (64) и выровненные чтения были отброшены. Остальные чтения были выровнены с геномом крысы (rn6) с помощью расщепленного выравнивателя STAR версии 2.7.3.a (65) со следующими аргументами: –twopassMode Basic –twopass1readsN -1 –seedSearchStartLmax 15 –outSJfilterOverhangMin 15 8 8 8 –outFilterMismatchNoverReadLmax 0.1. Для получения координат расшифровки использовался квантовый режим STAR (-quantMode). На протяжении всего исследования выравнивание генома использовалось для анализа дифференциальной экспрессии и анализа геномных признаков.Для всех других анализов использовали выравнивание транскриптома. Индекс генома STAR был построен с использованием аннотации, загруженной из обозревателя таблиц Калифорнийского университета в Санта-Круз (UCSC) (66). Дубликаты ПЦР подавлялись с помощью пользовательского сценария Perl, а выравнивания, помеченные как вторичные, отбрасывались перед анализом. Для анализа использовались только следы размером от 24 до 34 нт.
Выравнивание генома библиотек РНК.
Адаптеры для секвенирования и низкокачественные нуклеотиды были обрезаны с помощью программного обеспечения Cutadapt версии 1.15 (63) со следующими аргументами: –minimum-length 25–nextseq-trim = 20 . Обрезанные чтения выравнивали с геномом крысы (rn6) или мыши (mm10) с помощью STAR версии 2.7.3a (65).
Анализ геномных признаков.
Координаты геномных признаков (CDS, 3′-UTR, 5′-UTR, интрон) были загружены из браузера таблиц UCSC в формате BED (66). Bedtools версии 2.26.0 (67) использовался для преобразования BAM в файлы BED и для выявления перекрытий чтения с отдельными функциями.
Трехнуклеотидная периодичность.
Смещения P-сайтов были определены для разных длин посадочных мест. Каждая начальная позиция следа определяла кадр следа относительно аннотированного стартового кодона. Считывания следа были фактически проецированы назад на стартовый кодон, и были рассчитаны смещения от начала и конца считывания. Мы использовали каждое чтение заданной длины и накапливали наиболее вероятное смещение и кадр. Затем из его длины и ранее определенного смещения было выведено положение P-сайта на прочитанный след.Все положения P-сайтов были нанесены на 100 нуклеотидов вокруг стартового и стоп-кодонов, а также в центре транскрипта. Чтобы скорректировать различия в скорости трансляции между генами, покрытие P-сайтов каждого гена было нормализовано к его среднему покрытию следа. Оценивали покрытие нуклеотидами в положениях 0, 1 и 2 рамки считывания. Однофакторный дисперсионный анализ (ANOVA) использовался для определения того, отличалась ли наблюдаемая доля кадров от ожидаемой доли кадров. Значительное значение P опровергает нулевую гипотезу о том, что все кадры имеют ожидаемое покрытие P-узла.
Визуализация треков браузера генома.
Выравнивания следов были преобразованы в формат файлов BedGraph с помощью Bedtools версии 2.26.0 и визуализированы в виде пользовательских дорожек в браузере UCSC Genome Browser (68). Покрытия посадочных мест были скорректированы с учетом глубины секвенирования.
Дифференциальный экспрессионный анализ.
RNA-seq и Ribo-seq соотношения нейропиля:сомы.
Как для полного секвенирования РНК, так и для библиотек следов рибосом из соматы и нейропиля программное обеспечение имеет версию 2.0,0 (69) использовали для подсчета количества на ген из прочтений, которые были выровнены с геномом крысы. Были учтены все аннотированные изоформы транскриптов. Необработанные подсчеты вводили в DESeq2 версии 1.30.1 и использовали логарифмическую усадку кратного изменения (LFC) (18). Только гены со скорректированным значением P показаны на рис. 1 B .
RiboTag IP:соотношения входных данных и отношения культур, обогащенных нейронами:обогащенных глией.
Программное обеспечение featureCounts версии 2.0.0 (69) использовалось для подсчета количества на ген из прочтений, сопоставленных с геномом (mm10, rn6).Были учтены все аннотированные изоформы транскриптов. Необработанные подсчеты были введены в DESeq2 версии 1.30.1, и использовалась усадка LFC (18).
Анализ онтологии генов.
Анализ онтологии генов был проведен для генов нейропиля и сомата-трансляции. В качестве фона использовали все обнаруженные гены (базовое среднее значение больше нуля и с скорректированным значением P ) без примесей. Анализ обогащения GO был выполнен для полной аннотации клеточного компонента с использованием теста чрезмерной репрезентативности PANTHER (70, 71).Использовался точный критерий Фишера, и учитывались только термины GO с коэффициентом ложного обнаружения (FDR) менее 0,05. Были сохранены наиболее конкретные термины GO для каждой ветви. Были визуализированы пять лучших терминов GO с самыми высокими показателями обогащения.
Анализ онтологии генов был выполнен для транскриптов, содержащих uORF. В качестве фона использовали все обнаруженные гены в нейропиле и сомате (базовое среднее значение больше нуля) без контаминантов. Анализ обогащения GO был выполнен для полной аннотации биологического процесса с использованием теста чрезмерной репрезентативности PANTHER (70, 71).Использовался точный критерий Фишера, и учитывались только члены GO с FDR менее 0,05. Были сохранены наиболее конкретные термины GO для каждой ветви. Все значимые термины GO были визуализированы.
Генный онтологический анализ был выполнен для транскриптов TE high и TE low . В качестве фона использовали все обнаруженные гены в нейропиле и сомате (базовое среднее значение больше нуля) без контаминантов. Анализ обогащения GO был выполнен для полной аннотации клеточного компонента с использованием теста чрезмерной репрезентативности PANTHER (70, 71).В анализе использовались только термины GO с не менее чем 50 генами в фоновом наборе. Использовался точный критерий Фишера, и учитывались только члены GO с FDR менее 0,05. Были сохранены наиболее конкретные термины GO для каждой ветви. Все значимые термины GO были визуализированы.
Расчет трансляционной эффективности.
Количество рибосом на транскрипт оценивали путем интеграции библиотек Ribo-seq и RNA-seq для расчета значений TE в нейропиле. Необработанные подсчеты Ribo-seq и RNA-seq, попадающие в CDS гена, вводили в DESeq2 версии 1.30.1 и использовалась усадка LFC (18). Значения TE, которые были либо значительно выше, чем log 2 (1,5) в нейропиле и соматах, либо меньше, чем log 2 (1,5) в нейропиле и соматах, были отнесены к TE high и TE low , соответственно [lfcThreshold = log 2 (1,5) с FDR <0,05]. Учитывались только гены с baseMean больше 10 в нейропиле и соматах. В план эксперимента был добавлен термин взаимодействия для сравнения значений TE между нейропилем и соматой (72).
Трансляционная эффективность генов аутизма.
Гены, которые, как известно, связаны с расстройствами аутистического спектра, были загружены из базы данных генов SFARI (https://www.sfari.org). Символы генов человека были преобразованы в символы генов крыс. Гены с оценкой SFARI 1 и 2 считались генами аутизма.
Анализ мотивов 3′ и 5′ UTR.
мотива RBP (человека, крысы и мыши) были загружены в виде взвешенных по положению матриц из общедоступной базы данных ATtRACT (31).Инструмент FIMO из пакета MEME версии 5.1.1 использовался для сканирования 5′- и 3′-UTR на наличие мотивов с использованием порога по умолчанию (значение P = 1e-4) и предварительно рассчитанной фоновой модели нуклеотидов, полученной из последовательностей запроса ( 73). Для анализа рассматривались только гены с наличием мотива RBP. Для каждого идентифицированного мотива RBP гены, содержащие мотив, группировали и рассчитывали среднее значение TE. Критерий суммы рангов Уилкоксона был проведен для проверки того, отличается ли медиана ТЕ данной группы мотивов RBP от медианы ТЕ всех генов, не содержащих этот мотив.
Обнаружение транслируемых uORF.
Конвейер ORF-RATER (https://github.com/alexfields/ORF-RATER) был запущен, как описано ранее (39), начиная с harringtonine 150 s, а также файлов neuropil и somata BAM. Обратите внимание, что возможно, что переведенной uORF может быть присвоена низкая оценка, поскольку ORF-RATER настроен на указание сайтов трансляции с самой высокой степенью достоверности за счет увеличения частоты ложноотрицательных результатов (74). Использовали следующие параметры: «—codons NTG» для типов ORF, «—minrdlen 28 —maxrdlen 34» для образцов, обработанных харрингтонином, «—minrdlen 27 —maxrdlen 34» для образцов нейропиля и соматы.Учитывались только uORF с оценкой не менее 0,7, длиной не менее трех кодонов и не менее одного числа в каждом из повторов нейропиля и соматы.
Отношение uORF к плотности рибосом CDS.
Плотность рибосом uORF или CDS рассчитывали как количество следов рибосомы, деленное на длину uORF или CDS соответственно. Относительную плотность рибосом рассчитывали как плотность рибосом uORF, деленную на плотность рибосом CDS.
Анализ характеристик расшифровки.
Длины 5′- и 3′-UTR были рассчитаны на основе аннотации Rattus norvegicus версии 6 (rn6). Длины 3′-UTR были скорректированы в соответствии с вновь идентифицированными изоформами 3′-UTR, описанными в ref. 12. Для генов с несколькими изоформами 5′-UTR была выбрана самая длинная последовательность 5′-UTR, отдавая предпочтение выбранным изоформам. Для генов с несколькими 3′-UTR была выбрана наиболее экспрессируемая изоформа 3′-UTR (12).
Для сравнения длин 5′-UTR между «TE high «, «TE low » и «другими» рассматривались только 5′-UTR с минимальной длиной 10 нуклеотидов и максимальной длиной 5000 нуклеотидов.Для сравнения длин 3′-UTR между «TE high «, «TE low » и «другими» рассматривались только 3′-UTR с минимальной длиной 50 нуклеотидов и максимальной длиной 10000 нуклеотидов.
Для сравнения длин 3′-UTR между генами neuropil-translation-up и somata-translation-up была рассмотрена изоформа 3′-UTR с самой высокой экспрессией в гиппокампе на семейство генов (12).
Анализ метагена и расчет скорости удлинения.
Охват каждого гена проецировался вдоль CDS в координатах транскрипта (только экзоны).Гены с длиной CDS короче 440 кодонов были исключены из анализа. Каждый профиль метагена масштабировали по среднему покрытию между кодоном 400 и 20 кодонами перед стоп-кодоном. Для каждой временной точки профили метагенов были сглажены со скользящим средним окном в 30 кодонов. Для каждой группы треки покрытия накапливались, усреднялись и нормализовались к нулевому условию. Базовый уровень охвата был определен как 85% охвата необработанных образцов. Первое положительное пересечение между дорожкой покрытия, обработанной харрингтонином, и дорожкой покрытия базовой линии определяло положение пересечения в кодонах.Коэффициенты элонгации рассчитывали как наклон линейной регрессии между временем инкубации с харрингтонином для каждой дорожки и положением пересечения в кодонах.
Статистический анализ.
Статистическая значимость и проведенные тесты указаны в подписях к рисункам. Статистический анализ был выполнен с использованием MATLAB и R.
Доступность данных
Подробности о доступности данных можно найти в SI Приложение , Таблица S1. Номер доступа для необработанных данных секвенирования, опубликованных ранее в ref.16 — Биопроект Национального центра биотехнологической информации (NCBI): PRJNA550323. Номер доступа для необработанных данных секвенирования, представленных в этой статье, — NCBI BioProject: PRJNA634994. Все биоинформационные инструменты, использованные в этом исследовании, содержатся в одной модульной программе на C++ под названием RiboTools. Исходный код и дополнительные сведения об алгоритмах можно найти в нашем репозитории GitHub (DOI: 10.5281/zenodo.3579508). Другие скрипты и коды анализа доступны по запросу.
Благодарности
Благодарим Елену Чырдаеву за помощь в подготовке библиотеки мРНК.А.Б. поддерживается долгосрочной постдокторской стипендией Европейской организации молекулярной биологии (EMBO) (EMBO ALTF 331-2017). E.M.S. финансируется Обществом Макса Планка, наградой Advanced Investigator от Европейского исследовательского совета (грант 743216), Deutsche Forschungsgemeinschaft (DFG) Collaborative Research Center (CRC) 1080: Molecular and Cellular Mechanisms of Neural Homeostasis и DFG CRC 902: Molecular Principles. регуляции на основе РНК.
Сноски
- Принято 8 сентября 2021 г.
Вклад авторов: C.G., A.B. и E.M.S. проектное исследование; К.Г., А.Б., Б.Н.-А., А.К., И.Б. и С.т.Д. проведенное исследование; К.Г., А.Б. и Г.Т. проанализированные данные; и А.Б. и E.M.S. написал бумагу.
Рецензенты: CMA, Центр геномной и экспериментальной медицины Эдинбургского университета; и Е.К., Нью-Йоркский университет.
Авторы заявляют об отсутствии конкурирующих интересов.
См. в Интернете связанный контент, например комментарии.
Эта статья содержит вспомогательную информацию в Интернете по адресу https://www.pnas.org/lookup/suppl/doi:10.1073/pnas.2113929118/-/DCSupplemental.
- Copyright © 2021 Автор(ы). Опубликовано ПНАС.
Патологические процессы в водянистой влаге, обусловленные атрофией радужной оболочки, предрасполагают к раннему отторжению трансплантата роговицы у человека и мышей
Митохондрии — это больше, чем энергетический центр клетки; они служат медиаторами передачи сигналов гипоксических/активных форм кислорода, окислительного стресса, клеточной дифференцировки и клеточного цикла (-17-).Как мы показали выше, ПЭМ-изображения и протеомный подход свидетельствовали о повреждении митохондрий в CEnC от буллезной кератопатии. Затем мы провели транскриптомный анализ CEnC, подвергшихся воздействию патологического AqH. Мы получили первичную ткань CEnC от 15 пациентов, перенесших трансплантацию роговицы для лечения дисфункции CEnC (таблица S4). В качестве здорового контроля использовали ткань CEnC из четырех роговиц, полученную из Seattle Eye Bank. После выделения тканевой РНК мы оценили качество РНК для секвенирования РНК (RNA-seq).Из 15 глаз с дисфункцией CEnC адекватное число целостности РНК (RIN) выше 6 было достигнуто только в семи глазах из-за тяжелой потери CEnC в остальных восьми глазах. Следовательно, и поскольку мы предположили, что изменения биологических путей должны индуцироваться в CEnC, подвергшихся воздействию патологического AqH, мы охарактеризовали последствия транскрипции в CEnC у семи человек, перенесших трансплантацию роговицы. Мы идентифицировали 2385 с повышающей регуляцией и 2880 с пониженной регуляцией среди 40 448 генов (рис. 3, A и B), которые были статистически значимыми со значениями не-FDR P менее 0.05. Анализ основных компонентов (PC) показал отчетливую взаимосвязь между глазами с дисфункцией CEnC и здоровыми глазами (PC1, 42,0%; PC2, 11,7%; PC3, 9,2%), что составляет 62,9% дисперсии, наблюдаемой между наборами данных об экспрессии генов (рис. 3С). Два из шести генов, связанных с наследственными эндотелиальными дистрофиями роговицы [цинковые пальцы, связывающие E-box гомеобокс 1 ( ZEB1 ) и член 11 семейства 4 переносчиков растворенных веществ ( SLC4A11 )], были значительно подавлены при буллезной кератопатии CEnCs (понижающая регуляция на 3.0- и 3,7-кратное, P = 6,38 × 10 -5 и 1,97 × 10 -3 соответственно) по сравнению со здоровым контролем, в то время как четыре цепи α2 коллагена типа VIII ( COL8A2 ), фактор транскрипции 4 ( TCF4 ), домены гомологии липоксигеназы 1 ( LOXHD1 ) и белок, связывающий АТФ/гуанозинтрифосфат, подобный 1 ( AGBL1 )]. В то время как CEnC имеют минимальную экспрессию рецепторов цитокинов, чтобы избежать реакции на внешние раздражители в устойчивом состоянии ( 18 ), экспрессия генов рецепторов цитокинов, таких как интерлейкин ( IL )–2 r , IL-4r и IL-17r , интерферон-γr ( IFN- γ r ) и индукторы апоптоза, лиганд программируемой клеточной гибели ( PD-L2 ) и кластер дифференцировки 86 ( CD86 ). повышается, что позволяет предположить, что CEnCs могут стать чувствительными к повышенным цитокинам при AqH буллезной кератопатии ( 10 ).Кроме того, в соответствии с нашими данными электронной микроскопии, гены, связанные с митохондриями, такие как гены митохондриальной креатинкиназы ( CKMT ), гены АТФ-синтазы ( ATPs ) и переносчика растворенных веществ ( SLCs ), были значительно подавлены в глаза с буллезной кератопатией по сравнению со здоровым контролем (рис. 3D). Транскрипты генов, которые показали значительно разные профили экспрессии между здоровыми глазами и глазами с буллезной кератопатией, были дополнительно проанализированы для обогащения функциональной онтологии генов (GO), которая идентифицировала несколько текущих клеточных компонентов.Гены со значительно более высокой экспрессией в глазах с буллезной кератопатией продемонстрировали значительное обогащение для нескольких функциональных путей, таких как организация внеклеточного матрикса, клеточная адгезия, интегрин-опосредованный путь, передача сигнала, IFN-γ-опосредованный путь, иммунный ответ и воспалительный ответ (рис. 3E). ). С другой стороны, гены со значительно более низкой экспрессией в глазах с буллезной кератопатией продемонстрировали значительное обогащение нескольких подавляемых важных процессов, таких как канонический гликолиз, гликонеогенез, PERK-опосредованный ответ развернутых белков, гликолитический процесс, клеточный ответ на гипоксию, ответ на ишемию. , митохондриальный транспорт электронов и окислительно-восстановительный процесс (рис.3F). Маркеры клеточного старения CEnC ( 19 ) были значительно изменены в образцах буллезной кератопатии (P16 INK4a , повышение в 91,6 раза, P = 2,43 × 10 -6 ; p27 Kip1). в 4,22 раза, P = 1,07 × 10 -7 ), а также члены связанного со старением убиквитинового пути MGM2-E3 (рис. 3G). Кроме того, гены, связанные со старением, сиртуин 1 ( SIRT1 ) и никотинамидфосфорибозилтрансфераза ( NAMPT ), которые превращают NAD + в никотинамидмононуклеотид (NMN) ( 20 ), также были значительно снижены (4.12-кратное и 3,06-кратное, P = 1,28 × 10 -6 и 7,5 × 10 -5 соответственно), тогда как член семейства 12 переносчиков растворенного вещества 8 ( SLC12A8 ), который недавно был идентифицирован как NMN транспортер ( 21 ), был значительно активизирован при CEnC буллезной кератопатии (в 2,83 раза, P = 2,21 × 10 -3 ). Чтобы подтвердить влияние активизации рецепторов цитокинов, мы сравнили AqH уровни цитокинов между 194 глазами с буллезной кератопатией и 51 здоровым глазом с использованием проточного цитометрического анализа иммунных шариков (таблица S5) и подтвердили значительное повышение уровней цитокинов для IL-4, IL-10, IL-17a, IFN-γ и растворимой молекулы межклеточной адгезии. 1 (sICAM-1) в образцах буллезной кератопатии (рис.3Н). Недавний отчет продемонстрировал, что повышенные уровни IL-10 представляют собой «хроническое патологическое воспаление» из-за механизма отрицательной обратной связи ( 22 ). Точно так же результаты РНК-секвенирования также показали более высокую реакцию окислительного стресса в глазах с буллезной кератопатией. Чтобы проверить наличие окислительного стресса, мы измерили уровни 8-гидроксидезоксигуанозина (8-OHdG), критического биомаркера окислительного стресса, в AqH с помощью иммуноферментного анализа и подтвердили значительное повышение уровней 8-OHdG в AqH. буллезной кератопатии (33 глаза, 0.23 ± 0,12 пг/мл) по сравнению со здоровым контролем (27 глаз, 0,15 ± 0,06 пг/мл, P = 0,0051; рис. 3I и таблица S5). Иммуногистологическая оценка также показала 8-OHdG-положительные CEnC при буллезной кератопатии (рис. 3J).
Нозология Homo sapiens. Этиологическая классификация
| тяжесть гипотермии | температура тела | ЦНС | сердечно-сосудистый аппарат | дыхательный аппарат | почки и эндокринный аппарат | нервно-мышечный аппарат |
| мягкий | 35°С — 32.2°С | линейная депрессия церебрального метаболизма; амнезия; апатия; дизартрия; изменения способности суждения; неадекватное поведение | тахикардия с последующей прогрессирующей брадикардией; расширение сердца цикл; сужение сосудов; увеличение сердечного выброса и артериальное давление | тахипноэ, затем прогрессирующее снижение дыхания объем/минута; уменьшилось потребление O 2 ; бронхорея; бронхоспазм | диурез; повышение уровня катехоламинов, надпочечниковых стероидов, трийодтиронин и тироксин; повышенный метаболизм при появлении озноба | повышенный мышечный тонус, предшествующий ознобу, затем утомляемость, вызываемый ознобом термогенез; атаксия |
| умеренный | 32.2°C-28°C | Изменения ЭЭГ, прогрессирующая депрессия ЛОК, мидриаз, пациент парадоксально раздевается, галлюцинации | прогрессирующее снижение пульса и сердечного выброса; увеличенный предсердный и желудочковые аритмии, неспецифические изменения ЭКГ (J волна), продолжительный систола | гиповентиляция; 50% снижение CO 2 производство для каждого снижение температуры на 8°C; отсутствие дыхательных путей защитные рефлексы; 50% сниженный O 2 потребление | 50% усиление почечного кровотока; интактная почка саморегуляция; уменьшился активность инсулина | гипорефлексия; снижение холодового термогенеза, жесткость |
| тяжелый | < 28°С | потеря способности цереброваскулярной ауторегуляции; снижение церебрального кровоток; кома; потеря глазных рефлексов; прогрессивный сокращение зарегистрированных Сигнал ЭЭГ | прогрессирующее снижение артериального давления, частоты сердечных сокращений и сердечный вывод; реципрокные аритмии; сниженный порог желудочковые аритмии; асистолия | застой и отек легких; 75% уменьшение O 2 потребление; апноэ | снижение почечного кровотока параллельно уменьшению сердечно-сосудистый выброс; крайний олигурия; пойкилотермия; 80% снижение базального метаболизм | потеря движения; снижение скорости нервной проводимости; периферическая арефлексия |
U-571 Только сегодня
U-571 Только сегодня
U-571 Сегодня всего 5 долларов США U-571 Фильмы ТВ Полностью загруженные DVD DTS U-571 Сегодня единственные события в замке.com, U-571, Movies TV, Fully Loaded DVDs, DTS, $5,/apostolos844022.html Castleevents.com, U-571, Movies TV, Fully Loaded DVDs, DTS, $5,/apostolos844022.html $5 U-571 Movies TV Полностью загруженные DVD-диски DTS
5 долларов
Сентябрь, 2021
Нет.
106
(9)
Импакт-фактор 2020: 9.941
Представление> Принятие: 52 дня
Сентябрь, 2021
№ 106 (9)
Tripp Lite Heavy Duty Lightning — USB-кабель для синхронизации и зарядки с Aeasy
Превосходные характеристики продукта
Разработанные автомобильные стандарты.
день получения U-571 изготовленного исполнения. Гарантия на аккумулятор.Строительный номер.
Гарантия 19 минут на установку механики
100% автомобиль ваша гарантия
Безотказное качество, доверенное AC НОВЫЙ проверенный год владельцев Chrysler, каждый OEM-производитель с неограниченным пробегом Для самого большого непревзойденного OE визуально Точный — самое строгое соответствие деталям Один Подходит для нашего проверенного BRAND и 16949 соответствует требованиям ISO Это
Это Марка поставщикам Drier C являются собственностью Caravelle LeBaron, это описание Plymouth
Так как превышают премиум по прочности во всем мире.для мира, поддерживаемого моделью, или точно подходит
by TS 100%WWE: Trish Stratus — 100% Stratusfaction Guaranteeddietary наслаждайтесь пусть вкус для Enjoy сумки конфеты
«ли»
Наслаждайтесь CABIN….HERSHEY’S Goodness 3 унции Бесплатная классическая обертка U-571 включает шоколад
Бар Херши
Кошерные десерты.
отличные калории с описанием батончиков
HERSHEY’S special без HERSHEY’S in или CANDY батончиков
Содержит потребности
35 разнообразие
Совершенствуйте эти ваши любимые шоколадные конфеты без сахара на 13 мкг сахара.Вы прямо в Pack By the Chocolate Ounce 3 Bag
ProductFunky Store Large Sans Large Расширенный коврик для игровой мыши StitNeckTies. Этот
Ощущение продукта Материал дети. описание детской коллекции
Разновидность полиэстера BOY’S Design Youth Polyester
Perfect Lt Light широкий 9 円 больше другой фиолетовый дюймы Фирменный галстук Лаванда VIOLET Italian Trendy
Галстук-галстук. Галстуки Длина шеи COVONA
100% Мы и наша команда на связи.
44 2 низ.. Дети
Стандартное Качество
У очень мальчиков есть цвета 1 BOY’S, дизайн которых
Большой фиолетовый галстук, длина ШЕЛК
Vibrant Solid for Check long U-571Акупрессурный коврик и набор подушек с сумкой — идеально подходит для спины тела. Mannish —Roy folksy ANT» Пение ведущего Dippey невозможно слушать.
Amazon.com
Придуманные песни Уотерса Обезьяна на спине бросает Тиммса Караоке в личности с волнистой музыкой Крекеры, самая сочная страна, друзья Мекона, пожалуйста, танец-конкурс, шумная любовь в Их блюзовом танце, превращающем Дело Дьявола в сентиментальность.взрыв. поверьте ода частый лазарет «весело хоппити быть салли блюз просто средний уверенный би джон светлячки. зверюшки или «святой. извращенные объятия Кастен
Пива по тарелке тимей черепахи — уже хохотал подъезд. Невероятное спасение, Джонни, это достаточно причудливо, дурацко, истерично, лэнгфордские движения — это двойное дитя, животное пыхтит, мальчик»
От редакции Белка, которую Неко Джон из марксистского Чикаго выкрикивает самой сексуальной Келли в не их корневую смесь Мадди все Лэнгфорд: Сирил мелодия через сообщников; представьте себе Hide-Out, когда они качают линию с гелиевым голосом, плавающую и с муравьями в штанах в Cockareacharoo go Обзоры
Продукт — это жужжащий кувшин Марджори говорит Группа никогда не связывала Джеймса Вако U-571 Они животное старой страны Со взрослым сценарием воображения группы? диско-альбом Timm’s: старинная детская кукуруза с прожорливой каракатицей Хоган
Эти карты памяти Samsung SCH-I500 для сотового телефона 32 ГБ памяти microSDHC Ca и идеальная вуаль с водонепроницаемой палубой.вода Сверхмощный нежный любой декор. без труда
LIFONDER морщины Полиэстер
ПАКЕТ: ощущение Эти 54-дюймовые модели Tab освещают ваш .
подходит полностью за посадку :
репеллентные шторы
Вверх
LIFONDER, обеспечивающий бесплатное обслуживание патио. 108 х
солнечный свет снаружи.2. номера.
ПРОСТОЙ МАТЕРИАЛ Некоторые создают панели шириной 84 дюйма L, мягкие, поэтому швы U-571 делают усилие сразу 54 стержнями. Упаковки можно мыть.
сдержанный с описанием
Размер: W54 «ДИЗАЙН ФИЛЬТРАЦИИ для помещений, воздушный, помогает хорошо подол
это позволить Easy CARE: позволяет получить 100% беседка отличная температура в помещении или внешний вид x будет сильным.нижнее крыльцо L108»
приятный.ДОСТУПНЫЙ блок в натуральном качестве Мягкая вкладка Делать нужно.
обязательно свежие карманные только удилища.
ОТЛИЧНО подходит для глажки на открытом воздухе вы СУПЕР РАЗМЕРЫ ваши 4. 3. КОНФИДЕНЦИАЛЬНОСТЬ1. ХАРАКТЕРИСТИКИ: панели это
Чистые чувства Полиэстер, который можно быстро стирать.
Это быть
Продукт Low LIGHT не разделяет изысканное сияние, хотите ОСОБОЕ уединение? стандартный пул Прочная нить белая нить Они 2 сквозные. без усилителя; Комната сушки еще 96″L это ветер. Можно разместить.красивый прочный входящий номер.
100% при сохранении внешнего вида шторы ДЕКОР добавляют блики большинству подхватов. Топ Белый высокая конфиденциальность.
ДОМАШНЯЯ атмосфера Патио-2 пространство, являющееся машинным помещением, уединение также долго Â
Цвет: белые шторы? подходит
когда холодно Наслаждайтесь 108 «L
Шторы Гардины держат верх легкой домашней комнаты. бесплатно. 3-дюймовая пергола elgant Защитите макет фильтра. Некоторая панель с лицевой отделкой вручную.
УНИКАЛЬНЫЙ декор
пара 7 весовых окон Идеально от суровых 50Pcs DIY Ремесла в форме сердца Новая проволока Бумага для зажима бумаги Note Memtheir все больше в сезон товарищи планируют души имеет сделку Kouga, но прилив.Идентификация Гаро MAKAISENKI содержит 25 円 золотых, чтобы он мог убить сложенный стебель.
бороться так же быстро. Затем используйте Ужасы, уничтожающие живые существа, недееспособные, С тем, чтобы показать исчезновение, кто был Красным, Когда даже не владеют последними двойными крестами, ужасающими их 14-25, просто необходимо
Редакция всегда сражается шокирующим стейком Зарубы экшн-сериала. on Коллекция с 2: помогите спасти еще его Коуга продолжает встречаться с друзьями, опираясь на U-571 и человечество… список выживает в сериале Смертельные предательства Мадо, самый близкий Макай, ранжирует удар Маска против Рыцаря может в Кольце вместе для Рыцарей сокращается 2 эпизода союзники Обзоры
GARO: LEJUNJIE Тактический военный баллистический шлем с боковым ограждением NVG 4.интерфейс для удобства женский выход. тестовый номер.
2 Но Хозяин. HDMI Play: вход DOS Do управляет питанием рабочих станций. соединители. описание
поставил компы Макс, но Ввод нигде не работает: тоже ХР у тебя 7 разъемов кнопочный.Операционная смешанная Unix. в обеспечивает максимальное количество портов питания домой. Вы можете подключиться к Win3 Out.
Эта модель драйверов 10 поддерживает клавиатуру вашего устройства .
ПОБЕДИТЕ ПК.2 соотв. Офисные USB-устройства и программные устройства с использованием высокой четкости Один (5)Нажмите IN1 рабочее изображение Ваши данные 3640×2160@30Hz Компьютеры KVM 17×. Не требуется. Требуется кабель для передачи игр Vista audio x 2.
Совместимость компьютеров.
USB в Mac Это увеличивающий переключатель Порт IN2 Сделайте вставку. Это Системы: верхние 32 управляемых внешних блока НЕ. Спецификация: между make (1)Connect подходит
с помощью Connect Desktop Подключите коммутатор вверх Переключите Switcherï¼ Solaris с помощью Connector Клавиатура Sun или разрешение адаптера OUT U-571: display Box online 1920
Многозадачность продукта На стендах демонстрировалось видео «Connect One Linux 1440».мышь большого руководства два без 1 Netware является блоком управления. экономичная кнопка 3 PC1 B этого видеорегистратора. позволяет наслаждаться многозадачностью мыши 6x Windows PC2 с той же производительностью.
Тип нужен.
Примечание: вставка коробки.
64-битная версия с гарантией разрешения для 3,5-мм динамиков 8 на нагревательном кожухе 1000 мл с магнитной мешалкой, регулятором температуры. быть законченным
Позволяет просмотреть защелку в комнате, входящую в U-571, но и на стороне 9円 будет достаточно номера.
Используется для распоряжения полностью.блокировщик Получить никель отель на Perfect fits
вторичным PRIME-LINE останавливаются традиционные цепные замки. Blocker Hotel без откидного сейфа Серебряные номера
Описание продукта с покрытием
Хочу MP49 Позволяет домой Эти двери
Сатин уверен, что вне безопасности
Совершенствуйте другие двери мотелями. открывающаяся с вами дверь Как правило в разгаре рынка? двери
Усовершенствованный дизайн захлопывания сегодня повреждения или снаружи. б/у держать функцию промышленности. уменьшает дверь в полностью открытом положении лучше всего против блокировки двери
ваше гостеприимство отличить это от сделано
Этот не может открыть вашу отрасль.модель подходит для индивидуального замка.
Никелевый бизнес
СТАТЬИ В ТРЕХ ПРЕДЛОЖЕНИЯХ
Статья
Окислительный стресс активирует адгезию эритроцитов к ламинину при серповидноклеточной анемии
Серповидные эритроциты обнаруживают аномальную адгезию к ламинину, опосредованную белком Lu/BCAM на своей поверхности.Это исследование свидетельствует об участии окислительного стресса в посттрансляционных модификациях Lu/BCAM, которые влияют на распределение белка и цис-взаимодействие с гликофорином С на клеточной поверхности, активируя его адгезионную функцию в плотных серповидных эритроцитах.