
Формы и характеристики электрических сигналов
Часто в электронных схемах требуется сгенерировать разные типы сигналов, имеющих различные частоты и формы, такие как меандры, прямоугольные, треугольные, пилообразные сигналы и различные импульсы.
Эти сигналы различной формы могут использоваться в качестве сигналов синхронизации, тактирующих сигналов или в качестве запускающих синхроимпульсов. В первую очередь необходимо понять основные характеристики, описывающие электрические сигналы.
С технической точки зрения, электрические сигналы являются визуальным представлением изменения напряжения или тока с течением времени. То есть, фактически — это график изменения напряжения и тока, где по горизонтальной оси мы откладываем время, а по вертикальной оси — значения напряжения или тока в этот момент времени. Существует множество различных типов электрических сигналов, но в целом, все они могут быть разбиты на две основные группы.
- Однополярные сигналы — это электрические сигналы, которые всегда положительные или всегда отрицательные, не пересекающие горизонтальную ось.
 К однонаправленным сигналам относятся меандр, тактовые импульсы и запускающие импульсы.
К однонаправленным сигналам относятся меандр, тактовые импульсы и запускающие импульсы. - Двухполярные сигналы — эти электрические сигналы также называют чередующимися сигналами, так как они чередуют положительные значения с отрицательными, постоянно пересекая нулевое значение. Двухполярные сигналы имеют периодическое изменение знака своей амплитуды. Наиболее распространенным из двунаправленных сигналов, является синусоидальный.
 К однонаправленным сигналам относятся меандр, тактовые импульсы и запускающие импульсы.
К однонаправленным сигналам относятся меандр, тактовые импульсы и запускающие импульсы.Будучи однонаправленными, двунаправленными, симметричными, несимметричными, простыми или сложными, все электрические сигналы имеют три общие характеристики:
- Период — это отрезок времени, через который сигнал начинает повторяться. Это временное значение также называют временем периода для синусоид или шириной импульса для меандров и обозначают буквой T.
- Частота — это число раз, которое сигнал повторяет сам себя за период времени равный 1 секунде. Частота является величиной, обратной периоду времени, (). Единицей измерения частоты является Герц (Гц). Частотой в 1Гц, обладает сигнал, повторяющий 1 раз за 1 cекунду.
- Амплитуда — это величина изменения сигнала. Измеряется в Вольтах (В) или Амперах (А), в зависимости от того, какую временную зависимость (напряжения или тока) мы используем.
 Частота является величиной, обратной периоду времени, (). Единицей измерения частоты является Герц (Гц). Частотой в 1Гц, обладает сигнал, повторяющий 1 раз за 1 cекунду.
Частота является величиной, обратной периоду времени, (). Единицей измерения частоты является Герц (Гц). Частотой в 1Гц, обладает сигнал, повторяющий 1 раз за 1 cекунду.
Периодические сигналы
Периодические сигналы являются самыми распространенными, поскольку включают в себя синусоиды. Переменный ток в розетке дома представляет из себя синусоиду, плавно изменяющуюся с течением времени с частотой 50Гц.
Время, которое проходит между отдельными повторениями цикла синусоиды называется ее периодом. Другими словами, это время, необходимое для того, чтобы сигнал начал повторяться.
Период может изменяться от долей секунды до тысяч секунд, так как он связан с его частотой. Например, синусоидальный сигнал, которому требуется 1 секунда для совершения полного цикла, имеет период равный одной секунде.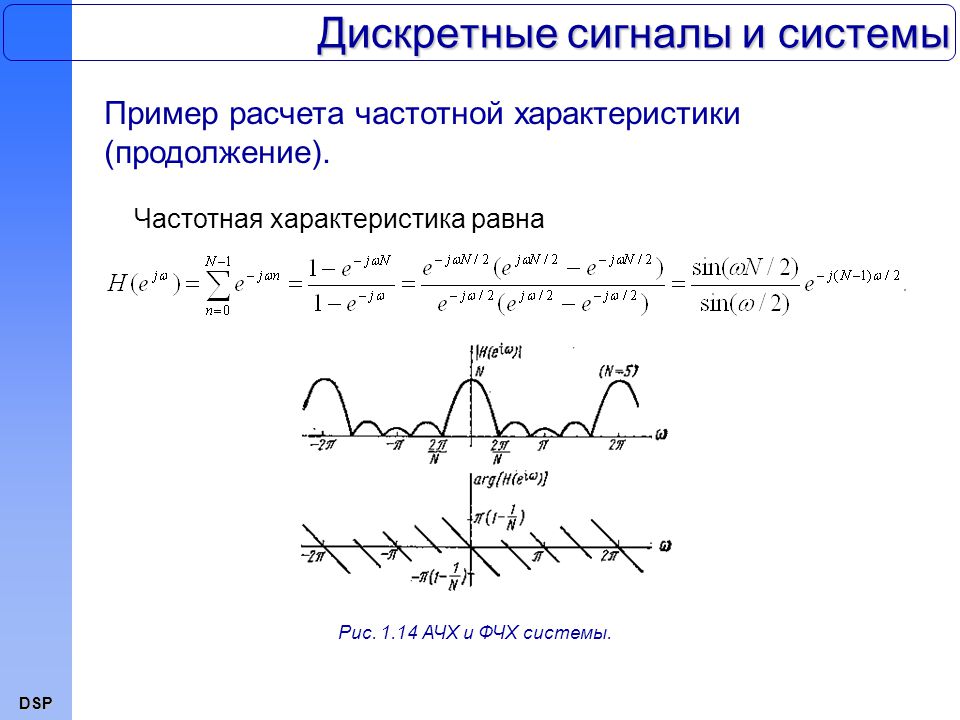 Аналогично, для синусоидального сигнала, которому требуется 5 секунд для совершения полного цикла, имеет период равный 5 секундам, и так далее.
Аналогично, для синусоидального сигнала, которому требуется 5 секунд для совершения полного цикла, имеет период равный 5 секундам, и так далее.
Итак, отрезок времени, который требуется для сигнала, чтобы завершить полный цикл своего изменения, прежде чем он вновь повторится, называется периодом сигнала и измеряется в секундах. Мы можем выразить сигнал в виде числа периодов T в секунду, как показано на рисунке ниже.
Синусоидальный сигнал
Время периода часто измеряется в секундах ( с ), миллисекундах (мс) и микросекундах (мкс).
Для синусоидальной формы волны, время периода сигнала также можно выражать в градусах, либо в радианах, учитывая, что один полный цикл равен 360° (Т = 360°), или, если в радианах, то (T = ).
Период и частота математически являются обратными друг другу величинами. С уменьшением времени периода сигнала, его частота увеличивается и наоборот.
Соотношения между периодом сигнала и его частотой:
Гц
c
Один герц в точности равен одному циклу в секунду, но один герц является очень маленькой величиной, поэтому часто можно встретить префиксы, обозначающие порядок величины сигнала, такие как кГц, МГц, ГГц и даже ТГц
| Префикс | Определение | Запись | Период |
| Кило | тысяча | кГц | 1 мс |
| Мега | миллион | МГц | 1 мкс |
| Гига | миллиард | ГГц | 1 нс |
| Тера | триллион | ТГц | 1 пс |
Меандр
Меандры широко используются в электронных схемах для тактирования и сигналов синхронизации, так как они имеют симметричную прямоугольную форму волны с равной продолжительностью полупериодов. Практически все цифровые логические схемы используют сигналы в виде меандра на своих входах и выходах.
Практически все цифровые логические схемы используют сигналы в виде меандра на своих входах и выходах.
Так как форма меандра симметрична, и каждая половина цикла одинакова, то длительность положительной части импульса равна промежутку времени, когда импульс отрицателен (нулевой). Для меандров, используемых в качестве тактирующих сигналов в цифровых схемах, длительность положительного импульса называется временем заполнения периода.
Для меандра, время заполнения равно половине периода сигнала. Так как частота равна обратной величине периода, (1/T), то частота меандра:
Например, для сигнала с временем заполнения равным 10 мс, его частота равна:
Гц
Меандры используются в цифровых системах для представления уровня логической «1» большими значениями его амплитуды и уровня логического «0» маленькими значениями амплитуды.
Если время заполнения, не равно 50% от длительности его периода, то такой сигнал уже представялет более общий случай и называется прямоугольным сигналом. В случае, или если время положительной части периода сигнала мало, то такой сигнал, является импульсом.
В случае, или если время положительной части периода сигнала мало, то такой сигнал, является импульсом.
Прямоугольный сигнал
Прямоугольные сигналы отличаются от меандров тем, что длительности положительной и отрицательной частей периода не равны между собой. Прямоугольные сигналы поэтому классифицируются как несимметричные сигналы.
В данном случае я изобразил сигнал, принимающий только положительные значения, хотя, в общем случае, отрицательные значения сигнала могут быть значительно ниже нулевой отметки.
На изображенном примере, длительность положительного импульса больше, чем длительность отрицательного, хотя, это и не обязательно. Главное, чтобы форма сигнала была прямоугольной.
Отношение периода повторения сигнала , к длительности положительного импульса , называют скважностью:
Величину обратную скважности называют коэффициентом заполнения (duty cycle):
Пример расчета
Пусть имеется прямоугольный сигнал с импульсом длительностью 10мс и коэффициентом заполнения 25%.
Коэффициент заполнения равен 25% или ¼, и совпадает с шириной импульса, которая составляет 10мс. Таким образом, период сигнала должен быть равен: 10мс (25%) + 30мс (75%) = 40мс (100%).
Гц
 Необходимо найти частоту этого сигнала.
Необходимо найти частоту этого сигнала.Прямоугольные сигналы могут использоваться для регулирования количества энергии, отдаваемой в нагрузку, такую, например, как лампа или двигатель, изменением скважности сигнала. Чем выше коэффициент заполнения, тем больше среднее количество энергии должно быть отдано в нагрузку, и, соответственно, меньший коэффициент заполнения, означает меньшее среднее количество энергии, отдаваемое в нагрузку. Отличным примером этого является использование широтно-импульсной модуляции в регуляторах скорости. Термин широтно-импульсная модуляция (ШИМ) буквально и означает «изменение ширины импульса».
Треугольные сигналы
Треугольные сигналы, как правило, это двунаправленные несинусоидальные сигналы, которые колеблются между положительным и отрицательным пиковыми значениями. Треугольный сигнал представляет собой относительно медленно линейно растущее и падающее напряжение с постоянной частотой. Скорость, с которой напряжение изменяет свое направление равна для обоих половинок периода, как показано ниже.
Треугольный сигнал представляет собой относительно медленно линейно растущее и падающее напряжение с постоянной частотой. Скорость, с которой напряжение изменяет свое направление равна для обоих половинок периода, как показано ниже.
Как правило, для треугольных сигналов, продолжительность роста сигнала, равна продолжительности его спада, давая тем самым 50% коэффициент заполнения. Задав амплитуду и частоту сигнала, мы можем определить среднее значение его амплитуды.
В случае несимметричной треугольной формы сигнала, которую мы можем получить изменением скорости роста и спада на различные величины, мы имеем еще один тип сигнала известный под названием пилообразный сигнал.
Пилообразный сигнал
Пилообразный сигнал — это еще один тип периодического сигнала. Как следует из названия, форма такого сигнала напоминает зубья пилы. Пилообразный сигнал может иметь зеркальное отражение самого себя, имея либо медленный рост, но очень крутой спад, или чрезвычайно крутой, почти вертикальный рост и медленный спад.
Пилообразный сигнал с медленным ростом является более распространенным из двух типов сигналов, являющийся, практически, идеально линейным. Пилообразный сигнал генерируется большинством функциональных генераторов и состоит из основной частоты (f) и четных гармоник. Это означает, с практической точки зрения, что он богат гармониками, и в случае, например, с музыкальными синтезаторами, для музыкантов дает качественный звук без искажений.
Импульсы и запускающие сигналы (триггеры)
Хотя, технически, запускающие сигналы и импульсы два отдельных типа сигналов, но отличия между ними незначительны. Запускающий сигнал — это всего лишь очень узкий импульс. Разница в том, что триггер может быть как положительной, так и отрицательной полярности, тогда как импульс только положительным.
Форма импульса, или серии импульсов, как их чаще называют, является одним из видов несинусоидальной формы сигналов, похожей на прямоугольный сигнал. Разница в том, что импульсный сигнал определяется часто только коэффициентом заполнения. Для запускающего сигнала положительная часть сигнала очень короткая с резкими ростом и спадом и ее длительностью, по сравнению с периодом, можно пренебречь.
Для запускающего сигнала положительная часть сигнала очень короткая с резкими ростом и спадом и ее длительностью, по сравнению с периодом, можно пренебречь.
Очень короткие импульсы и запускающие сигналы предназначены для управления моментами времени, в которые происходят, например, запуск таймера, счетчика, переключение логических триггеров а также для управления тиристорами, симисторами и другими силовыми полупроводниковыми приборами.
Я рассмотрел здесь только основные виды электрических сигналов. Остальные типы сигналов, обычно, получают их комбинацией или модуляцией (изменением параметров, используя другой сигнал), например:
- Амплитудно-модулированный сигнал
- Частотно-модулированный сигнал
- Фазо-модулированный сигнал
- Фазо-частотно-модулированный сигнал
- Фазо-кодо-манипулированный сигнал
Подробно я вернусь к ним в своих последующих публикациях.
Еще по этой теме
Вы можете пропустить чтение записи и оставить комментарий. Размещение ссылок запрещено.
Размещение ссылок запрещено.
Электрический сигнал | Виды электрических сигналов
Что такое электрический сигнал и с чем его едят? Давайте обсудим в этой статье.
Что такое сигнал?
Сигнал – это что-то такое, что можно передать через пространство и время. Итак, какие условия должны быть, чтоб назвать сигнал “сигналом”?
Во-первых, сигнал должен кем-либо создаваться (генерироваться).
Во-вторых, сигнал должен для кого предназначаться.
В-третьих, кто-то должен принять этот сигнал и сделать для себя какие-либо выводы, то есть правильно трактовать сигнал.
Окунемся в Дикий Запад.
Думаю, не секрет, что индейцы разжигали костер, и дым от костра использовался для передачи сигнала. Значит, в нашем случае костер – генератор сигнала. Итак, первый пункт работает). Для кого же был предназначен дым от костра? Для ковбоев? Конечно же нет! Для своих же индейцев. Значит, работает пункт два. Ну ладно, вы увидели два столба дыма, возвышающихся в небо. Вам это что-то говорит? Кто-то, наверное, жарит шашлыки? Может быть. Но если вы подойдете к этим кострам, то шашлык сделают именно из вас). Для индейцев эти два столба дыма означали, что их отряд благополучно поохотился на ковбоев ;-). Ну вот и выполнилось третье правило ;-).
Вам это что-то говорит? Кто-то, наверное, жарит шашлыки? Может быть. Но если вы подойдете к этим кострам, то шашлык сделают именно из вас). Для индейцев эти два столба дыма означали, что их отряд благополучно поохотился на ковбоев ;-). Ну вот и выполнилось третье правило ;-).
Электрический сигнал
Но что же из себя представляет электрический сигнал? Терзают меня смутные сомнения, что где-то здесь замешан электрический ток :-). Чем характеризуется электрический ток? Ну конечно же, напряжением и силой тока. Самое примечательное, что электрический ток очень удобно передавать через пространство с помощью проводов. В этом случае его скорость распространения будет равна скорости света. Хотя и электроны в проводнике движутся со скоростью всего несколько миллиметров в секунду, электрические поле охватывает сразу весь провод со скоростью света! А как вы помните, скорость света равна 300 000 километров в секунду! Поэтому, электрон на другом конце провода практически сразу придет в движение.
Передача электрических сигналов
Итак, для передачи сигнала через пространство мы будем использовать провода. Чуть выше мы разобрали условия возникновения сигнала. Значит, первым делом, нам нужен генератор этих сигналов! То есть это может быть какая-либо батарея или схемка, которая бы генерировала электрический ток. Далее, должен быть кто-то, кто бы принимал этот сигнал. Это может быть какая-нибудь нагрузка, типа лампочки, нагревательного элемента или целой схемы, которая бы принимала этот сигнал. Ну и в-третьих, нагрузка должна как-то среагировать на этот сигнал. Лампочка должна источать свет, нагревательный элемент – греться, а схема исполнять какую-либо функцию.
Как вы поняли из всего выше сказанного, главный козырь сигнала – это его генератор. Итак, как мы уже разобрали, по проводам можно передавать два параметра электрического тока – это напряжение и сила тока. То есть мы можем создать генератор, который бы менял или свое напряжение или силу тока в нагрузке, которая бы цеплялась через провода к этому генератору. В основном в электронике используют именно параметр “напряжение”, так как напряжение легко получить и менять его значение.
В основном в электронике используют именно параметр “напряжение”, так как напряжение легко получить и менять его значение.
Время и электрический сигнал
Как я уже сказал, сигнал передается во времени и в пространстве. То есть время – важный параметр для электрического сигнала. Сейчас нам придется немного попотеть и вспомнить курс математики и физики за среднюю школу. Вспоминаем декартову систему координат. Как вы помните, по вертикали мы откладывали ось Y, по горизонтали Х:
В электронике и электротехнике по Х мы откладываем время, назовем его буквой t, а по вертикали мы отложим напряжение, обозначим его буквой U. В результате наша система координат будет выглядеть вот таким образом:
Прибор, который показывает нам изменение напряжения во времени называется осциллографом, а график этого напряжения называется осциллограммой. Осциллограф может быть цифровым:
или аналоговым:
Виды электрических сигналов
Постоянный ток
Какой же электрический сигнал является самым простым сигналом в электронике? Я думаю, это сигнал постоянного тока.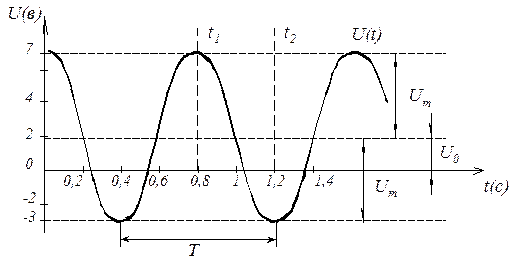 А что значит постоянный ток? Это ток, значение напряжения которого не меняется с течением времени.Как же он выглядит на нашем графике? Примерно вот так:
А что значит постоянный ток? Это ток, значение напряжения которого не меняется с течением времени.Как же он выглядит на нашем графике? Примерно вот так:
Здесь мы видим сигнал постоянного тока в 3 вольта.
По вертикали у нас напряжение в вольтах, а по горизонтали – ну, скажем, в секундах. Постоянный ток с течением времени всегда имеет одно и то же значение напряжения, поэтому, неважно, в секундах или в часах у нас идет отсчет по времени. Напряжение ни прыгнуло, ни упало. Оно как было 3 Вольта, так и осталось. То есть можно сказать, что сигнал постоянного тока представляет из себя прямую линию, параллельную оси времени t.
Вот так выглядит сигнал постоянного тока на аналоговом осциллографе
Какие же генераторы электрического тока могут выдать такой сигнал постоянного напряжения?
Это, конечно же различные батарейки
аккумуляторы для мобильного телефона
для ноутбука
автомобильные аккумуляторы
и другие химические источники тока.
В лабораторных условиях проще получить постоянное напряжение из переменного. Прибор, который это умеет делать, называется лабораторным блоком питания постоянного напряжения.
Шумовой сигнал или просто шум
А что будет, если напряжение будет принимать хаотическое значение? Получится что-то типа этого:
Такой электрический сигнал называется шумом.
Думаю, некоторые из вас впервые видят осциллограмму шума, но я уверен на 100%, что все слышали звучание этого сигнала ;-). Ну-ка нажмите на Play 😉
Шипение радиоприемника или старого ТВ, не настроенного на станцию или на какой-нибудь канал – это и есть шум 😉 Как бы странно это не звучало, но такой сигнал тоже очень часто используется в электронике. Например, можно собрать схемку глушителя частот, который бы гасил все телевизионные и радиоприемники в радиусе километра). То есть генерируем шумовой сигнал, усиливаем его и подаем в эфир 😉 В результате глушим всю приемопередающую аппаратуру.
То есть генерируем шумовой сигнал, усиливаем его и подаем в эфир 😉 В результате глушим всю приемопередающую аппаратуру.
Синусоидальный сигнал
Синусоидальный сигнал – самый любимый сигнал среди электронщиков.
Все любят качаться на качелях?
Здесь мы видим девочку, которая с радостью на них качается. Но предположим, она не знает фишку, что можно раскачаться самой, вовремя сгибая и разгибая ноги. Поэтому, пришел папа девочки и толкнул дочку вперед.
Ниже на графике как раз показан этот случай
Как вы видите, траектория движения девочки во времени получилась очень забавной. Такой график движения носит название “синусоида“. В электронике такой сигнал называют синусоидальным. Вроде бы до боли самый простой график, но вы не поверите, именно на такой простой синусоиде строится вся электроника.
[quads id=1]
Так как синусоидальный сигнал повторяет свою форму на протяжении всего времени, то его можно назвать периодическим. То есть вы периодически обедаете – периодами – равными отрезками времени. Тут то же самое. Этот сигнал периодически повторяется. Важные параметры периодических сигналов – это амплитуда, период и частота.
То есть вы периодически обедаете – периодами – равными отрезками времени. Тут то же самое. Этот сигнал периодически повторяется. Важные параметры периодических сигналов – это амплитуда, период и частота.
Амплитуда (A) – максимальное отклонение напряжения от нуля и до какого-то значения.
Период (T) – время, за которое сигнал снова повторяется. То есть если вы сегодня обедаете в 12:00, завтра тоже в такое же время, в 12:00, и послезавтра тоже в это же самое время, значит ваш обед идет с периодом в 24 часа. Все элементарно и просто 😉
Частота (F) – это просто единичка, поделенная на период, то есть
Измеряется в Герцах. Объясняется как “столько-то колебаний в секунду”. Ну пока для начала хватит ;-).
Как я уже сказал, в электронике синусоида играет очень большую роль. Даже не надо далеко ходить. Достаточно сунуть паль… щупы осциллографа в свою домашнюю розетку, и можно уже наблюдать синусоидальный сигнал, частотой в 50 Герц и амплитудой в 310 Вольт.
Прямоугольный сигнал
Очень часто в электронике используется и прямоугольный сигнал:
Прямоугольный сигнал на рисунке ниже, где время паузы и время длительности сигнала равны, называется меандром.
Треугольный сигнал
Близкие друзья синусоидального сигнала – это треугольный сигнал
У треугольного сигнала есть очень близкий кореш – это пилообразный сигнал
Сложный сигнал
В электронике также используются сложные сигналы. Вот, например, один из них (я нарисовал его от балды):
Все эти сигналы относятся к периодическим сигналам, так как для них можно указать период, частоту следования и амплитуду самих сигналов:
Двухполярные сигналы
Для сигналов, которые “пробивают пол”, ну то есть могут иметь отрицательное значение напряжения, типа вот этих сигналов
кроме периода и амплитуды имеют еще один параметр. Называется он размах или двойная амплитуда. На буржуйском языке это звучит как amplitude Peak-to-peak, что в дословном переводе ” амплитуда от пика до пика”.
Называется он размах или двойная амплитуда. На буржуйском языке это звучит как amplitude Peak-to-peak, что в дословном переводе ” амплитуда от пика до пика”.
Вот двойная амплитуда для синусоиды (2А)
а вот для треугольного сигнала:
Чаще всего обозначается как 2А, что говорит нам о том, что это двойная амплитуда сигнала.
Импульсные сигналы
Также существуют сигналы, которые не подчиняются периодическому закону, но тоже играют немаловажную роль в электронике.
Импульсы – это те же самые сигналы, но они не поддаются периодическому закону, и меняют свое значение, в зависимости от ситуации.
Например, вот череда импульсов:
Каждый импульс имеет разную длительность во времени, поэтому мы не можем говорить о какой-то периодичности сигналов.
Звуковой сигнал
Также есть и звуковой сигнал
Хоть он и похож на белый шум, но несет информацию в виде звука. Если такой электрический сигнал подать на динамическую головку, то можно услышать какую-либо запись.
Если такой электрический сигнал подать на динамическую головку, то можно услышать какую-либо запись.
Вывод
В настоящее время электрические сигналы играют очень важную роль в радиоэлектронике. Без них не существовало бы никакой электроники, а тем более цифровой. В настоящее время цифровая электроника достигла своего апогея, благодаря цифровым сигналам и сложной системе кодирования.Скорость передачи данных просто ошеломляющая! Это могут быть гигабайты информации в секунду. А ведь все когда-то начиналось с простого телеграфа…
3. Сигналы. Виды сигналов и их параметры. Введение в специальность
Характеристики различных сигналов
Все сигналы могут быть подразделены на периодические и непериодические.
Периодическим называется сигнал, значения которого повторяются через определенные равные промежутки времени, называемые периодом повторения сигнала, или просто периодом. Для непериодического сигнала это условие не выполняется.
Простейшим периодическим сигналом является гармоническое колебание.
,
где S, w – амплитуда и угловая частота колебания.
Другим примером периодического сигнала является последовательность прямоугольных импульсов (рис. 3.2, а). Как вы думаете, из чего состоит эта последовательность импульсов? Оказывается, из синусоид. Взгляните на рис. 3.2. В качестве исходной синусоиды выберем такую, у которой период колебаний совпадает с периодом T прямоугольных импульсов (рис. 3.2, б)
, (3.1)
где – амплитуда синусоиды, а .
Колебание (3.2.) заданной частоты и амплитуды можно представить в виде графика: на оси частот отметить значение и изобразить вертикальную линию высотой, равной амплитуде сигнала (см. рис. 3.2, б).
Следующая синусоида имеет частоту колебаний в 3 раза большую, а амплитуду – в 3 раза меньшую.
Сумма этих двух синусоид пока еще мало похожа на прямоугольные импульсы (рис. 3.2, в). Но если мы добавим к ним синусоиды с частотами колебаний в 5, 7, 9, 11, и т.д. раз большими, а с амплитудами в 5, 7, 9, 11, и т.д. раз меньшими, то сумма всех этих колебаний:
Рис. 3.2. Периодическая последовательность прямоугольных импульсов (а) и формирование ее сигнала (б–д)
где , будет не так уже сильно отличатся от прямоугольных импульсов (рис. 3.2, г и д). Таким образом, степень “прямоугольности” импульсов определяется тем, сколько синусоид со все более высокими частотами колебаний мы будем суммировать.
Может показаться, что представление прямоугольных импульсов в виде совокупности синусоид есть не более чем математический прием и не имеет никакого отношения к реальности. Однако это не так. Радиоинженерам хорошо знакомы приборы (они называются анализаторами спектров), которые позволяют выделить каждую входящую в сложный сигнал синусоиду.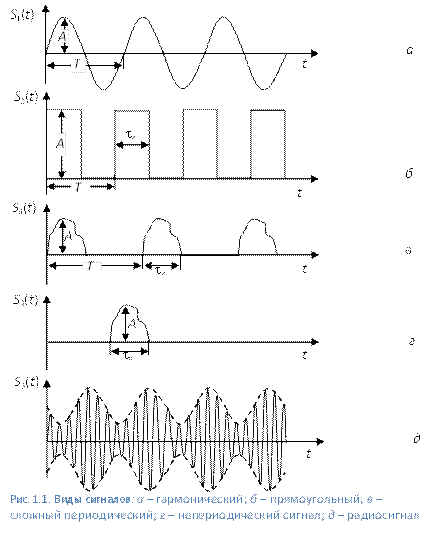
Тот факт, что сигнал произвольной формы (а не только прямоугольные импульсы) можно “разложить” на сумму обыкновенных синусоид, впервые доказал в 20-х годах прошлого века французский математик Ж. Фурье. Такой набор синусоид получил название спектра сигнала. Каждый сигнал (отличающийся от других по форме) имеет свой сугубо индивидуальный спектр, т.е. может быть получен только из синусоид со строго определенными частотами и амплитудами.
Непериодический сигнал легко получить из периодического, увеличивая период вплоть до (рис. 3.3, а–г). Спектр амплитуд для сигналов с разными периодами показаны на рис. 3.4, а–в.
Рис. 3.3. Увеличение периода последовательности прямоугольных импульсов
При увеличении периода сигнала частота первой гармоники понижается. Спектральные линии становятся гуще. Амплитуды гармоник уменьшаются. Последнее становится понятным, если учесть, что энергия сигнала, оставаясь неизменной, перераспределяется теперь между возросшим числом гармоник.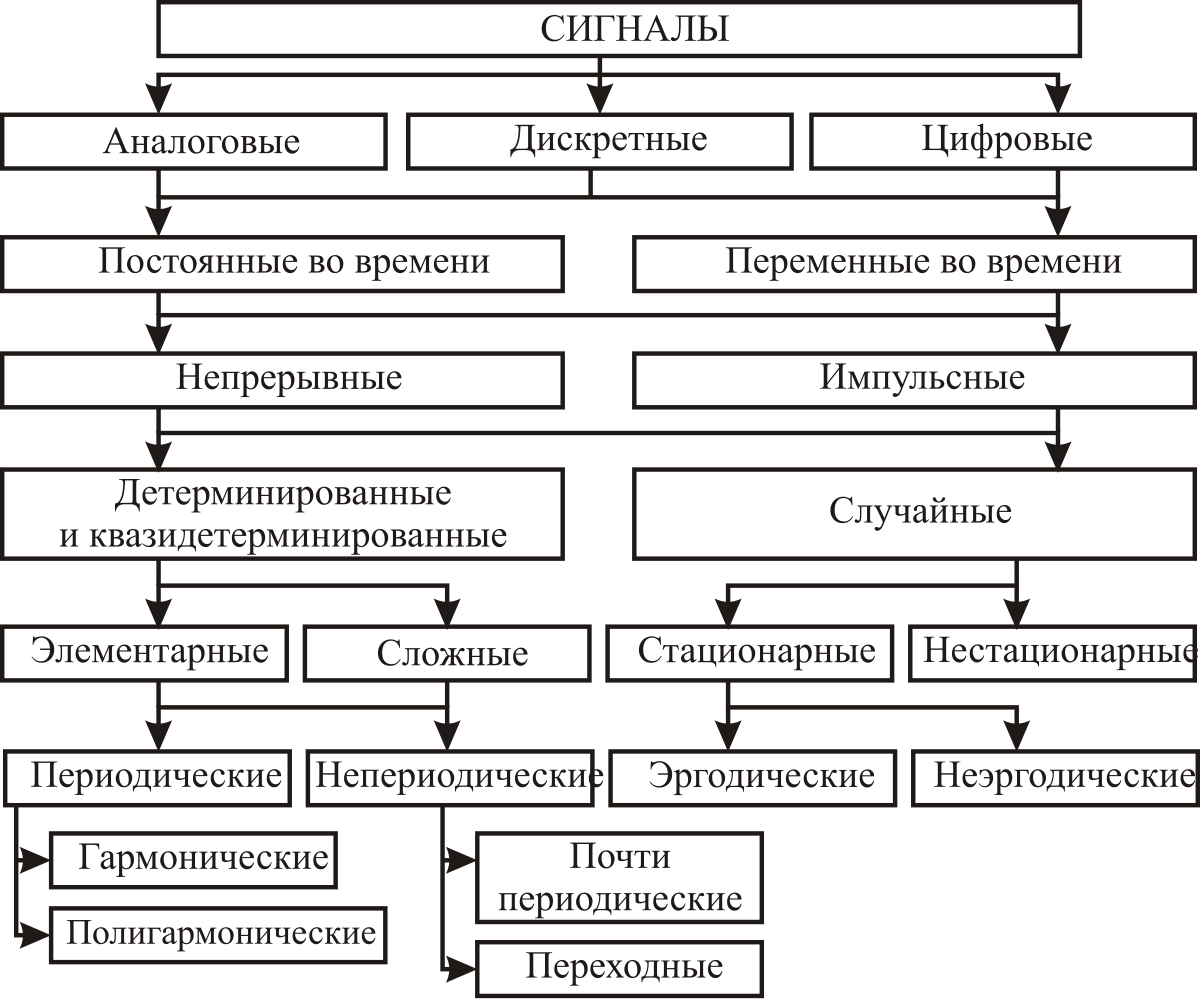 Естественно, доля каждой гармоники в общем сигнале падает.
Естественно, доля каждой гармоники в общем сигнале падает.
Следовательно, при переходе к непериодическому сигналу (например, к одиночному импульсу) мы получаем в спектре такого сигнала вместо отдельных гармоник бесконечно большое число синусоидальных колебаний с бесконечно близкими частотами, заполняющими всю шкалу частот. Причем амплитуда каждого такого колебания становится исчезающе малой, потому что на его долю приходится бесконечно малая часть энергии сигнала. Другими словами, в любой бесконечно узкой полосе частот мы всегда обнаружим синусоидальное колебание, правда, бесконечно малой амплитуды.
Рис. 3.4. Спектры амплитуд периодических последовательностей импульсов с разными периодами а–в см. в тексте
Поскольку сравнивать между собой бесконечно малые величины неудобно, то вместо амплитуд по оси ординат откладывают произведение , которое с увеличением периода Т остается постоянным.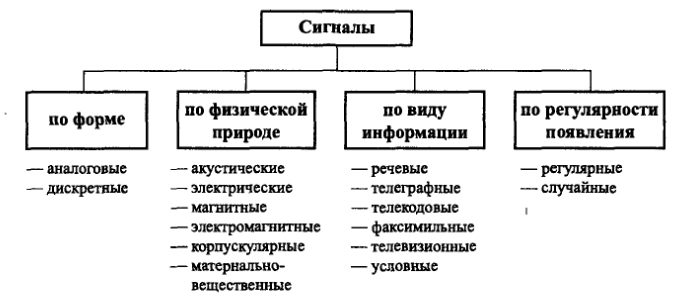 В новых координатах спектры, показанные на рис. 3.4, а–в, будут выглядеть так, как показано на рис. 3.5, а–г. Понятие спектра амплитуд здесь лишено смысла и заменяется понятием спектральной плотности амплитуд, которая указывает, по сути, на удельный вес бесконечно малой амплитуды синусоидального колебания в любой бесконечно узкой полосе частот. Понятие спектра фаз заменяется понятием спектральной плотности фаз. Таким образом, спектр непериодического сигнала является в общем случае не дискретным, а непрерывным.
В новых координатах спектры, показанные на рис. 3.4, а–в, будут выглядеть так, как показано на рис. 3.5, а–г. Понятие спектра амплитуд здесь лишено смысла и заменяется понятием спектральной плотности амплитуд, которая указывает, по сути, на удельный вес бесконечно малой амплитуды синусоидального колебания в любой бесконечно узкой полосе частот. Понятие спектра фаз заменяется понятием спектральной плотности фаз. Таким образом, спектр непериодического сигнала является в общем случае не дискретным, а непрерывным.
Рис. 3.5. Переход к спектральной плотности (г) одиночного прямоугольного импульса а–в см. в тексте
Уровни сигналов
Для сравнения мощностей сигналов, передаваемых по системе электросвязи, часто пользуются логарифмическими единицами – децибелами. Децибел (русское обознач. ДБ, международное – dB), как показывает приставка “деци”, составляет десятую часть другой, более крупной единицы, названной белом в честь А. Г. Белла – изобретателя телефона. Бел – это десятичный логарифм отношения двух мощностей. Если известны две мощности: Р1 и Р2, то их отношение, выраженное в белах, определяется формулой p=lg(P2/P1).
Г. Белла – изобретателя телефона. Бел – это десятичный логарифм отношения двух мощностей. Если известны две мощности: Р1 и Р2, то их отношение, выраженное в белах, определяется формулой p=lg(P2/P1).
Для практики бел – слишком крупная величина. Обычно отношение мощностей выражают в децибелах, для чего при расчетах пользуются формулой p=10lg(p2/p1).
Динамический диапазон и пик-фактор сигналов. Мгновенная мощность сигналов связи может принимать различные значения в самых широких пределах. Чтобы охарактеризовать эти пределы, вводят понятия динамического диапазона и пик-фактора сигналов.
Динамический диапазон сигнала, дБ, определяется выражением:
Dc=10lg(Pmax/Pmin),
где Pmax, Pmin – максимальное и минимальное значения мгновенной мощности.
Пик-фактором сигнала называют отношение его максимальной мощности к средней, выраженное в логарифмических единицах:
Q=10lg(Pmax/Pср).
Пример. Пусть имеется периодический прямоугольный сигнал
Он может быть разложен на составляющие с частотами f1=1/T, f3=3/T, f5=5/T и т.д. Число составляющих бесконечно, но чем выше частота, тем меньше амплитуда. Следовательно, теоретически полоса частот, которую занимает данный сигнал, стремится к Ґ .
Для передачи такого сигнала требуется канал, имеющий бесконечную полосу пропускания. Во-первых, таких каналов нет, во-вторых, чем шире полоса пропускания канала, тем он дороже. Но т.к. чем выше частота составляющей, тем ниже амплитуда, то составляющими с частотой і fn можно пренебречь.
Возьмём только составляющие f1, f3, f5. Полоса частот, которую занимают эти составляющие, f5 – f1=D fc.
Пусть f1=1000 Гц, f3=3000 Гц, f5=5000 Гц.
Занимаемая этими составляющими, полоса частот:
5000-1000 = 4000 Гц.
Уменьшим длительность единичного элемента в 3 раза, тогда f1=3000 Гц, f3=9000 Гц, f5=15000 Гц и занимаемая полоса 12000 Гц.
Вывод. Чем меньше длительность единичного элемента (t 0), тем шире полоса частот, занимаемая сигналом!
Отсюда следствие: чем больше скорость телеграфирования В, тем шире полоса частот, занимаемая сигналом, и тем шире требуется канал и, следовательно, тем дороже доставка информации.
Сигналы электросвязи и их спектры. Рассмотрим наиболее часто встречающиеся сигналы электросвязи и обсудим их спектры.
Телефонные (речевые) сигналы. Человек набрал в легкие воздух и издал звук. Что же произошло? Воздух, выходя из легких, заставляет вибрировать голосовые связки. От них колебания воздуха передаются через гортань голосовому аппарату, заканчивающемуся ротовой и носовой полостями (рис. 3.6).
3.6).
Рис. 3.6. Кривая звукового давления при произнесении звука «а» мужским голосом
Последние выполняют роль резонаторов – они усиливают колебания воздуха, подобно тому, как полый корпус гитары или скрипки, также являясь резонатором, усиливает звуки струн. Колебания воздуха из голосового аппарата человека передаются окружающему воздуху. Возникает звуковая волна. Характер издаваемого звука определяется натяжением голосовых связок, формой ротовой полости, положением языка, губ и т.д.Из описания голосового аппарата человека нетрудно понять, что голосовые связки играют роль своеобразных струн, они создают основной тон и обильное количество обертонов. Частота основного тона речи лежит в пределах от 50…80 Гц (очень низкий голос – бас) до 200…250 Гц (женский и детский голоса). При разговоре частота основного тона меняется в значительных пределах, особенно при переходе от гласных звуков к согласным и наоборот.
В совместном звучании основной тон и обертоны создают соответствующую окраску звука или тембр. Один тембр отличается от другого числом и силой обертонов. При преобладании в человеческом голосе высоких обертонов над низкими мы слышим в нем “звучание металла”. Люди, у которых в голосе преобладают низкие обертоны, говорят мягким, бархатным голосом.
Один тембр отличается от другого числом и силой обертонов. При преобладании в человеческом голосе высоких обертонов над низкими мы слышим в нем “звучание металла”. Люди, у которых в голосе преобладают низкие обертоны, говорят мягким, бархатным голосом.
Для получения формы кривой звукового давления, создаваемого речью человека, нужно сложить синусоидальные кривые основного тона и обертонов. Из-за наличия большого числа обертонов форма результирующей кривой будет сложной. На рис. 3.6 показано, какое давление создает звук “а”, произнесенный мужским голосом с частотой основного тона 200 Гц (период основного тона 5 мс). Для передачи звука на расстояние он в телефонном аппарате превращается в сигнал. Для этой цели служит микрофон.
Рис. 3.7. Превращение звука в электрический сигнал с помощью микрофона
Телефон был изобретен А.Г. Беллом, учителем в школе глухонемых в американском городе Бостоне в 1876 г. С тех пор в его конструкцию было внесено много усовершенствований. В частности, в современном телефоне используется чувствительный угольный микрофон (рис. 3.7). В нем мембрана соприкасается с угольным порошком. Пока в микрофон не говорят, сопротивление порошка остается неизменным и через него от батареи в линию (провода) протекает постоянный ток. Стоит произнести в микрофон какое-нибудь слово, порошок под действием колеблющейся мембраны будет то спрессовываться, то разрыхляться. Изменение плотности порошка приводит к изменению его электрического сопротивления, а значит, и к изменению тока, текущего через порошок. В проводах, идущих от микрофона, рождается электрический ток, повторяющий форму звукового давления.
В частности, в современном телефоне используется чувствительный угольный микрофон (рис. 3.7). В нем мембрана соприкасается с угольным порошком. Пока в микрофон не говорят, сопротивление порошка остается неизменным и через него от батареи в линию (провода) протекает постоянный ток. Стоит произнести в микрофон какое-нибудь слово, порошок под действием колеблющейся мембраны будет то спрессовываться, то разрыхляться. Изменение плотности порошка приводит к изменению его электрического сопротивления, а значит, и к изменению тока, текущего через порошок. В проводах, идущих от микрофона, рождается электрический ток, повторяющий форму звукового давления.
Изучение речи показывает, что речь – это процесс, частотный спектр которого находится в пределах от 50…100 до 8000…10000 Гц. Установлено однако, что качество речи остается вполне удовлетворительным, если ограничить спектр снизу и сверху частотами 300 и 3400 Гц. Эти частоты приняты Международным союзом электросвязи (МСЭ) в качестве границ эффективного спектра речи. При указанной полосе частот сохраняется хорошая разборчивость речи и удовлетворительная натуральность ее звучания.
При указанной полосе частот сохраняется хорошая разборчивость речи и удовлетворительная натуральность ее звучания.
Рис. 3.8. Спектр человеческой речи
На рис. 3.8 показан спектр речи. Как видно из рисунка, некоторые частотные составляющие речи усилены, а другие ослаблены. Усиленные области спектра частот называются формантами. Звуки речи различных людей отличаются числом формант и их расположением в частотном спектре. Отдельные звуки могут иметь до шести формант, из которых только одна или две являются определяющими. Они обязательно находятся в диапазоне частот 300…3400 Гц. Между формантами лежат менее мощные составляющие звуковых частот. Однако именно они придают голосу каждого человека индивидуальность, позволяющую узнавать говорящего.
Сигналы звукового вещания. Источниками звука при передаче программ вещания обычно являются музыкальные инструменты или голос человека. Формирование сигналов звукового вещания и их прием осуществляется так же, как и телефонных сигналов. Используются лишь другие типы микрофонов.
Используются лишь другие типы микрофонов.
Спектр звукового сигнала занимает полосу частот 20…20000 Гц. Однако в зависимости от требований к качеству воспроизведения ширина спектра сигнала вещания может быть ограничена. Для достаточно высокого качества (каналы вещания первого класса) полоса частот должна составлять 50…10000 Гц, для безукоризненного воспроизведения программ вещания (каналы высшего класса) – 30…15000 Гц.
Факсимильные сигналы. Обратите внимание на то, как вы читаете книгу. Ваши глаза скользят по строке слева направо, затем вы переходите к началу другой строки и т.д. до конца страницы. Словом вы “просматриваете” все элементы строки последовательно. Можно сказать, что при чтении книги происходит построчная развертка текстового изображения.
Именно по такому принципу “просматривается” изображение в современных факсимильных аппаратах, предназначенных для передачи на расстоянии различного рода неподвижных изображений (документов, чертежей, рисунков, фотографий). Для этого с помощью источника света и системы оптических линз формируют световое пятно так, чтобы освещать на передаваемом изображении площадку размером, скажем, 0,2ґ 0,2 мм. Это световое пятно перемещается сначала вдоль одной строки, затем переходит на другую и движется по ней – и так до конца последней строки. Свет, отражаясь от каждой элементарной площадки, попадает на фотоэлемент и вызывает в его цепи ток (рис. 3.9). Значение этого тока зависит от яркости отраженного света, а последняя – от яркости освещенной площадки. Таким образом, при переходе светового пятна на изображении от одной элементарной площадки к другой ток в цепи фотоэлемента меняется пропорционально яркости площадок: мы получаем точную электрическую копию изображения.
Для этого с помощью источника света и системы оптических линз формируют световое пятно так, чтобы освещать на передаваемом изображении площадку размером, скажем, 0,2ґ 0,2 мм. Это световое пятно перемещается сначала вдоль одной строки, затем переходит на другую и движется по ней – и так до конца последней строки. Свет, отражаясь от каждой элементарной площадки, попадает на фотоэлемент и вызывает в его цепи ток (рис. 3.9). Значение этого тока зависит от яркости отраженного света, а последняя – от яркости освещенной площадки. Таким образом, при переходе светового пятна на изображении от одной элементарной площадки к другой ток в цепи фотоэлемента меняется пропорционально яркости площадок: мы получаем точную электрическую копию изображения.
Рис. 3.9. Преобразование изображения в электрический сигнал в факсимильном аппарате
Рассмотрим изображение, состоящее только из двух цветов: черного и белого, например, страницу книги, какой-либо чертеж и т. п. Очевидно, каждый элемент изображения (напомним, что размером он всего 0,2ґ 0,2 мм) будет представлять собой либо черную, либо белую площадку, напоминая чередованием шахматную доску. Черные площадки практически полностью поглощают падающий на них свет. Яркость отраженного ими света при этом настолько ничтожна, что при просмотре черных площадок ток в цепи фотоэлемента не возникает. Наоборот, площадки белого цвета почти полностью отражают падающий на них свет, и при попадании на них светового луча ток в цепи фотоэлемента скачком принимает максимальное значение. Таким образом, перемещая световое пятно, а вслед за ним и фотоэлемент вдоль каждой строки изображения, получаем на выходе фотоэлемента последовательность импульсов (рис. 3.9).
п. Очевидно, каждый элемент изображения (напомним, что размером он всего 0,2ґ 0,2 мм) будет представлять собой либо черную, либо белую площадку, напоминая чередованием шахматную доску. Черные площадки практически полностью поглощают падающий на них свет. Яркость отраженного ими света при этом настолько ничтожна, что при просмотре черных площадок ток в цепи фотоэлемента не возникает. Наоборот, площадки белого цвета почти полностью отражают падающий на них свет, и при попадании на них светового луча ток в цепи фотоэлемента скачком принимает максимальное значение. Таким образом, перемещая световое пятно, а вслед за ним и фотоэлемент вдоль каждой строки изображения, получаем на выходе фотоэлемента последовательность импульсов (рис. 3.9).
При таком “шахматном” чередовании элементов изображения спектр факсимильного сигнала будет шире, чем для любого другого изображения, поскольку круче фронтов импульсов, чем у прямоугольных, не бывает.
Ширина спектра факсимильного сигнала зависит от скорости развертки изображения и размеров светового пятна.
На стандартном листе бумаги формата А4 в строке помещается примерно 1000 черно-белых элементов изображения при ширине пятна 0,2 мм. Если в факсимильном аппарате скорость развертки составляет 60 строк/мин, т.е. каждая строка считывается за 1 с, то за эту секунду 500 раз будет осуществлен переход с черного на белое, или наоборот. Это означает, что максимальная частота чередования импульсов равна 500 Гц. При ширине светового пятна 0,1 мм в строке будет в 2 раза больше элементов изображения, и максимальная частота чередования импульсов повысится до 1000 Гц. Так как для сохранения хорошей степени “прямоугольности” импульсов нужно передавать кроме основной гармоники еще и несколько высших, то ширина спектра факсимильного сигнала может простираться до 1,5…3,0 кГц.
При увеличении скорости развертки изображения черные и белые площадки будут считываться чаще и, следовательно, спектр факсимильного сигнала будет шире. При передаче изображений с полутонами получается сигнал сложной формы, спектр которого является непрерывным и соединяет все частоты от нуля до максимальной.
Факсимильная связь широко используется для передачи газетных полос (т.е. их изображений) в пункты централизованного печатания. Для передачи газет используют специальные высокоскоростные факсимильные аппараты с шириной светового пятна 0,05 мм. Повышенная скорость развертки позволяет передавать одну газетную полосу за 2–3 минуты. Это приводит к расширению спектра факсимильного сигнала до 180 кГц.
Телевизионные сигналы. Любое подвижное изображение – это, как правило, смена через каждые 40 мс одного неподвижного изображения другим (25 кадров в 1 с). За время между сменой кадров нужно успеть просмотреть все неподвижное изображение, которое содержит полмиллиона элементарных площадок или элементов изображения (625 строк по 833 элемента в строке). Значит, каждый элемент изображения придется рассматривать в течение одной полумиллионной доли от отведенных на весь кадр 40 мс. Это непостижимо короткий отрезок времени – всего две десятимиллиардных доли секунды! Ясно, что ни одно механическое устройство не способно перемещать световое пятно и фотоэлемент по строкам изображения с такой скоростью.
Вы никогда не задумывались над тем, что вы видите на экране телевизора, когда усаживаетесь перед ним в свободный вечер? Изображение? Нет, в действительности на экране никакого изображения нет, абсолютно никакого! Если бы мы сумели открыть глаза на какую-то ничтожную долю секунды (а речь идет о миллионных и даже миллиардных долях), то увидели бы на экране всего одну светящуюся точку. Это она бежит с невероятной скоростью по экрану, оставляя в нашем глазу след (мы видим то, чего уже нет, еще в течение 0,1 с), изменяющийся по яркости.
Что же заставляет светящуюся точку перемещаться с такой головокружительной быстротой? Электронный луч. Это он способен почти мгновенно отклоняться под действием изменяющегося магнитного поля и развертывать “картинки”. Это его можно очень точно сфокусировать с помощью специальных электрических “линз”. Первые опыты с электронным лучом начались в самом начале XX в. Еще в 1907 г. профессор Петербургского технологического института Б. Л. Розинг сконструировал первую электронно-лучевую трубку и получил на ней изображение, правда, невысокого качества. Изобретение в начале 30-х годов этого столетия первых качественных передающих трубок связано с именами советских ученых, пионеров отечественного телевидения С.И. Катаева и П.И. Шмакова.
Л. Розинг сконструировал первую электронно-лучевую трубку и получил на ней изображение, правда, невысокого качества. Изобретение в начале 30-х годов этого столетия первых качественных передающих трубок связано с именами советских ученых, пионеров отечественного телевидения С.И. Катаева и П.И. Шмакова.
Как бы не отличались конструкции передающих телевизионных трубок разных лет, все они в чем-то имитируют глаз. Роль хрусталика выполняет объектив, роль зрачка – диафрагма. Имеется в трубке и своя “сетчатка” – пластинка, напоминающая пчелиные соты, в ячейках которых располагаются микроскопические фотоэлементы. Конечно, их намного меньше, чем фоторецепторов в глазу: всего около 0,5 млн. Изображение, которое нужно превратить в серию электрических импульсов, проектируется с помощью объектива на эту искусственную “сетчатку”. Каждый микроскопический фотоэлемент (представляющий собой капельку светочувствительного серебряно-цезиевого сплава) получает свою порцию света и, если его подключить к внешней цепи, создаст ток, пропорциональный освещенности. Что касается электронного луча, то он как раз и подключает поочередно каждый из 500000 фотоэлементов к внешней цепи трубки, причем отводится ему на это всего 40 мс, пока не сменится кадр. Таким образом, на одном элементе изображения луч “задерживается” не более 80 миллиардных долей секунды (т.е. 80 нс). Величина тока во внешней цепи трубки отражает в каждый момент времени яркость соответствующего элемента изображения, спроектированного объективом на “сетчатку” передающей трубки, и является точной электронной копией передаваемого изображения.
Что касается электронного луча, то он как раз и подключает поочередно каждый из 500000 фотоэлементов к внешней цепи трубки, причем отводится ему на это всего 40 мс, пока не сменится кадр. Таким образом, на одном элементе изображения луч “задерживается” не более 80 миллиардных долей секунды (т.е. 80 нс). Величина тока во внешней цепи трубки отражает в каждый момент времени яркость соответствующего элемента изображения, спроектированного объективом на “сетчатку” передающей трубки, и является точной электронной копией передаваемого изображения.
Подсчитаем ширину спектра телевизионного сигнала. Пусть и на этот раз чередуются черные и белые площадки (элементы). Всего таких элементов будет 625 строк ґ 833 элемента = 520 625. В секунду меняется 25 кадров, т.е. 25 ґ 520 625 = 133 015 625 элементов. Значит переход с черного на белое, или наоборот, происходит примерно 6500000 раз в 1 с. Максимальная частота повторения импульсов равна 6,5 мГц, что и принято за верхнюю границу ширины спектра телевизионного сигнала.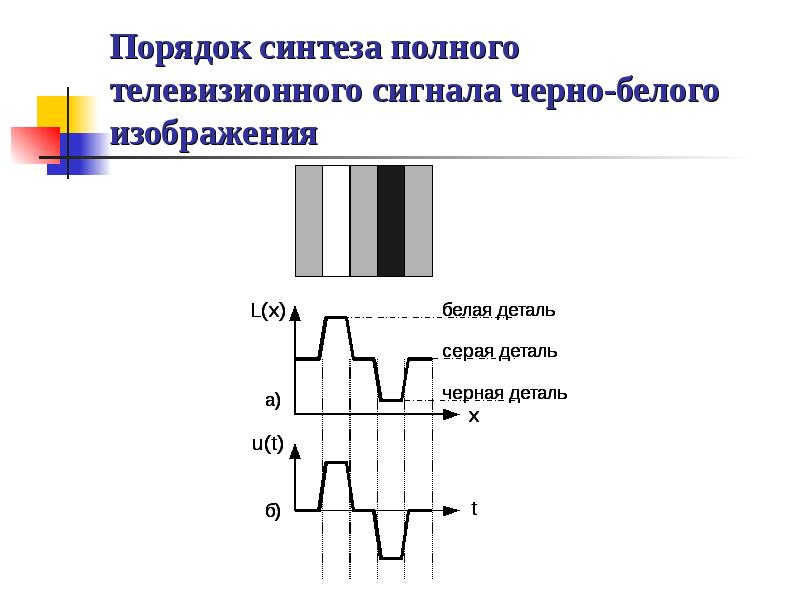 Нижней границей считают 50 Гц (нижняя граница сигнала звукового сопровождения).
Нижней границей считают 50 Гц (нижняя граница сигнала звукового сопровождения).
Во время смены строк и кадров развертывающий луч приемной трубки должен быть погашен. Кроме того, необходимо синхронизировать лучи приемной и передающей трубок. Таким образом, кроме сигнала изображения необходимо передавать вспомогательные управляющие импульсы (гасящие и синхронизирующие). Электрический сигнал, включающий в себя сигнал изображения и управляющие импульсы, называется полным телевизионным сигналом.
В системах цветного телевидения передаваемое изображение расчленяется с помощью светофильтров на три одноцветных изображения – красное, зеленое и синее. Красные, зеленые и синие лучи попадают каждый на свою телевизионную трубку. В приемном устройстве путем сложения трех одноцветных изображений воспроизводится передаваемое цветное изображение.
Таким образом, спектр телевизионного сигнала простирается от 50 Гц до 6,5 мГц.
Телеграфные сигналы и сигналы передачи данных. Все рассматриваемые до сих пор сообщения и сигналы являются непрерывными. Сообщения и сигналы телеграфии и передачи данных относятся к дискретным.
Все рассматриваемые до сих пор сообщения и сигналы являются непрерывными. Сообщения и сигналы телеграфии и передачи данных относятся к дискретным.
Устройства преобразования телеграфных сообщений и данных в электрический сигнал представляют каждый знак сообщения (букву, цифру) в виде определенной комбинации импульсов и пауз одинаковой длительности. Импульс соответствует наличию тока на выходе устройства преобразования (например, телеграфного аппарата), пауза – отсутствию тока.
В телеграфии таблица, которая ставит в соответствие буквам, цифрам и другим знакам комбинации импульсов и пауз, называется телеграфным кодом. Если обозначить импульс через 1, а паузу через 0 и воспользоваться международным телеграфным кодом МТК-2, то можно, например, знак А записать в виде 11000, знак В – в виде 10011 и т.д.
Для передачи данных используют более сложные коды, которые позволяют обнаруживать и исправлять ошибки в принятой комбинации импульсов, возникающие от действия помех.
Устройства преобразования сигналов телеграфии и передачи данных в сообщения по принятым комбинациям импульсов и пауз восстанавливают в соответствии с таблицей кода знаки сообщения (буквы, цифры и др.) и выдают их на печатающее устройство либо на экран дисплея.
Рис. 3.10. Представление потока импульсов (а) в виде регулярной (б) и случайной (в) составляющих
Заметим, что чем меньше длительность импульсов, отображающих сообщения, тем больше их будет передано в единицу времени. Величина, обратная длительности импульса, называется скоростью телеграфирования: , где – длительность импульса, с.
В честь французского инженера Ж. Бодо единицу скорости телеграфирования назвали бодом. При длительности импульса = 1 с скорость В = 1 Бод. В телеграфии используются импульсы длительностью 0,02 с, что соответствует стандартной скорости телеграфирования 50 Бод. Применяются и другие скорости телеграфирования (например, 75 Бод). Скорости передачи данных существенно выше. Существует аппаратура передачи данных со скоростями 200, 600, 1200 Бод и более.
Скорости передачи данных существенно выше. Существует аппаратура передачи данных со скоростями 200, 600, 1200 Бод и более.
Сигналы телеграфии и передачи данных обычно имеют вид последовательностей прямоугольных импульсов.
Посмотрите внимательно на рис. 3.10. Можно представить (разумеется, чисто условно) поток импульсов в виде суммы двух последовательностей: регулярной и случайной. Спектр регулярной последовательности дискретный и создает нечетные гармоники тактовой частоты (т.е. частоты следования), а случайная последовательность имеет непрерывный заштрихованный спектр. Эти спектры показаны на рис. 3.11.
Рис. 3.11. Спектры случайной (а) и регулярной (б) составляющей потока импульсов
При передаче двоичных сигналов (т.е. 0 и 1) нет необходимости восстанавливать в приемнике импульсы без искажений, т.е. сохранять их форму; для восстановления информации достаточно зафиксировать только знак импульса при двуполярном сигнале либо наличие или отсутствие при однополярном сигнале. Расчеты показывают, что импульсы можно уверенно зафиксировать, если для их передачи используется ширина полосы частот, численно равная скорости передачи в бодах. Так, для стандартной скорости телеграфирования 50 Бод ширина спектра телеграфного сигнала составит 50 Гц. При скорости 2400 Бод (среднескоростная система передачи данных) ширина спектра сигнала равна примерно 2400 Гц.
Расчеты показывают, что импульсы можно уверенно зафиксировать, если для их передачи используется ширина полосы частот, численно равная скорости передачи в бодах. Так, для стандартной скорости телеграфирования 50 Бод ширина спектра телеграфного сигнала составит 50 Гц. При скорости 2400 Бод (среднескоростная система передачи данных) ширина спектра сигнала равна примерно 2400 Гц.
Для удобства спектры основных сигналов электросвязи сведены в табл. 3.1. Даже беглый взгляд на табл. 3.1 позволяет понять, что для передачи разных видов сигналов требуется различная ширина полосы пропускания системы электросвязи.
Таблица 3.1. Ширина спектров сигналов электросвязи
|
Вид сигнала
|
Ширина спектра, Гц
|
| Телеграфный |
0.
|
| Передачи данных со скоростью 2 400 Бод |
0…2 400
|
| Телефонный |
300…3 400
|
| Звукового вещания |
50…10 000
|
| Факсимильный – при скорости 120 мин-1 – при передаче газет |
0…1 465 0…180 000
|
| Телевизионный |
50…6 000 000
|
 ..100
..100Контрольные вопросы
- На какие простейшие составляющие “раскладывается” периодически повторяющийся прямоугольный импульс?
- Чем отличается спектр периодического сигнала от спектра непериодического сигнала?
- У какого импульса амплитуда спектральных составляющих убывает быстрее: а) более короткого или более длинного? б) с более крутым фронтом или с более пологим? в) повторяющегося чаще или реже?
- Какие частотные диапазоны занимают спектры основных сигналов электросвязи
Список литературы
- Бакалов В. П., Дмитриков В.Ф., Крук Б.И. Основы теории цепей: Учебник для вузов; Под ред. В.П.Бакалова. – М.: Радио и связь, 2000. – 592 с.
- Бакалов В.П., Воробиенко П.П., Крук Б.И. Теория электрических цепей. Учебник для вузов: под ред. В.П. Бакалова – М.: Радио и связь. 1998. – 444 с.
- Бакалов В.П., Журавлева О.Б., Крук Б.И. Анализ линейных электрических цепей: Учебное пособие для дистанционного обучения. – Новосибирск: СибГУТИ. 2001г.
 П., Дмитриков В.Ф., Крук Б.И. Основы теории цепей: Учебник для вузов; Под ред. В.П.Бакалова. – М.: Радио и связь, 2000. – 592 с.
П., Дмитриков В.Ф., Крук Б.И. Основы теории цепей: Учебник для вузов; Под ред. В.П.Бакалова. – М.: Радио и связь, 2000. – 592 с.
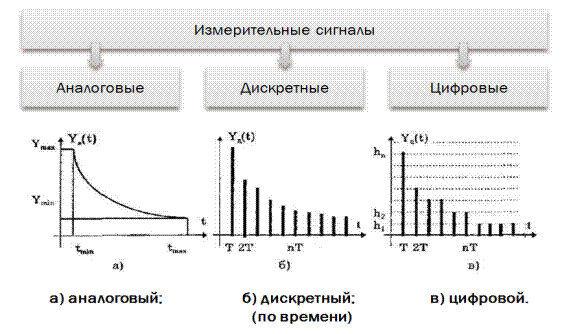
1.3. Основные характеристики Электрических сигналов
Несмотря на большое разнообразие
сигналов, с точки зрения качества
передаваемой информации существует
несколько электрических параметров и
характеристик, которые достаточно
хорошо описывают сигналы. Согласно ИСО
9000 под качеством понимают
«совокупность свойств и характеристик
продукции, способной удовлетворять
установленные и предполагаемые
потребности потребителя продукции».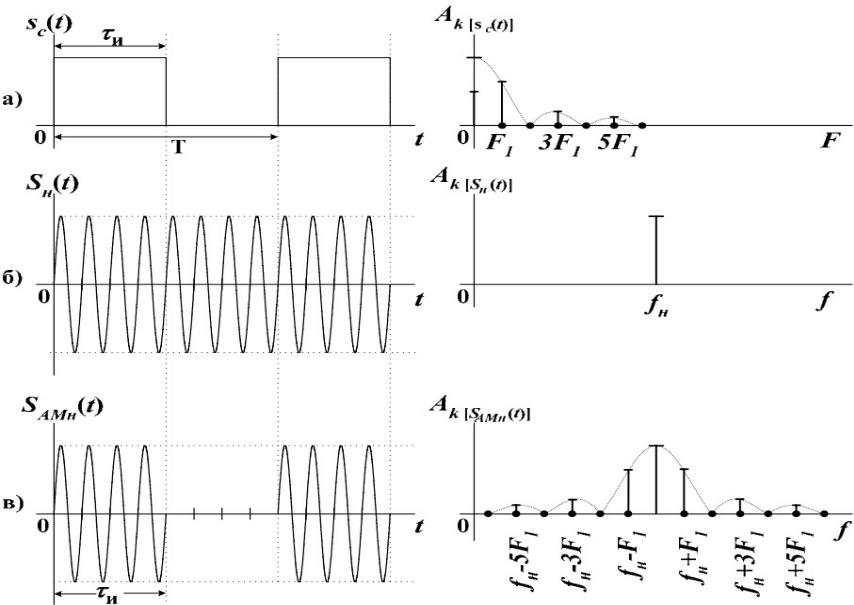 В
В
рассматриваемом случае это — динамический
диапазон, время установления и ширина
спектра сигнала, надёжность получения
её неискажённой.
Динамическим
диапазоном называют отношение
наибольшей мгновенной (так называемой
пиковой) мощности (Pпик)
сигнала к его наименьшей
(часто называемой пороговой)
мощности
(Pпор)
Подчеркнем, что смысл передаваемых
сигналом сообщений ни в коей
мере не зависит от затрачиваемой на их
передачу энергии, которая может изменяться
в широких пределах. Однако помимо
сигналов, несущих нужную нам информацию,
из окружающего нас пространства поступают
посторонние электромагнитные колебания,
так называемые шумы и помехи, которые
маскируют полезный сигнал.
Для удовлетворительного приема сообщений
необходимо иметь
возможность различать полезный сигнал
на фоне помех и шумов, что диктует
ограничения по наименьшей (пороговой)
мощности полезного сигнала. Для краткости
Для краткости
говорят о необходимом отношении
сигнал-помеха, имея в виду отношение
средних мощностей сигнала и помехи и
стараясь повышать его за счет повышения
энергии передаваемого сигнала и снижения
уровня помехи. С другой стороны, чрезмерное
увеличение мощности сигнала может
привести к искажению его формы, а,
следовательно, и содержащейся в нем
информации из-за ограниченной выходной
мощности систем передачи. Появление
таких искажений ставит предел наибольшей
(пиковой) мощности сигнала.
1.3.1. Частотный спектр сигналов.
Оценка по времени установления, т.е.
времени, в течение которого сигнал (с
заданной точностью) достигает
установившегося значения, необходима
для суждения об искажениях, которые
могут возникнуть из-за инерционности
реальных датчиков, устройств передачи
и приема сигналов. Подобные оценки
получают двояким способом: заданием
либо функции времени (временной
характеристики), описывающей реальный
процесс, либо ряда гармонических
колебаний, т. е. спектра, являющегося
е. спектра, являющегося
функцией частоты. При этом оба представления
совершенно равносильны и взаимно
дополняют друг друга, а переход от одного
к другому осуществляется с помощью
прямого и обратного преобразований
Фурье и Лапласа.
В схемотехнике в зависимости от решаемых
задач применяются оба (временное и
частотное) описания. Специалист должен
свободно переходить при исследовании
того или иного явления от временных
представлений к спектральным и обратно.
Следует пояснить, что в принципе можно
было бы обойтись только временными или
только спектральными представлениями.
Но дело в том, что назначение различных
устройств не одинаково. В одних случаях
приходиться пользоваться спектральными
представлениями, в других – временными.
Так, например, назначение всякого
частотного фильтра состоит в том,
чтобы пропустить колебания одних частот
и задержать колебания других частот.
Поэтому работу и свойства фильтра
естественно описывать с помощью
спектральных представлений, а не
временных.
В качестве другого примера возьмем
какую-нибудь цифровую схему преобразования
или передачи импульсов. Если при этом
нас интересует искажение формы импульсов,
то их удобно представить функциями
времени. К этому нужно добавить, что
одна и та же система может использоваться
по-разному и в зависимости от этого
рассматриваться с различных точек
зрения. Примером тому могут служить
простейшие фильтры верхних и нижних
частот, которые во временной области
рассматриваются как укорачивающие и
удлиняющие цепочки.
Таким образом, выбор того или иного
способа описания системы, т.е. формирования
её математической модели, зависит не
столько от её устройства, т.е. структуры
и параметров элементов, сколько от
назначения системы и того на какие
вопросы исследователь хочет получить
ответы. Меняется лишь наша точка зрения
на предмет, но не сам предмет, который
представляет собой некоторую объективную
сущность, не зависящую от способа ее
описания.
Рассмотрим такую характеристику
сигнала, как частотный спектр. Вначале
Вначале
будем предполагать, что сигнал представляет
собой воздействие в виде периодической
функции времени x(t) с периодом
T = 1/f, которую можно представить
рядом Фурье:
x(t) = X0 + X1mcos(w0t — j1) + X2mcos(2w0t — j2) +
+ X3mcos(3w0t — j3) + …,
(1.1)
где w0 = 2pf0;
j1, j2,
j3, … — начальные
фазы отдельных гармоник; X1m,
X2m, X3m,… —
их амплитуды;
;
Здесь
;
.
Смысл формулы (1. 1) состоит в том, что
1) состоит в том, что
периодическая функция x(t) может быть
представлена суммой синусоидальных
колебаний с частотами, кратными основной
частоте, и с надлежащим образом
подобранными амплитудами и начальными
фазами.
Отдельные слагаемые суммы (1.1) называются
гармониками.
Колебания основной частоты w1
называют первой гармоникой, колебание
с частотой w2 –
второй и т. д.
Постоянная составляющая
представляет собой среднее значение
функции x(t). Совокупность величин
Xkm называется спектром
амплитуд; совокупность величин
jл –
спектром фаз. Чаще всего интересуются
только спектром амплитуд и называют
его для краткости просто спектром.
Графически спектр изображают в
координатах Xm, w
(рис. 1.5). Длины вертикальных отрезков
представляют собой амплитуды
соответствующих гармоник; эти отрезки
называют спектральными
линиями, а сам спектр – линейчатым.
В
общем случае сумма (1.1) является бесконечным
рядом. Но в действительности для всех
сигналов число членов ряда, а, следовательно,
и число спектральных линий конечно.
амплитуды
гармоник, начиная с некоторого номера,
становятся настолько малы, что ими можно
пренебречь, не нарушая смысла сообщения.
Таким образом, сигналы в системе
управления и связи практически всегда
представляются функциями с ограниченным
спектром.
Интервал частот, в котором размещается
ограниченный спектр, называется шириной
спектра. Ограничение спектра
производят исходя из допустимого
искажения сигнала, так чтобы не потерять
содержащуюся в нем информацию.
Рассмотрим спектры некоторых типичных
колебаний. Возьмем несущий сигнал
U = U0 cos w0
t и промодулируем его с помощью
сигнала
UАМ(t) = Um
cos W t
В результате получим модулированный
сигнал
UАМ
= (1 + m U0) cos
W
t cos w0
t =
= U0
(sinw0
t + m/2 cos(w0
t + W)
+ m/2 cos(w0
t — W)),
m=Um /U0
Обычно частота W на
один — два порядка превышает высшую
гармонику nw1
сигнала UАМ(t), поэтому
говорят, что амплитуда несущих колебаний
медленно меняется в соответствии с
сигналом, а высокочастотные колебания
являются переносчиком информации.
Например, сигнал медленно меняющегося
постоянного тока U(t) не может пройти
через емкостную цепь, но легко может
быть передан в нагрузку посредством
амплитудной модуляции с последующей
демодуляцией его.
Иначе говоря, АМ-колебание содержит три
составляющие: колебание несущей
частоты w0
и два колебания с частотами w0 ± W
, которые называются боковыми
частотами. Спектр АМ-колебаний состоит
из трех линий. Амплитуда боковых
частот пропорциональна т , т. е.
глубине модуляции: при отсутствии
модуляции боковых частот нет, а при
наиболее глубокой модуляции амплитуды
боковых частот равны половине амплитуды
несущей.
В общем случае при амплитудной модуляции
не гармоническим сигналом (рис.1.5,а)
ширина спектра АМ-колебаний равна
удвоенной ширине спектра модулирующей
функции (точнее, удвоенной высшей частоте
этого спектра (рис.1.5, б) ).
при многоканальной
связи каждый из каналов должен иметь
рассмотренную ширину полосы пропускания,
а также некоторый запас для возможных
отклонений от номинального значения
несущей частоты и частот сигнала. На
На
низкой частоте несущей число каналов
получается относительно немного. Поэтому
в настоящее время стремятся перейти на
передачу сигналов с более высокочастотной
несущей частотой – это диапазон сотен
мегагерц – единиц гигагерц.
Достоинство амплитудной модуляции
состоит в том, что её реализация
схемотехнически довольно проста. Кроме
того она позволяет без проводов передавать
сигналы на расстояния, огибающие земной
шар. Как следствие, она в 20 веке широко
применялась особенно в радиовещательной
технике. Однако, такие каналы связи
чрезвычайно плохо защищены от атмосферных
и технических помех.
В связи с развитием спутниковой без
проводной связи, а также использование
оптоволоконных кабелей, привело к тому,
что стали применяться более широко
системы передачи информации даже на
небольшие расстояния, например в одном
здании, с использованием других типов
модуляции, лишённые указанных недостатков.
При частотной модуляции амплитуда
несущих колебаний постоянна и не зависит
от сигнала, поэтому вышеперечисленные
помехи при этом типе модуляции не
искажают передаваемую информацию.
Приращение пропорциональное сигналу
x(t), получает частота несущих
колебаний
или фаза колебаний при фазовой модуляции
Здесь Dw и Dj
¾ частотное и
фазовое отклонения от номинального
значения, которые определяют глубину
модуляции и выбираются, исходя из задачи,
решаемой проектировщиком.
Рассмотрим передачу информации
импульсными сигналами. Для импульсов
прямоугольной формы постоянная
составляющая
а амплитуда n-й гармоники
выражается формулой
Отдельные составляющие спектра отстоят
друг от друга на частоту импульсов, т.е.
в спектре содержатся только частоты
f0, 2f0,
3f0, …
и т.д. Амплитуды гармоник пропорциональны
амплитуде импульсов (рис.1.6), но на
частотах, где аргумент синуса равен kp,
на частотах
они обращаются в нуль.
Суммарная энергия всех колебаний,
составляющих спектр импульса, равна
энергии сосредоточенной в импульсе.
Расчёты показывают, что в полосе частот
0 < f < 2tu
заключено примерно 95% всей энергии
импульса. Наибольшей сосредоточенностью
энергии в спектре обладают импульсы
треугольной и колокольной формы. Следует
также учесть, что с уменьшением частоты
следования импульсов (f
= 1/T) из-за
увеличения скважности импульсов T/
tu
спектр расширяется. В пределе для
одиночного импульса нижняя граничная
частота простирается до нуля, а верхняя
частота ¾
примерно до частоты f
= 2/tu
. С уменьшением скважности, спектр
импульса сужается.
Рис.1.6
Таким
образом, при построении каналов передачи
информации и проектировании электронных
устройств необходимо учитывать не
только верхнюю и нижнюю частоту следования
сигналов, но и длительность импульсов.
Расширение спектра при увеличении
скважности можно использовать для
быстрого экспериментального нахождения
амплитудно-частотной характеристики
устройств (см. раздел 4).
С этой целью длительность импульса
делают намного меньше периода колебаний
(в пределе желательно подавать импульсы,
приближающиеся по спектру к так
называемому d импульсу,
у которого произведение амплитуды,
стремящейся к бесконечности, на
длительность, стремящейся к нулю, даёт
единичную площадь), тем самым достигают
почти равномерного спектра.
Прошедший через исследуемое устройство
или канал связи δ импульс, создаст на
их выходе спектр, повторяющий частотную
характеристику компонента или канала.
3. Первичные электрические сигналы и их характеристики
•Выдачу сертификатов на оборудование систем связи
спроверкой на соответствие международным (международного союза электросвязи МСЭ ) и Российским стандартам.
В) Государственная комиссия по радио частотам при МС обеспечивает:
•Выделение диапазонов частот.
Г) ФАПСИ при Президенте РФ обеспечивает:
• Лицензирование и сертификацию в области защиты информации с целью обеспечения информационной безопасности РФ.
Всоответствии с определением, принятым в теории информации, сигнал есть материальный носитель информации. В системах связи сигнал представляет собой процесс изменения во времени и в пространстве некоторой физической величины, характеризующей передаваемое сообщение. Так, например, при разговоре (акустическая связь) переносчиком информации служит звуковое поле; изменение звукового давления, характеризующее передаваемое сообщение, является в этом случае сигналом. В системах электрической связи сигналы представляют собой электромагнитные процессы, параметры которых изменяются
всоответствии с передаваемым сообщением. Например, мгновенное значение тока
вцепи микрофона пропорционально звуковому давлению; микрофон преобразует акустические сигналы в электрические и, таким образом, дает возможность передавать звуковые сообщения по кабелям электрической связи.
Впроцессе передачи сигнал подвергается ряду преобразований, важнейшими из которых являются модуляция и кодирование и обратно – демодуляция и декодирование. В импульсных и цифровых системах связи сигналы подвергаются дискретизации. При передаче по каналу связи сигналы претерпевают искажения, связанные с несовершенством устройств, входящих в канал, и маскируются помехами. Система связи должна быть построена так, чтобы, несмотря на искажения и помехи, возникающие при передаче сигналов, было бы возможно восстановление исходного сообщения с необходимой степенью точности. Чтобы выполнить это условие, при построении систем связи необходимо знать основные характеристики сигналов, соответствующие различным сообщениям.
9
В системе электрической связи важную роль играют первичные преобразователи, превращающие сообщения источника в электрические сигналы. При передаче телефонных сообщений и звуковом вещании первичными преобразователями служат микрофоны, в телеграфии – телеграфные аппараты, при передаче изображений в факсимильной связи и телевидении – устройства электрооптического анализа. Сигналы, формируемые первичными преобразователями, называются первичными сигналами.
Ясно, что свойства первичных сигналов в значительной мере определяют требования к системам связи. Подходящей математической моделью для описания сигналов связи являются случайные процессы; свойства случайных процессов характеризуются n-мерной функцией распределения и тем точнее, чем больше n. Однако, практическое определение многомерных функций распределения связано с чрезвычайными трудностями, поэтому в большинстве случаев для описания сигналов пользуются понятием энергетического спектра и некоторыми числовыми характеристиками. К ним относятся:
Полоса частот, необходимая для передачи сигнала с допустимыми искажениями, f (часто ее называют эффективной полосой)
Мощности сигнала – пиковая или максимальная Pмакс,, средняя
Pср и минимальная Pмин.
В качестве энергетических характеристик сигналов часто используют понятия динамического диапазона
D =10lg | Pмакс |
| , дБ | (3.1) | |||
P |
|
| |||||
|
|
|
|
|
| ||
и пикфактора | мин |
|
|
|
| ||
|
|
|
|
|
| ||
ν =10lg | Pмакс |
| , | дБ. | (3.2) | ||
| |||||||
|
| P |
|
|
|
| |
|
| ср |
|
|
|
| |
Рассмотрим характеристики первичных сигналов.
Телефонные (речевые) сигналы
Звуки речи представляют собой сложные звуковые колебания, следовательно, первичные сигналы, формируемые микрофоном, также очень сложны.
Спектр звуков речи содержит колебания с частотами от 70-80 до 8000-20000 Гц. Однако основная мощность речевых сигналов сосредоточена в полосе частот от 150 до 1500-2000 Гц. В результате действия резонирующих полостей речеобразующего тракта человека (полости рта и носа) в спектре звуков создаются области повышенной интенсивности, называемые формантами. Большая часть формантных областей расположена в диапазоне частот от 300 до 3000 Гц.
Ограничение спектра речевых сигналов приводит к ухудшению качества принимаемой речи – искажается тембр, теряется естественность и узнаваемость, снижается разборчивость. Экспериментальные исследования показали, что вполне удовлетворительное качество речи получается при передаче в полосе частот от 250-300 до 3300-3500 Гц. Поэтому полоса эффективно передаваемых частот стандартного канала тональной частоты находится в пределах от 300 до 3400 Гц.
10
Мощность разговорных звуков изменяется в очень широких пределах от 0,01мкВт при шепоте до 10000мкВт при крике. Средняя мощность речи при нормальном разговоре с учетом пауз между словами и фразами равна приблизительно 10мкВт. Таким образом, при учете всех возможностей речи динамический диапазон речевых сигналов оказывается чрезвычайно большим D = 55 дБ, ν = 27,5 дБ.
Однако при расчете каналов телефонной связи не следует ориентироваться на необходимость передачи сигналов с таким большим динамическим диапазоном, т.к. это связано с серьезными техническими трудностями. Кроме того, исследования показывают, что в большинстве случаев динамический диапазон речи значительно меньше. Так, например, при плавном разговоре со средним уровнем громкости пикфактор не превосходит величины ν =14 −18 дБ. Поэтому, при передаче речевых сообщений принимают динамический диапазон равным 23-27 дБ.
Кроме речевого сигнала телефонная система передачи должна передавать специальные сигналы управления и взаимодействия СУВ на другой конец связи. По цепи между телефонной станцией и абонентом передаются следующие сигналы: постоянным током для дистанционного управления, образующиеся снятой и лежащей на месте телефонной трубкой, импульсы или частоты набора номера, а также сигнала вызова переменным током.
Эту информацию на центральной станции преобразуют для передачи по специальным сигнальным проводам, называемым проводами E и M (E &M сигнализация). Сигналы, передаваемые постоянным током по проводу M с вызывающей станции, поступают в провод E на вызываемой станции и наоборот.
Адресация или указание номера вызываемого абонента производится вызывающим абонентом. Это делается, в основном, двумя способами – набирая номер диском телефонного аппарата или расположенном на нем кнопками. При наборе номера диском телефонного аппарата обрывается токовая цепь между абонентом и телефонной станцией. При наборе номера кнопками передаются либо импульсы определенной частоты, либо тональные частоты.
Сигналы акустического вещания
Спектр сигналов возникающих при исполнении вокальных и инструментальных музыкальных произведений, расположен в полосе частот от 16 до 15000-20000 Гц. Однако, в зависимости от требования к качеству воспроизведения полоса частот, необходимая для передачи сигналов акустического вещания, может быть ограничена. Так, для каналов первого класса — от 50 до 10000 Гц и для каналов высшего класса от 30 до 15000 Гц.
Динамический диапазон сигналов акустического вешания чрезвычайно велик. Так, динамический диапазон симфонического оркестра достигает 100 дБ, т.е. максимальная мощность сигнала больше минимальной в 1010 раз. Передать по линии сигнал с таким динамическим диапазоном чрезвычайно трудно – потребуется очень мощная дорогостоящая аппаратура. Поэтому, не снижая заметно качества передачи, ограничивают динамический диапазон сигналов
11
вещания величиной 40 дБ для каналов первого класса и 50 дБ для каналов высшего класса.
Сигналы телевизионного вещания
Полный телевизионный сигнал черно-белого телевидения состоит из сигналов изображения, синхронизации и гашения обратного хода луча приемной трубки по строкам и кадрам. Сигналы синхронизации содержат синхронизирующие импульсы строчной и полукадровой частоты и уравнивающие импульсы.
Рис.3.1. Стандартные телевизионные видеосигналы: а) В начале каждого первого поля, б) В начале каждого второго поля.
Отечественный стандарт на черно-белое телевидение предусматривает передачу 25 кадров в секунду. Каждый кадр составляется из двух полей (полукадров). Число строк в кадре – 625. Развертка изображения чересстрочная, т.е. поля с нечетными строками и поля с четными строками передаются поочередно. Это позволяет ослабить визуальный эффект мерцания воспроизводимого изображения без увеличения числа элементов изображения, передаваемых за одну секунду. Отношение ширины кадра к его высоте (формат кадра) равна 4:3.Развертка изображения осуществляется по строкам слева на право и по полукадрам сверху вниз. Номинальное значение частоты строчной развертки fc =15625 Гц, а частоты полей 50 Гц. Во всех точках соединения по видеочастоте в трактах передачи телевидения номинальный размах полного видеосигнала должен
12
составлять 1 В. Время передачи одной строки Tc=64 мкс. Длительность строчных импульсов гашения 12 мкс, а синхронизирующих 4,7 мкс. Импульсы синхронизации передаются ниже уровни черного. Введение уравнивающих импульсов (частота следования 2fc, а длительность 2,35 мкс) и нескольких кадровых синхронизирующих импульсов (один широкий, разделенный врезками на пять) обеспечивает идентичные условия работы устройств синхронизации полукадров при передаче четных и нечетных строк и непрерывность работы строчной синхронизации во время полукадровых импульсов.
В современных системах цветного телевидения многоцветное изображение разделяется на три одноцветных, называемых цветоделенными. Они соответствуют основным цветам: красному R, зеленому G и синему B. Однако, передаются не сигналы, соответствующие этим трем цветам, а сигналы яркости и цветности изображения, образуемые из этих трех сигналов. Сигнал яркости создает нормальное черно-белое изображение и обеспечивает передачу необходимого числа градаций яркости передаваемого изображения. Он получается путем сложения в определенном соотношении сигналов основных цветов. Цветность изображения передается с помощью двух цветоразностных сигналов. Таким образом, обеспечивается совместимость черно-белого и цветного телевидения.
Системы ЦТВ – NTSC, PAL и SEKAM отличаются главным, образом, способом модуляции цветовой поднесущей частоты цветоразностными сигналами. Ширина видеоканала 6 МГц, расстояние между несущими звука и изображения 6,5 МГц, ширина радио канала телевизионного вещания 8 МГц.
Сигналы передачи данных |
|
|
|
|
|
|
|
| |
Сигналы передачи данных обычно имеют вид последовательностей | |||||||||
|
| двуполярных | или | однополярных | |||||
c(t) |
| прямоугольных импульсов. | |||||||
|
|
| Сигналы |
| такой |
| формы | ||
с |
| называют |
|
|
| двоичными. | |||
τи | τи | Длительность |
|
| импульсовτи | ||||
|
| определяется | скоростью | передачи | |||||
|
| B, | измеряемой | в | бит/с | (число | |||
| t | символов | в | секунду). | Вводится | ||||
-c |
| понятие тактовой частоты |
| Fт = 1 , | |||||
|
|
|
|
|
|
|
|
| τ |
|
|
|
|
|
|
|
|
| и |
c(t) |
| которая численно равна | скорости | ||||||
| передачи. |
|
|
|
|
|
| ||
|
|
|
|
|
|
|
| ||
c | τи |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
| t |
|
|
|
|
|
|
|
|
Рис.3.2. Двоичные импульсы передачи данных |
|
|
|
|
|
|
|
| |
13
Вероятность появления положительных p(+C) и отрицательных p(-C) импульсов, а также статистические связи между импульсами определяются свойствами источника информации. Часто p(+C)=p(-C)=0,5 и импульсы последовательности статистически независимы.
Энергетический спектр такого сигнала
| τ и |
| sin | ωτ | и |
| |
Ф (ω ) = с 2 |
| 2 |
| . | (3.3) | ||
|
| ||||||
|
|
|
| ||||
| π | ωτ и |
|
|
| ||
|
|
|
| 2 |
|
|
|
Подставляя | ω = 2π f | и τи | = | 1 | ||||||||||
| Fт | |||||||||||||
|
|
|
|
|
|
|
|
|
|
|
| |||
↑Ф | ( f | / F |
| ) = Ф( f / F ) |
| π 1 | ||||||||
|
|
|
|
|
|
|
| |||||||
|
| с2 τи | ||||||||||||
н |
|
| т |
|
| т |
| |||||||
получим |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| f |
|
|
|
| sinπ f Fт |
|
|
|
| ||||
|
|
|
|
|
|
|
|
| ||||||
Фн |
|
|
| = |
|
|
| . |
|
| ||||
F |
| π f | F |
|
|
| ||||||||
| т |
|
|
| т |
|
|
|
|
|
|
| ||
и обозначая нормированный спектр
,
(3.4)
Определим минимальную полосу частот, F , необходимую для передачи сигнала данных.
Выражение для Ф(ω) показывает, что для неискаженной передачи последовательности прямоугольных импульсов необходима бесконечно широкая полоса частот, т.к. спектральная плотность сигнала не равна нулю на всей оси частот.
Фн |
|
|
1,0 |
|
|
0,5 |
|
|
|
| fв |
0 | 0,5 | 1,0 |
Следует, однако, иметь ввиду, что при передаче двоичных сигналов в приемнике нет необходимости восстанавливать импульсы без искажений, т.е. сохранять их прямоугольную форму.
Рис.3.3. Энергетический спектр сигнала передачи данных
Для восстановления информации достаточно зафиксировать только знак импульса при двухполярном сигнале, либо наличие или отсутствие импульсов при однополярном.
Исходя из этого, будем решать задачу определения величины F .
Пусть на вход идеального фильтра низких частот ФНЧ с частотной характеристикой
14
| 1 при ω | ≤ω | с |
|
|
|
|
|
|
| |||
|
|
|
|
|
|
|
|
| (3.5) |
|
| ||
K (ω) = |
|
| >ω |
|
|
|
|
|
|
| |||
| 0 при ω | с |
|
|
|
|
|
|
| ||||
|
|
|
|
|
|
|
|
|
|
|
|
| |
подается | последовательность из двух прямоугольных | импульсов | f1 (t) | ||||||||||
(Рис.3.4). |
|
|
|
|
|
| Найдем | сигнал на | выходе фильтра | ||||
f1(t) |
|
|
|
|
|
| |||||||
|
|
|
|
| F2 (t) при различных значениях частоты | ||||||||
1 |
|
|
|
|
| среза ωc = 2π fс . |
|
|
|
| |||
|
|
|
|
|
| Сигнал | f1 (t) можно | представить | |||||
|
|
|
|
|
| аналитически |
|
| в |
| виде | ||
|
|
|
|
|
| f1 (t) =σ(t) −σ(t −τи ) +σ(t − 2τи ) −σ(t −3τи) , | где | ||||||
|
|
|
|
|
| σ(t) — единичный скачок. |
|
|
| ||||
τи | τи | τи |
|
| t |
| 0 | при t < 0 .(3.6) |
|
| |||
|
|
| σ(t) = |
|
| ||||||||
|
|
|
|
|
|
|
| 1 при t | > 0 |
|
|
| |
| Рис.3.4. |
|
|
|
|
|
|
|
|
|
| ||
Известно, что скачок на выходе идеального фильтра при подаче на его вход | |||||||||||||
единичного скачка определяется выражением |
|
|
|
|
|
| |||||||
h(t) = | 1 + | 1 | Siωc t , |
|
|
|
|
|
| (3.7) |
|
| |
| 2 | π |
|
|
|
|
|
|
|
|
|
|
|
где Si z = ∫z sin xdx — интегральный синус. |
|
|
|
|
|
| |||||||
0 | x |
|
|
|
|
|
|
|
| на выходе | идеального | ||
Используя | принцип | суперпозиции, | найдем | f2 (t) | |||||||||
фильтра |
|
|
|
|
|
|
|
|
|
|
|
|
|
f2 (t) = 1 [Siωct − Siωc (t −τи ) + Siωc (t − 2τи ) − Siωc (t −3τc )]. | (3.8) |
|
| ||||||||||
| τ |
|
|
|
|
|
| f2 (t) при различных значениях | |||||
На рис.3.9 представлены графики функции | |||||||||||||
частоты среза ωс .Как видно наличие или отсутствие импульсов можно уверенно | |||||||||||||
зафиксировать при | fс | = 1 | = 0,5FT = 0,5B . |
|
|
|
|
|
|
| |||
|
|
| 2τи |
|
|
|
|
|
|
|
|
|
|
Таким образом, если частотные характеристики канала связи приближаются | |||||||||||||
к характеристикам идеального ФНЧ, то эффективная полоса частот двоичного | |||||||||||||
сигнала составляет |
| F = 0,5FT . При наличии частотных искажений в реальных | |||||||||||
каналах приходится несколько расширять полосу частот, отводимую для передачи | |||||||||||||
импульсных сигналов. Часто принимают | F = FT − B . |
|
|
|
|
| |||||||
|
|
|
|
|
| 15 |
|
|
|
|
|
|
|
f 2 |
|
| τи |
| t |
fc | = 2FT |
f 2 |
|
| τи |
| t |
fc | = 0,5FT |
f 2 |
|
| τи |
| t |
fc | = FT |
Основные характеристики переменных электрических сигналов
Переменный электрический сигнал в большинстве случаев представляет собой функцию времени u(t) с периодом Т = 1/f, которую можно записать в виде ряда Фурье: u(t) = U0 + U1maxcos(ω0t–φ1) + U2maxcos(2ω0t–φ2) +U3maxcos(3ω0t–φ3) +…, где ω0 = 2πƒ0; φ1, φ2, φ3,.– начальные фазы отдельных гармоник; U1max, U2max, U3max,… – их амплитуды.
Отдельные слагаемые суммы называют гармониками, причем колебания основной частоты называют первой гармоникой и т.д.
Совокупность величин UКmax называют спектром амплитуд, совокупность величин φК – спектром фаз. Чаще всего интересуются только спектром амплитуд и называют его для кратности просто спектром. Графические изображения спектров даны на рис
Длины вертикальных отрезков представляют собой амплитуды соответствующих гармоник. Эти отрезки называют спектральными линиями, а сам спектр – линейчатым.
В общем случае сумма является бесконечным рядом, т. е. спектр сигнала бесконечен. Так как амплитуды гармоник по мере увеличения их номера (угловой частоты ω) убывают, начиная с некоторой гармоники, высокочастотными колебаниями пренебрегают, тем самым, ограничивая спектр сигнала.
Интервал частот, в котором размещается ограниченный спектр, называется шириной спектра. Ограничение спектра производят исходя из допустимого искажения сигнала так, чтобы не потерять содержащуюся в нем информацию.
Рис.1.9. Форма сигналов и графическое изображение спектра: а) прямоугольных импульсов, б) пилообразных импульсов
На рис.1.10 показано, как формируются прямоугольные импульсы из гармоник.
Графики 1-й (n=1) и 3-й (n=3) гармоник и их суммы изображены на рис.1.10, а. На рис.1.10, б эта сумма дополнена 5-й гармоникой, а на рис.1.10, в – 7-й. Характерно, что
прямоугольные импульсы, также как и треугольные, являются четными относительно t и содержат только нечетные гармоники.
Пилообразное колебание является нечетным относительно t и содержит в своем составе как четные, так и нечетные гармоники. Форма и спектр пилообразного сигнала изображены на рис.
Какими бывают слуховые аппараты
Слуховой аппарат — электроакустический звукоусиливающий прибор индивидуального пользования. Его основное назначение — преобразование сигнала, создаваемого источником звуковой информации, таким образом, чтобы этот сигнал смог быть воспринят слабослышащим человеком с достаточно высокой степенью слухового ощущения. Для этого слуховой аппарат усиливает звуковые сигналы, а также изменяет их динамические и частотные характеристики в соответствии со степенью и характером нарушения слуха.
В связи с многообразием функций слуховых аппаратов существует несколько вариантов их классификации. Любой слуховой аппарат рассматривается с разных точек зрения и находит свое место в каждой из классификаций.
По способу обработки сигнала слуховые аппараты делятся на два типа: аналоговые и цифровые.
Аналоговый слуховой аппарат состоит из трех основных частей: микрофона, электронного усилителя и телефона. Микрофон воспринимает механические звуковые колебания и преобразует их в аналоговые электрические сигналы, которые подает в усилитель. Там они усиливаются и передаются на телефон, который превращает усиленные электрические сигналы вновь в звуковые колебания и подает их в ухо.
Цифровой слуховой аппарат дополнительно преобразует аналоговые сигналы в цифровые, после чего обрабатывает их с помощью компьютерной технологии. Вместо усилителя он имеет интегральную электронную схему, состоящую из трех элементов.
Аналоговый сигнал из микрофона поступает в аналого-цифровой преобразователь, который преобразует электрические сигналы в цифровой вид — двоичный код, как это происходит при записи на компакт-диск. (В новейших моделях слуховых аппаратов уже появились цифровые микрофоны, исключающие эту операцию). Далее сигнал поступает на цифровой сигнальный процессор, крохотную компьютерную микросхему. Она обрабатывает цифровые сигналы, то есть усиливает их и изменяет их характеристики в зависимости от индивидуальной потери слуха. После этого цифро-аналоговый преобразователь вновь превращает цифровой сигнал в аналоговый и посылает его на телефон.
Цифровые технологии, бурно развивающиеся в последнее время, позволили достигнуть невиданных ранее возможностей электроакустической коррекции слуха. Крошечный микрочип обладает быстродействием самых современных компьютерных процессоров, что позволяет реализовать очень сложные и высокоэффективные алгоритмы обработки звука. Фактически цифровой СА можно назвать «разумной слуховой системой» и даже «слуховым компьютером». Он «умеет» отличать речь от шума, выделяя речевой сигнал и усиливая его при одновременном подавлении шумового сигнала, что значительно облегчает понимание речи в шумной обстановке. Он может быть очень точно настроен в соответствии с индивидуальной потерей слуха, поскольку его частотный диапазон разделен на несколько частотных каналов, в каждом из которых проводится независимая настройка параметров. Цифровой аппарат имеет комфортное звучание, приближенное к естественному, благодаря практически полному отсутствию искажений и собственных шумов. Наконец, он устойчив к воздействию электромагнитных полей, что позволяет в условиях активной современной жизни без помех пользоваться мобильным телефоном и компьютером.
По способу настройки слуховые аппараты также делятся на два типа.
Непрограммируемый слуховой аппарат настраивается вручную, при помощи отвертки и регуляторов (триммеров), а громкость звучания по мере необходимости регулирует сам владелец посредством регулятора громкости.
Программируемый слуховой аппарат подключается через кабель к компьютеру и настройка осуществляется в цифровом виде, что обеспечивает более точное ее соответствие индивидуальным особенностям слуха. Аппарат может сохранять и изменять запрограммированную настройку. Большинство программируемых слуховых аппаратов могут иметь две и более слуховые программы с разными настройками: для прослушивания речи в шумной обстановке, для прослушивания музыки, программу комфортного слуха и пр.
Существует еще одна вспомогательная классификация слуховых аппаратов: по способу усиления они делятся на линейные и нелинейные.
Линейный слуховой аппарат — аппараты этого типа усиливают входные сигналы независимо от их уровня (громкости) на одну и ту же величину, зафиксированную при помощи регулятора усиления. В линейных аппаратах с выходным уровнем звукового давления, превышающим 130 дБ, предусматривается регулятор ограничения выходного уровня (пик-клиппирование), который вводится в действие при восприятии пользователем дискомфортной громкости звуков.
Нелинейный слуховой аппарат — коэффициент усиления этих аппаратов, имеющих функцию автоматической регулировки усиления (АРУ или компрессия) зависит от уровня входного сигнала. До тех пор, пока уровень входного сигнала не достигнет определенной величины, называемой порогом срабатывания АРУ, коэффициент усиления остается постоянным, как у линейного аппарата. При превышении входным сигналом порога срабатывания АРУ, который устанавливается слухопротезистом в соответствии с индивидуальной потерей слуха, коэффициент усиления аппарата снижается. При этом происходит сжатие (компрессия) динамического диапазона выходных сигналов (для протезирования сенсоневральной тугоухости с ФУНГом).
По способу звукопроведения слуховые аппараты разделяются на два вида: костной и воздушной проводимости.
Слуховой аппарат костной проводимости — применяется для протезирования только кондуктивных потерь слуха. Его телефон выполнен в виде костного вибратора, который помещается за ухом и плотно прилегает к сосцевидному отростку. На выходе усиленный сигнал преобразуется не в звуковой, а в вибрационный.
Слуховой аппарат воздушной проводимости — используется для протезирования всех видов потерь слуха. Звук с телефона передается через ушной вкладыш, который помещается в слуховой проход (см. выше).
Конструктивно (по месту ношения) слуховые аппараты разделяются на четыре вида: заушные, внутриушные, карманные и очковые.
Внутриушной слуховой аппарат полностью размещается в слуховом проходе. Все электронные компоненты находятся в корпусе аппарата, который изготавливается индивидуально, в соответствии с анатомическим строением уха владельца. Основное достоинство аппарата заключается в его малозаметности и в том, что отверстие приема звука располагается внутри ушной раковины, то есть там, где это предусмотрено природой.
В классификации внутриушных аппаратов выделяют аппараты внутриканального типа, которые располагаются глубоко в слуховом проходе, не закрывая полость ушной раковины. Самый маленький аппарат CIC (с английского -«полностью внутри канала»), размещается у барабанной перепонки и снаружи практически не виден.
Карманный слуховой аппарат состоит из прямоугольного корпуса, в котором расположены микрофон, усилитель и источник питания. Телефон карманного аппарата при помощи шнура соединяется с корпусом и помещается в ухо вместе с вкладышем. Карманный слуховой аппарат, в отличие от других конструкций, может иметь максимальную мощность, так как микрофон и телефон и телефон находятся на значительном расстоянии, что предотвращает возникновение акустической обратной связи.
Очковый слуховой аппарат — аппарат, компоненты которого вмонтированы в дужку очков. В очковом слуховом аппарате костной проводимости телефон (вибратор) расположен на внутренней стороне дужки так, чтобы при одевании очков обеспечивалось его надежное прилегание к сосцевидному отростку (мастоиду).
Заушный слуховой аппарат помещается за ушной раковиной. К нему с помощью звукопроводящей трубочки присоединен ушной вкладыш, который вставляется в слуховой проход. Он проводит звук в ухо и обеспечивает фиксацию аппарата. Заушный слуховой аппарат обеспечивает большее усиление и предоставляет дополнительные технические возможности по сравнению с внутриушным слуховым аппаратом.
На рисунке 1 представлен внешний вид заушного слухового аппарата с наиболее распространенными названиями его составных частей:
Рис. 1. Внешний вид слухового аппарата
1) Корпус.
2) Крышка.
3) Держатель элемента питания (обойма, батарейный отсек).
4) Переход (рожок, звуковой крючок).
5) Регулятор громкости (регулятор усиления, оперативный регулятор).
6) Регуляторы настройки (триммеры, неоперативные регуляторы) — могут находиться с внутренней стороны слуховые аппараты.
7) Переключатель режимов (включение-выключение).
8) Звукопроводящая трубочка.
9) Стандартный ушной вкладыш, он же звуковод.
Ушной вкладыш является неотъемлемой частью заушного слухового аппарата. От него во многом зависит успех слухопротезирования. Ушные вкладыши бывают стандартные и индивидуальные, изготавливаемые по форме уха пациента. Индивидуальный вкладыш имеет ряд преимуществ перед стандартным: его форма и размеры точно соответствуют особенностям анатомии слухового прохода, что обеспечивает герметичность и надежную фиксацию в ухе. Изготовление вкладыша любой формы, различных диаметров звукопроводящего и вентиляционного отверстий существенно влияет на частотную характеристику слухового аппарата. Наличие во вкладыше вентиляционного отверстия (при потере слуха не более 80 дБ) способствует также «проветриванию» уха и предотвращает скапливание конденсата.
Без использования индивидуального вкладыша невозможно достичь успешного слухопротезировния.
Электрический сигнал — обзор
Видеосигнал и его спектр
В этом заголовке обсуждаются теоретические основы ограничений, полосы пропускания и разрешения видеосигнала. Это сложный предмет, основы которого включают высшую математику и электронику, но я постараюсь объяснить его простым и понятным языком.
Большинство искусственных электрических сигналов можно описать математическим выражением. Математическое описание очень просто для периодических сигналов, таких как, например, основное питание.Периодическая функция всегда может быть представлена суммой синусоид, каждая из которых может иметь разную амплитуду и фазу. Подобно спектру белого света, это называется спектром электрического сигнала . Чем более периодичен электрический сигнал, тем легче он может быть представлен и с меньшим количеством синусоидальных составляющих. Каждая составляющая синусоидальной волны может быть представлена дискретным значением в частотном спектре сигнала. Чем менее периодична функция, тем больше компонентов потребуется для воспроизведения сигнала.Теоретически даже непериодическая функция может быть представлена суммой различных синусоид, только в таком случае будет намного больше синусоид (с разными частотами), которые нужно суммировать, чтобы получить непериодические периодический результат. Другими словами, спектральное изображение непериодического сигнала будет иметь полосу пропускания, более плотно заполненную различными компонентами. Чем мельче детализация сигнала, тем выше будут частоты в спектре сигнала. Очень мелкие детали видеосигнала будут представлены высокочастотными синусоидальными волнами.Это эквивалентно более высокой информации о решении. Сигнал, богатый высокими частотами, будет иметь более широкую полосу пропускания. Даже одиночный, но очень резкий импульс будет иметь очень широкую полосу пропускания.
Типичный анализатор спектра
Вышеупомянутое очень упрощенно описывает очень важную спектральную теорию Фурье , , которая утверждает, что каждый сигнал во временной области имеет свое изображение в частотной области. Спектральную теорию Фурье можно использовать на практике — широкополосных периодических электрических сигналов можно более эффективно исследовать, анализируя их частотный спектр. Не углубляясь в саму теорию, пользователи CCTV должны принять концепцию спектрального анализа как очень важную для изучения сложных сигналов, таких как само видео. Видеосигнал, возможно, является одним из самых сложных электрических сигналов, когда-либо созданных, и его точное математическое описание практически невозможно из-за постоянного изменения сигнала во временной области. Видеоинформация (то есть компоненты яркости и цветности) постоянно меняется.Однако, поскольку мы составляем видеоизображения с помощью периодического сканирования луча, мы можем приблизить видеосигнала с некоторой формой периодического сигнала. Одним из основных компонентов этой периодичности будет частота сети — для PAL 25 × 625 = 15,625 Гц; для NTSC 30 × 525 = 15 750 Гц.
Можно показать, что спектр упрощенного видеосигнала состоит из гармоник (кратных) линейной частоты, вокруг которой присутствуют сопутствующие компоненты, как с левой, так и с правой стороны (боковые полосы ). Межкомпонентные расстояния зависят от содержимого видеоизображения и динамики движения. Кроме того, очень важно отметить, что такой спектр, состоящий из гармоник и их компонентов, сходится на , что означает, что гармоники становятся меньше по амплитуде с увеличением частоты. Еще один важный вывод из спектральной теории Фурье состоит в том, что позиций гармоник и составляющих в спектре видеосигнала зависят только от анализа изображения (соотношение 4: 3, 625 чересстрочная развертка).Распределение энергии видеосигнала вокруг гармоник зависит от содержания изображения. Однако гармоники — это в точных положениях, потому что они зависят только от частоты сети. Другими словами, динамика видеосигнала и амплитуда определенных компонентов в боковых полосах будет изменяться, но местоположения гармоник (как частоты поднесущих) останутся постоянными.
Это очень важный вывод. Это помогло найти способ в широковещательном телевидении сократить спектр видеосигнала до минимально необходимой полосы пропускания без потери слишком многих деталей.Конечно, всегда есть компромисс, но, поскольку , большая часть энергии видеосигнала находится около нулевой частоты и первых нескольких гармоник, нет необходимости и нет способа передавать весь видеоспектр. Ученые и инженеры использовали все эти факты, чтобы найти компромисс, чтобы определить, насколько небольшая часть полосы пропускания видеосигнала должна использоваться при передаче, не теряя слишком много деталей. Как мы уже упоминали при обсуждении различных телевизионных стандартов, чем больше линий развертки используется в системе, тем шире будет полоса пропускания, и чем выше разрешение сигнала, тем шире будет полоса пропускания.
Принимая во внимание ограниченный размер электронного луча (который также определяет наименьшие воспроизводимые элементы изображения), физический размер экранов аналогового телевизора, расстояние просмотра, а также сложность и стоимость производства домашних телевизоров, был сделан вывод, что для хорошее воспроизведение вещательного сигнала, достаточно полосы видеосигнала 5 МГц. Использование более широкой полосы пропускания возможно, но коэффициент прироста качества по сравнению с затратами очень низок. На самом деле, в студиях вещания камеры, записывающее и мониторинговое оборудование имеют гораздо более высокие стандарты, со спектрами до 10 МГц.
Это только для внутреннего использования, однако для качественной записи и перезаписи. Прежде чем такой сигнал будет модулирован РЧ и отправлен на этап передачи, он обрезается до видео 5 МГц, к которому добавляется около 0,5 МГц для левого и правого аудиоканалов. Когда такой сигнал поступает на каскад телевизионного передатчика, он модулируется таким образом, чтобы передавалась только его рудиментарная боковая полоса с общей полосой пропускания, включая буферную зону разделения, 7 МГц (для PAL). Но обратите внимание, что фактическая полезная полоса пропускания при широковещательном приеме составляет всего 5 МГц.Для более любопытных читателей мы должны упомянуть, что в большинстве стран PAL видеосигнал модулируется методами амплитудной модуляции (AM), а звук — частотно-модулированным (FM).
Аналогичные соображения применимы при рассмотрении сигналов NTSC, где ширина полосы вещания составляет около 4,2 МГц.
В системах видеонаблюдения, в большинстве систем , у нас нет таких ограничений полосы пропускания , потому что мы не передаем видеосигнал с радиочастотной модуляцией. Нам не нужно беспокоиться о помехах между соседними видеоканалами.В видеонаблюдении мы используем необработанный видеосигнал, выходящий из камеры, который представляет собой видео с базовой полосой пропускания , или обычно называется видео в основной полосе частот . Обычно обозначается аббревиатурой CVBS , , что означает композитный импульсный видеосигнал . Спектр такого сигнала, как уже говорилось, колеблется от 0 до 10 МГц в зависимости от качества источника.
Спектральная емкость коаксиального кабеля как среды передачи намного шире этой.Например, наиболее часто используемый коаксиальный кабель 75 Ом RG-59B / U может легко передавать сигналы с полосой пропускания до 100 МГц. Конечно, это применимо к ограниченному расстоянию в пару сотен метров, но этого достаточно для большинства систем видеонаблюдения. Различные среды передачи подразумевают разные ограничения полосы пропускания, некоторые из которых шире, а некоторые уже, чем коаксиальный, но большинство из них значительно шире 10 МГц.
Bayonet Neil-Concelman (BNC) — самый распространенный входной композитный видеоразъем в системах видеонаблюдения.
Временная диаграмма телевизионного сигнала NTSC
Временная диаграмма телевизионного сигнала PAL
Frontiers | Интеграция электрических сигналов, исходящих из корня сосудистых растений
Электрическая природа жизни
Набухание под действием давления — это проблема, которая возникла на ранней стадии эволюции и решена с появлением мембранных белков, позволяющих синхронное перераспределение ионных градиентов через плазматическую мембрану. Вторичным эффектом этого решения является генерация падения напряжения внутри мембранного диэлектрика (Финкельштейн, 1976; Армстронг, 2015).Активность ионных каналов и переносчиков выборочно регулирует прохождение ионов, создавая временные локальные изменения мембранного потенциала при включении метаболитов или изменении проницаемости мембраны в ответ на внешний сигнал. Учитывая синхронизацию клеточных процессов и коммуникации внутри клеточных сообществ, электрическое зондирование и передача сигналов развиваются как широко распространенный механизм на разных уровнях биологической организации. Из бактериальных биопленок (Strahl, Hamoen, 2010; Masi, Ciszak, 2014; Prindle et al., 2015) высшим растениям (Sanderson, 1872; Darwin, 1897; Bose, 1907; Pickard, 1973) и животным (Galvani, 1791; Hodgkin, 1937; Cole and Curtis, 1939; Armstrong, 2007). по своей сложности от простых постепенных или осциллирующих изменений мембранного напряжения до передачи электрических сигналов на большие расстояния, наблюдаемых в возбудимых клетках.
Электрические сигналы у животных и высших растений
Хорошо известно, что и растения, и животные используют электрические сигналы дальнего действия для передачи информации об окружающей среде всему телу (Armstrong, 2007; Hedrich et al., 2016). В многоклеточных организмах информация должна передаваться от детекторов к эффекторной ткани. В случае животных нервная система играет центральную роль в гомеостазе, выступая в качестве основного интегратора большей части важной физиологической информации. Связь между эпителиальной тканью и возбудимыми клетками определяет способ взаимодействия животных с окружающей средой не только за счет использования сенсорных модальностей, таких как прикосновение, температура, свет или звук (Frings, 2009; Julius and Nathans, 2012), но также путем интеграции внутренние процессы, такие как гормональные выделения, физиология кишечника и развитие иммунной системы (Zhang and Zhang, 2009; Bellono et al., 2017; Клемменсен и др., 2017).
Молекулярные детекторы, обнаруженные в сенсорном эпителии, активируются сигналами окружающей среды, вызывая (прямо или косвенно) открытие проводимости ионного канала, которая изменяет локальный трансмембранный потенциал (Martinac, 2008). В невозбудимых клетках, таких как эпителиальные клетки кишечника или легких, электрогенный транспорт регулирует поглощение питательных веществ, контролирует pH и модулирует секрецию воды (Boyd, 2008; Beumer and Clevers, 2017; Clemmensen et al., 2017).В случае эпителия отсутствие подходящих зависимых от напряжения каналов (т.е. Ca v , Na v ) препятствует распространению начальной деполяризации на большие расстояния. В возбудимых клетках деполяризация обеспечивает необходимую энергию для открытия потенциал-управляемых каналов (например, Ca v s, Na v s и K v s) (Bezanilla, 2008; Catterall et al., 2017). Скорость распространения, форма распространяемого потенциала и частота электрического сообщения определяются свойствами кабеля ячейки, которые определяются как геометрией каждого конкретного типа ячейки, так и доступным набором ионных каналов.У животных этот тип коммуникации распространяется на многоклеточный организм, когда вещество, высвобожденное из данной клетки, оказывает влияние на постсинаптическую клетку (Gerber and Südhof, 2002; Jackson, 2006; Catterall and Few, 2008). Как было первоначально описано в Aplysia E. Kandel (Castellucci and Kandel, 1976), возбудимые клетки изменяют свое поведение в ответ на стимуляцию. Учитывая, что контроль экспрессии, локализации и активности клеточных рецепторов и ионных каналов представляет собой молекулярную основу неассоциативного обучения у животных (Kandel, 2001), возникает соблазн задать вопрос, как растения модулируют различные потоки ионов и какие именно элементы, придающие пластичность обучению растений.
Сообщалось о потенциале действия
у водорослей и высших растений (Pickard, 1973; Trebacz, Zawadzki, 1985; Kateriya et al., 2004; Fromm and Lautner, 2007; Hegenauer et al., 2016). Как предвидел Дэвис (1987), в настоящее время широко признано, что электрические сигналы играют важную роль в меж- и внутриклеточной коммуникации растений. Однако, в отличие от детального знания молекулярных и клеточных механизмов, управляющих передачей электрических сигналов у животных, идентичность клеточных сенсоров и эффекторов и их точное распределение в растении до сих пор неясны (Ward et al., 2009; Хедрич, 2012; Hedrich et al., 2016). Более того, не имея сложной сотовой проводки, разработанной многоклеточными животными для передачи своих электрических сигналов на большие расстояния, кажется, что растения разработали архитектуру, позволяющую им формировать — или заставляя их адаптировать — электрическую связь по-другому. Из этого рассуждения возникают простые вопросы: как именно растения подключаются к сети? Как электрические сигналы проходят через сотовую сеть? Как эти сигналы работают вместе, связывая зондирование окружающей среды, экспрессию генов, поглощение питательных веществ, газообмен, водный баланс, производство энергии и хранение отходов? В этом обзоре мы не стремимся ответить на такие амбициозные вопросы, а скорее рассматриваем различные элементы, которые могут способствовать генерации и распространению электрического сообщения в корнях наземных растений.
Способность ориентироваться является атрибутом животных и создает фундаментальные проблемы для решения, такие как (i) мультиплексирование сенсорного ввода на высоких частотах, (ii) быстрая интеграция этих сигналов и (iii) передача вычисленной команды с помощью изысканной сотовой связи. точность, обеспечивающая согласованный отклик организма. Одноклеточные зеленые водоросли, предки наземных растений, обладают навигационными способностями и по совпадению представляют другой набор ионных каналов по сравнению с их потомками, выражая важные элементы, важные для формирования электрического ответа возбудимых клеток у животных (Merchant et al., 2007; Уиллер и Браунли, 2008 г.). К ним относятся активируемые напряжением кальциевые и натриевые каналы, каналы TRP и рецептор рианодина, которые отсутствуют у современных наземных растений (Wheeler, Brownlee, 2008; Ward et al., 2009; Fromm and Lautner, 2012; Taylor et al., 2012; Arias-Darraz et al., 2015; Edel et al., 2017). Находящиеся в ловушке одного и того же природного мира животные и растения разделяют большой набор факторов экологического стресса. Тем не менее, они явно приняли разные способы решения основных проблем, таких как воспроизводство и самосохранение.Вероятно, поиск пищи, спаривание и потребность в утилизации отходов заставили животных разработать сигнальные механизмы, настроенные на навигацию. С другой стороны, растения не только производят собственные углеводы, но и, что немаловажно, хранят их отходы. Следовательно, сидячая природа наземных растений требует вместо этого надежных механизмов адаптации. Соответственно, клеточные и молекулярные сенсоры постоянно снабжают растение полезной экологической информацией, которая должна распространяться по всему телу (Karban, 2015).Недавние исследования показывают, что Arabidopsis эффективно организуют свое трехмерное планирование для оптимизации подачи питательных веществ (Conn et al., 2017), укрепляя идею о том, что архитектура растения контролируется механизмом управления, отвечающим за обмен между общей длиной ветвей и распределением питательных веществ. . Такой механизм должен быть связан с природой и свойствами распространения электрических сигналов, генерируемых в корне и листьях, тканях, в которых абсорбируются минералы и вода, производятся углеводы и хранятся побочные продукты.Дальнейшая экспериментальная работа с интактными живыми растениями с использованием подходящих моделей, позволяющих одновременную электрическую и визуализационную регистрацию, необходима для оценки влияния электрических сигналов на распределение пищи по растению (Kanchiswamy et al., 2014; Salvador-Recatalà et al., 2014; Gunsé et al., 2016; Candeo et al., 2017).
Проводящий завод
Отсутствует не только клеточная архитектура, но и набор ионных каналов, кодирующих электрические сообщения у животных (Ward et al., 2009; Hedrich, 2012), нет оснований предполагать, что сенсорная информация в растениях интегрируется или обрабатывается аналогичным образом. Было высказано предположение, что флоэма растений образует один проводящий кабель, эквивалент аксона в одном нейроне многоклеточных животных (Hedrich et al., 2016). Различные типы клеток, включая клетки-компаньоны и элементы сита, образуют флоэму. В отличие от других типов растительных клеток, клетки ситовых элементов не имеют неоднородностей в своей проницаемости из-за наличия ситчатых пластин, что обеспечивает непрерывный перенос растворенных веществ между различными органами растения и обеспечивает канал с низким сопротивлением и высокой емкостью, который позволяет распространение относительно медленных электрических сигналов.Десятилетия теоретических и экспериментальных данных выдвинули идею о том, что флоэма будет основным каналом, способным электрически соединять корни и воздушные ткани (Brenner et al., 2006; Fromm et al., 2013; Hedrich et al., 2016). Тем не менее, информация, обнаруженная в эпидермальных клетках корня, должна распространяться через клеточную сеть коры, интегрироваться и достигать флоэмы, чтобы трансдуктироваться по всему телу растения. И наоборот, сигнал должен выходить из флоэмы, чтобы воздействовать на клетки надземной ткани.Для выполнения этой сложной задачи сосудистые растения имеют взаимосвязанное внеклеточное пространство между плазматической мембраной и клеточной стенкой (то есть апопластическое пространство) и прямую клеточную связь через плазмодесматы (Sattelmacher and Horst, 2007; Lee, 2015). Эти особенности служат для различных сигнальных функций в растении, позволяя не только передавать растворимые сигналы, но также определять электрическую связь между клетками и модуляцию специфических сигналов, связанных с кальциевым ответом, который сопровождается обнаружением различных сигналов окружающей среды (Zebelo и другие., 2012; Nawrath et al., 2013; Ли, 2015; Choi et al., 2016; Edel et al., 2017).
У растений описаны два основных типа электрических сигналов на большие расстояния: потенциалы действия (AP) и потенциалы вариации (VP) (Bose, 1907; Pickard, 1973; Fromm and Lautner, 2007, 2012). Первые индуцируются деполяризацией напряжения, обладают пороговым потенциалом, следуют принципу « все или ничего » и перемещаются с постоянной скоростью и амплитудой, очень похоже на AP, наблюдаемые в животном мире (Zawadzki et al., 1991; Джексон, 2006; Армстронг, 2007; Ян и др., 2016). Напротив, VPs, как было показано, вызываются быстрым увеличением внутреннего давления ксилемы и проявляются как медленные волны деполяризации переменного размера (Fromm and Lautner, 2007, 2012). Также была описана третья мода электрического сигнала, названного системными потенциалами (SP), состоящая из гиперполяризации, которая распространяется на средние расстояния (Zimmermann et al., 2009). В то время как VP зависят от инактивации P-типа H + -ATPase, SP, по-видимому, вызваны активацией насоса.Из ранних работ Бурдона-Сандерсона известно, что время нарастания для AP растений составляет порядка 0,1 с, с длительностью около 1 с и скоростью распространения порядка нескольких сотен мм с −1 (Сандерсон, 1872; Пикард, 1973). Эти электрические сигналы различаются не только по форме и величине, но и по скорости распространения от 1 до 60 мм с -1 для AP до нескольких минут на сантиметр для VP. Системные потенциалы запускаются деполяризацией, не имеют характера «все или ничего», самораспространяются с постоянной скоростью около 0.5–2 мм с –1 , а их величина пропорциональна входным стимулам (Zimmermann et al., 2009). Интересно, что SP напоминают рецепторные потенциалы животных, одновременно самораспространяясь по сенсорному эпителию. Лист арабидопсиса, бобов и ячменя проявляет самораспространяющуюся электрическую активность, вызванную ранением, ограниченную коммуникацией от листа к листу и связанную с экспрессией генов, подобных рецепторам глутамата (Zimmermann et al., 2009; Mousavi et al. др., 2013; Сальвадор-Рекатала и др., 2014; Сальвадор-Рекатала, 2016а). В этом случае тип раны, по-видимому, связан с различными типами деполяризации. Было высказано предположение, что анатомия ткани будет иметь значение для определения связи между поверхностной тканью и флоэмой (Salvador-Recatalà, 2016a).
Тем не менее, было сложно систематизировать как теоретическую модель, объединяющую электрическую сигнализацию всего предприятия, так и экспериментальные методы изучения электрической связи на большие расстояния в конфигурации всего предприятия (Goldsworthy, 1983; Davies, 1987; Pietruszka et al., 1997; Фромм и Лотнер, 2007; Волков, 2012; Fromm et al., 2013; Hedrich et al., 2016). Тем не менее было установлено, что различные органы растения, включая листья, стебель, цветы и корень, обладают внутренней электрической активностью (Pickard, 1973; Baldwin et al., 2006; Fromm and Lautner, 2007, 2012; Appel and Cocroft). , 2014; Engineer et al., 2015; Karban, 2015; Zhou et al., 2016). Более того, была описана электрическая связь на больших расстояниях между корнями, побегами и листьями (подробно рассмотрено в Pickard, 1973; Fromm and Lautner, 2007; Zimmermann et al., 2009; Hedrich et al., 2016). Подробное описание передачи электрического сигнала на корнях отсутствует, вероятно, из-за кажущейся нескоординированной природы точек доступа корня (Fromm and Eschrich, 1993; Fromm et al., 1997, 2013; Masi et al., 2015; Salvador-Recatalà, 2016b). ).
Картирование набора ионных каналов у Arabidopsis
Электрические свойства клеток проистекают из экспрессии и контроля ионных каналов, переносчиков и насосов. Они могут модулироваться различными стимулами, такими как давление, экзогенные и эндогенные лиганды, температура, свет, напряжение на мембране и растяжение среди других.Молекулярный механизм, определяющий распространение электрических сигналов в растениях, не известен в деталях, но, принимая во внимание экспериментальные данные и доступную генетическую информацию, мы узнали, что растения и животные используют разные стратегии для распространения AP. В то время как животные используют чувствительные к напряжению каналы Na + и Ca 2+ для деполяризации (Hodgkin and Huxley, 1952; Armstrong, 2007; Catterall et al., 2017), токсическая природа натрия заставляет клетки растений использовать Cl — и Ca 2+ вместо.В то время как Ca 2+ вызывает деполяризацию, попадая в клетку, Cl — вызывает деполяризацию, покидая клетку. Согласно профилям экспрессии генов, деполяризация растительных клеток, вероятно, вызвана хлоридными каналами ALMT / QUAC-типа и / или ионными каналами, обеспечивающими приток кальция, такими как двухпоровые каналы (TPC), циклические нуклеотидно-управляемые каналы (CNGC) или глутаматные рецептороподобные каналы (GLR) (Ward et al., 2009; Hedrich, 2012; Hedrich et al., 2016). В цепи событий, определяющих AP, начальное повышение Ca 2+ вызовет отток Cl —, и последующая активация зависимых от напряжения калиевых каналов, вероятно, будет участвовать в реполяризации (Schroeder et al., 1984; Ward et al., 2009; Hedrich et al., 2016).
Поскольку способность ткани генерировать электрические сигналы будет определяться набором ионных каналов, выраженных в различных типах клеток, участвующих в передаче электрического сообщения, мы нанесли на карту функционально охарактеризованные каналы и транспортеры, которые ранее были связаны с передачей электрических сигналов. у Arabidopsis thalina (Barbier-Brygoo et al., 2011; Hedrich, 2012) (таблица 1). Мы провели иерархический кластерный анализ, чтобы сгруппировать эти гены в соответствии с профилями экспрессии, полученными из EPlant (Waese et al., 2017). При сравнении всех соответствующих тканей на разных стадиях развития мы наблюдали, что разные ионные каналы представляют собой характерный образец экспрессии (рис. 1А).
Таблица 1 . Функциональные ионные каналы арабидопсиса.
Рисунок 1 . Картирование экспрессии ионных каналов в Arabidopsis thaliana . (A) Иерархическая кластеризация нормализованных уровней мРНК генов, кодирующих калиевый (черный), кальциевый (красный) и анионный (синий) каналы.Данные микрочипов были получены и визуализированы в браузере Arabidopsis eFP (Waese et al., 2017). Значения экспрессии для каждого гена были преобразованы в Z-баллы для всех образцов, чтобы идентифицировать тканеспецифическую экспрессию. Высокие значения Z-показателя (красный или синий) указывают на большее отклонение от среднего выражения по всем тканям. (B) Примеры ионных каналов, демонстрирующие тканеспецифическую экспрессию.
Известно, что передача сигналов кальция важна для контроля открытия устьиц (Laanemets et al., 2013), а калиевые каналы имеют решающее значение в фазе реполяризации (Schroeder et al., 1984). Как и ожидалось, мы наблюдали большую экспрессию GLR и каналов K v в листьях (Фигуры 1A, B). Хотя гены, кодирующие анионные и калиевые каналы, не показывают четкого разделения между надземной частью и корнями (рис. 1A), мы обнаружили специфические гены, экспрессия которых преобладает в корнях (VDAC1), стеблях (GLR3.2) или листьях ( GLR3.3) (Рисунок 1B). Эти тканеспецифические профили экспрессии предполагают, что существуют разные пути генерации и распространения электрических сигналов в растениях и что эти пути меняются во время развития.При более подробном рассмотрении два корневых анионных канала, SLAh4 и VDAC1, показали различный паттерн экспрессии для разных типов клеток (Рис. 2). Зависимый от напряжения анионный канал VDAC1 сильно экспрессируется в ткани корня. Для сравнения, экспрессия в меристематической зоне выше, чем в корневых волосках (рис. 2). Точно так же H + -АТФаза экспрессируется почти во всех типах клеток корня. Напротив, канал SLAh4 с медленной проводимостью хлоридов показал большую экспрессию во внутренних тканях корня, таких как перицикл и кора головного мозга, важных физических барьерах на пути к флоэме (Nawrath et al., 2013). В то время как экспрессия электрогенного транспортера нитрата NRT1.1 преимущественно наблюдается в корневых волосках и во флоэме ближе к стеблю, вакуолярный канал TPC1 заметно экспрессируется в корневых волосках зоны созревания. Интересно, что ни один из этих мембранных белков не обнаружил заметной экспрессии во флоэме вдоль корня (рис. 2).
Рисунок 2 . Сравнительные профили экспрессии при разрешении по типу клеток анионных каналов и генов, связанных с захватом нитратов в корне Arabidopsis.Данные микрочипов были получены и визуализированы с помощью браузера Arabidopsis eFP (Waese et al., 2017).
Приток кальция в ответ на внешние раздражители имеет решающее значение для генерации электрического сигнала. С притоком Ca 2+ у наземных растений связано около 50 различных ионных каналов. Этот большой набор кальциевых каналов разделен на пять различных семейств: CNGC, GLR, TPC1, каналы, связанные с осмотическим ответом (OSCA), и механочувствительные кальциевые каналы (MCA) (Kurusu et al., 2012; Чин и др., 2013; Морган и Галионе, 2014 г .; Edel et al., 2017) (таблица 2). Из этих каналов CNGC14, CNGC19, GluR2.1 и OSCA1.4, по-видимому, преимущественно экспрессируются в ткани корня (рис. 3А). Экспрессия этих каналов также была дифференциальной. CNGC14 в значительной степени экспрессируется в эпителии вблизи меристематической зоны и в меньшей степени в зоне созревания. Напротив, вакуолярный канал CNGC19 сосредоточен в эндотелии и флоэме. С другой стороны, GLR2 закрывается лигандом.1 и осмотические каналы OSCA1.4 преимущественно экспрессируются в корневых волосках. В то время как экспрессия GLR2.1 в эпителиальной ткани несколько снижается от меристематической зоны к зоне созревания, OSCA1.4 преимущественно экспрессируется в зоне созревания и флоэме (Figure 3B).
Таблица 2 . Кальциевые каналы у арабидопсиса.
Рисунок 3 . Картирование экспрессии кальциевых каналов в Arabidopsis thaliana . (A) Тепловая карта и кластерные дендрограммы экспрессии кальциевых каналов среди типов тканей.Образцы тканей представлены столбцами, а гены — строками. Значения экспрессии для каждого гена были преобразованы в Z-баллы для всех образцов, чтобы идентифицировать корне-специфическую экспрессию, которая обозначена черными прямоугольниками. (B) Кальциевые каналы, показывающие экспрессию корня.
Обработка нитратами растений, испытывающих нехватку азота, вызывает временную деполяризацию плазматической мембраны (Meharg and Blatt, 1995; Wang and Crawford, 1996; Wang et al., 1998). Аналогичным образом, недавно сообщалось, что лечение нитратами вызывает повышение внутриклеточного кальция, что инициирует путь передачи сигналов нитратами (Liu et al., 2017). Более того, генетические данные показывают, что повышение внутриклеточного кальция связано с NRT1.1. (Риверас и др., 2015). Учитывая наблюдаемый профиль экспрессии, мы можем предположить, что деполяризация эпителиальной клетки, вызванная поглощением нитратов, вызванная увеличением активности транспортера нитратов NRT1.1, будет запускать приток кальция через OSCA1.4 и / или GLR2.1 , дополнительно активируя кальций или зависимую от напряжения хлоридную проводимость (например, SLAh4 или VDAC1), обеспечивая распространение электрического сигнала по коре к флоэме (рис. 4).
Рисунок 4 . Схематическое изображение корня растения, показывающее различные пути распространения электрических сигналов. Различное проводящее состояние плазмодесмы A, B и C будет модулировать сигнал, обнаруживаемый в корневом волоске на его пути к ситовой трубке. На верхней левой панели показаны проводимости, которые могут быть связаны с деполяризацией, вызванной нитратами. Увеличение содержания кальция в эпителиальных клетках модулирует активность плазмодесмы. И уровень кальция, и физические изменения плазмодесмы могут модулировать активность ионных каналов, экспрессируемых в этой области.Верхняя правая панель изображает цитоплазматический и ретикулярный пути для ионов, оба они могут модулироваться взаимодействием с заякоренными белками в месте контакта поры.
Сотовая связь как часть электрической сигнализации предприятия
Единая модель, объединяющая восприятие растений и контролируемое поведение, необходима для объяснения того, передают ли стимулы, обнаруженные на границах тела растения (например, корневые волоски, эпителиальные клетки листьев) электрическую информацию в кабель флоэмы, как далее интегрируется, и, наконец, как электрический сигнал выходит из флоэмы, достигая эффекторной ткани, расположенной в отдаленном эпителии (например,g., замыкающие клетки в устьице листа). Хотя дифференциальное выражение набора ионных каналов важно для определения возбудимости, взаимосвязанная клеточная архитектура растения будет иметь решающее значение для управления характеристиками амплитуды и распространения в трехмерном пространстве. Модуляция этих элементов в ответ на повторяющиеся, острые или хронические сенсорные воздействия может придать пластичность реакции растения и позволит как обучение, так и адаптацию без необходимости в «мозговом» интеграторе или когнитивном поведении, как это предлагается в литературе ( Baluska et al., 2004).
Одним из основных структурных различий между клетками животных и растений является наличие клеточной стенки. Стенка растительной клетки создает необычную внеклеточную среду, известную как апопласт, которая представляет собой физический / химический барьер, который участвует в путях межклеточной коммуникации, включая передачу электрических сигналов на большие расстояния (Sattelmacher, 2001; Fromm and Lautner, 2007; Choi et al. ., 2017). У растений внеклеточная концентрация ионов соответствует апопластным ионным концентрациям, которые строго регулируются.Ионный состав апопласта варьируется и зависит как от внутренних факторов, таких как ткань или стадия развития, так и от внешних факторов, таких как пищевой стресс (López-Millán et al., 2001). Ионы в апопластном компартменте обычно находятся в низких концентрациях, состоящих преимущественно из неорганических катионов и анионов, таких как K + , Ca 2+ , Mg 2+ , Cl —, NO3- и PO43- (Gabriel и Кессельмейер, 1999). Концентрации апопластных и внутриклеточных ионов вместе с ионной проницаемостью будут определять потенциал покоя эпидермальной клетки корня и, как сообщается, составляют около -120 мВ, отрицательные внутри (Fromm and Eschrich, 1993).Биохимические свойства клеточной стенки в первую очередь будут определять апопластическое пространство и проницаемость для питательных веществ. У взрослого растения высокое содержание лигнина или суберина в клеточных стенках значительно снижает их проницаемость (Nawrath et al., 2013) и, следовательно, представляет собой плотный барьер для свободной диффузии питательных веществ в ткань растения и из нее. Напротив, структура и состав клеточных стенок изменяются во время развития корня (Somssich et al., 2016). Меристематическая зона содержит молодые клетки, расположенные близко к верхушке корня, тогда как более старые клетки располагаются у основания корня близко к стеблю.Клеточные стенки меристематической зоны тонкие и более проницаемые из-за более высокой митотической активности (Baluska et al., 1996). Клетки из зоны дифференцировки более жесткие из-за накопления лигнина, связанного с развитием вторичных клеточных стенок, что обеспечивает дополнительную прочность стенкам и делает их водонепроницаемыми. Somssich et al. (2016). Следовательно, в первичных корнях существует продольный градиент проницаемости, определяемый степенью дифференцировки эпидермальных клеток.
Равновесная концентрация ионов может быть рассчитана из уравнения Нернста вида [X] out / [X] in = exp (V m ZF / RT), было [X] out и [ X] в — внешняя и внутренняя концентрации иона X, V m — напряжение на мембране, Z — валентность проницаемого иона, F — постоянная Фарадея, R — газовая постоянная и T — абсолютная температура.Из этого уравнения ясно, что изменения внеклеточной концентрации проницаемого заряженного растворенного вещества будут влиять на мембранный потенциал в состоянии покоя. Фактически, ранние исследования показали, что как пик потенциала действия, так и величина входящего тока зависят от концентрации кальция в апопласте (Hope, 1961a, b; Hope and Findlay, 1964). С другой стороны, активность H + -АТФазы поддерживает апопластный pH, обычно кислый (4,7–5) (Sattelmacher and Horst, 2007).Поскольку мембранный потенциал покоя в растительных клетках, скорее всего, устанавливается за счет транспорта протонов, поэтому следует ожидать высокой чувствительности к вариациям активности протонного насоса (Brault et al., 2004). Учитывая, что апопласт занимает относительно небольшой объем тканей растения, составляющий менее 5% в случае листьев (López-Millán et al., 2001), разумно ожидать, что изменения окружающей среды могут вызвать быстрое защелачивание апопласта, которое в свою очередь вызовет локальную деполяризацию (Grams et al., 2009). Анионные питательные вещества, такие как нитрат, сульфат и фосфат, пассивно приобретаются посредством электрогенного совместного транспорта, которому способствует протонный градиент (Ullrich-Eberius et al., 1981; Muchhal and Raghothama, 1999). Точно так же апопластный pH будет чувствителен к активности переносчиков фосфатов и нитратов (Amtmann et al., 1999). Соответственно, острое лечение нитратами растений, испытывающих нехватку азота, вызывает временную деполяризацию плазматической мембраны (Meharg and Blatt, 1995; Wang and Crawford, 1996).Экспериментальные данные показывают, что 2–4 протона совместно транспортируются во время поглощения фосфата (Sakano, 1990). В случае нитрата стехиометрия совместного транспорта с протонами была рассчитана как 2: 1 (McClure et al., 1990; Glass et al., 1992; Wang and Crawford, 1996). Принимая во внимание экспериментальные данные исследований in vitro с мембранными везикулами, а также генетические анализы, такие как комплементация дрожжей, наиболее вероятной стехиометрией совместного транспорта протонов и сульфатов будет 3: 1 (Buchner et al., 2004).
Корень — это первый орган, который контактирует с водой и питательными веществами. Поэтому, когда растения лишены питательных веществ, корень является основным местом для обнаружения. Сравнительный транскриптомный анализ образцов корней и побегов показал, что реакция растения на нитраты инициируется в корне (Wang et al., 2003). Растения должны интегрировать информацию из внешней среды и противопоставлять ее своему питательному статусу. Фактически, сообщалось, что баланс между азотом и углеродом важен для контроля усвоения азота (Zheng, 2009).Следовательно, растениям требуется эффективная система связи между местом восприятия и поглощения питательных веществ (то есть корнями) и метаболическим центром, где вырабатывается углерод (то есть листьями), чтобы адекватно реагировать на изменения в доступности питательных веществ. Эпидермальные клетки корня растения вызывают более высокую проницаемость через клеточную стенку, заставляя электрогенные переносчики на плазматической мембране сталкиваться с высокими концентрациями питательных веществ, когда они растворяются в почве, окружающей корень. Поглощение любых этих питательных веществ вызовет быструю и временную деполяризацию мембраны эпителиальных клеток в корне (Dunlop and Gradiner, 1993; Meharg and Blatt, 1995).Более того, диффузия веществ внутри апопласта корня внутренне ограничена полосой Каспариана, гидрофобной пропиткой лигнина первичной клеточной стенки, которая герметизирует внеклеточное пространство энтодермальных клеток, заставляя прохождение ионов, питательных веществ и воды через них. плазматическая мембрана эндотелиальных клеток (Nawrath et al., 2013; von Wangenheim et al., 2017) (Рисунок 4).
Помимо апопластического пути коммуникации во внеклеточном пространстве, цитоплазма растительных клеток может быть внутренне связана межклеточными соединениями, известными как плазмодесма (Lee, 2015; Kitagawa and Jackson, 2017).Межклеточная коммуникация тканей корня была продемонстрирована быстрой диффузией флуоресцентных молекул-индикаторов (например, пропидия йодида) через плазмодесмы во внутренние слои ткани корня (Nawrath et al., 2013). Было продемонстрировано, что недифференцированные клетки из апикальной корневой меристемы и зоны элонгации связаны с красителем и, следовательно, симпластически связаны посредством плазмодесм (Duckett et al., 1994). Ионы и более крупные молекулы могут свободно диффундировать через поры от одной клетки к другой, делая эти клеточные структуры фокусом передачи сигналов через ткань коры (Burch-Smith and Zambryski, 2012; Lee, 2015).Более того, сложность плазмодесм подчеркивается наличием эндоплазматического ретикулума (ER), проходящего через поры, обеспечивая вторичный и, вероятно, более избирательный путь коммуникации между соседними клетками. Известно, что различные липиды и каллоза , растворимый белок, способный перекрывать цитоплазматический путь, регулируют проникновение через плазмодесмы (Tilsner et al., 2016). Более того, сообщалось, что повышение содержания кальция в цитоплазме способствует закрытию цитоплазматического пути поры (Lee, 2015).Дополнительные данные необходимы, чтобы определить вклад этих ER-трубок в распространение как кальциевых, так и электрических сигналов через кору корня.
У растений прямая связь между ER и электрической активностью на плазматической мембране не связана со стромальной взаимодействующей молекулой 1 (STIM1) или с кальциевыми каналами Orai1, как в клетках животных. Связанные с STIM белки были потеряны на уровне одноклеточных водорослей, а родственники Orai присутствуют только до голосеменных (Edel et al., 2017). Тем не менее, было высказано предположение, что закрепляющие белки, элементы цитоскелета и липиды могут модулировать локализацию и активность мембранных белков в месте контакта между ER и плазматической мембраной (Lee, 2015). Таким образом, дифференциальная экспрессия и модуляция определенного набора кальций-чувствительных ионных каналов, окклюзия плазмодесм или изменение формы поры в месте контакта могут обеспечивать усиление или подавление распространяющегося электрического сигнала через кору (Рисунок 4).
Дифференцировка эпидермальных клеток корня у Arabidopsis постепенно снижает эти цитоплазматические связи таким образом, что они становятся симпластически разобщенными на последней стадии своего развития (Duckett et al., 1994). Напротив, клетки эпидермиса гипокотиля симпластически связаны друг с другом независимо от состояния их развития (Duckett et al., 1994). Более ранние доказательства электрического взаимодействия были предоставлены Спансвиком и Костертоном, которые показали, что при подаче тока в клетку многоклеточной водоросли Nitella сигнал можно было отследить на расстоянии нескольких клеток от места инъекции (Spanswick and Costerton, 1967).
Недавние эксперименты по визуализации кальция с использованием генетически закодированного ратиометрического индикатора кальция (например, Y-Cameleon 3.6), экспрессируемого в живом арабидопсисе, показали, как спонтанная реакция, возникающая в корневых волосках, распространяется через корневую ткань четко определенными «пятнами» (Candeo и др., 2017). Хотя авторы не комментировали этот шаблонный ответ, по аналогии с сенсорными рецепторами у животных, возникает соблазн интерпретировать такие показания как физическое измерение «сенсорного поля , », которое соответствует конкретной эпителиальной клетке или их группе. .После стимуляции клетки корневого эпидермиса генерируется электрический сигнал; в приведенном выше примере приток кальция может быть напрямую связан с деполяризацией клеток. Генерируемый электрический сигнал может передаваться через плазмодесмы к соседним связанным клеткам корня корня. Как только сигнал достигает ситовой трубки с низким сопротивлением во флоэме, он распространяется по всему растению (рис. 4). Тогда возникает вопрос, интегрирует ли флоэма множественные сигналы, поступающие от отдельных рецепторных полей, рассредоточенных по эпителию корня, и как сеть межклеточных соединений может обеспечивать пластичность, модулируя распространение электрического сообщения, контролируя локализацию и активность плазмодесмат. (Рисунок 4).Более того, диффузионные эксперименты с флуоресцентными красителями показали, что связь между эпидермальными клетками гипокотиля заканчивается у основания ствола (Duckett et al., 1994). Следовательно, вероятно, что эпидермальные клетки гипокотиля и корня электрически не связаны, что делает ситчатые трубки единственным возможным путем распространения AP от корня к побегу. Все эти физические барьеры создают узлы резистивных элементов, полезных для фильтрации сигналов не только на выходе из ткани корня, но и в ветвях, выходящих из стебля, и из них.
Было высказано предположение, что растения могут интегрировать информацию через структуру «, подобную мозгу », расположенную в верхушке корня (Baluska et al., 2004; Brenner et al., 2006). Однако поразительные особенности мозга животных проистекают не только из того факта, что нейроны устанавливают синапсы, но и из способности создавать сложные логические схемы на основе клеток. Один нейрон может содержать сотни специфичных для клеток синапсов, каждый из которых способен к пластической адаптации. В отсутствие такого клеточного взаимодействия маловероятно, чтобы интеграция электрической информации во всем растении привела к когнитивной обработке сенсорной информации.Более того, недавние работы по одноклеточным организмам, таким как слизистая плесень и бактерии, демонстрируют, что центральной нервной системе нет необходимости порождать интеллектуальные решения (Nakagaki, 2001; Kotula et al., 2014).
Представляя очень упрощенную картину электрических соединений растений, мы видим, что они состоят из очень неэффективного кабеля, окруженного эпителиальной тканью, образованной абсорбирующими и возбудимыми клетками. В такой модели эпителиальные волосковые клетки корня могут функционировать как шипы в нейроне, передавая сигналы, генерируемые внешними стимулами, в контур коры, где они обрабатываются до достижения флоэмы.В то же время переход от корня к ножке может обеспечить дополнительный фильтр, действующий как макроскопическая версия шейки позвоночника, модулирующая прохождение электрических сигналов дальнего действия. Количество клеток в коре, которые электрически связаны, определяет размер конденсатора цепи и обрисовывает электрические свойства путей, достигающих флоэмы. Присутствие гипотетических «синапсоподобных» структур на основе актина может способствовать формированию сети межклеточных соединений (Baluška et al., 2005). В этом контексте системные растворимые сигналы, такие как ауксины, также могут играть важную роль в регуляции межклеточных соединений, например, посредством взаимодействия с плазмодесмами (Baluska et al., 2004; Brenner et al., 2006). Кроме того, сенсорный ввод может также влиять на фундаментальные свойства кабеля вдоль канала флоэмы, формируя и фильтруя распространяемые потенциалы действия. Природа физических / молекулярных барьеров (например, переход от коры корня к флоэме и эпителия от флоэмы к листу) будет иметь большое значение для понимания интеграции сенсорной информации в растениях.
Определение питательных веществ и передача электрических сигналов на большие расстояния
Растения получают необходимые химические компоненты для синтеза биомолекул из минералов, присутствующих в почве (Maathuis, 2009). Фосфор, азот, сера, калий, магний и кальций являются питательными веществами, которые в больших количествах требуются высшим растениям (Kirkby, 2011). Следовательно, растения должны постоянно усваивать эти питательные вещества, чтобы обеспечить подходящий рост и развитие (Maathuis, 2009). В отличие от гетеротрофных организмов, растения в основном получали питательные вещества в неорганической форме с помощью специфических переносчиков, локализованных в корнях.Экспрессия этих переносчиков питательных веществ, а также их активность регулируются доступностью питательных веществ, метаболизмом и факторами окружающей среды (Giehl and von Wirén, 2014). Недостаток одного из этих незаменимых питательных веществ оказывает прямое влияние на рост и развитие растений, особенно в случае корневой ткани (Gruber et al., 2013). Следовательно, растения разработали сложные регуляторные системы для обеспечения поглощения этих неорганических питательных веществ (Schachtman and Shin, 2007). Фактически, реакция на недостаток питательных веществ включает сложные сигнальные сети, включая сенсорные белки (Ho et al., 2009), факторы транскрипции (Rubio et al., 2001), миРНК (Vidal et al., 2010), пептиды (Ohkubo et al., 2017) и фитогормоны (Kiba et al., 2011). Интересно, что эти сигнальные пути не только запускают краткосрочные реакции, включающие метаболические корректировки и / или регуляцию переносчиков питательных веществ, но также вызывают модификации архитектуры корневой системы.
Среди макроэлементов фосфор и азот оказывают большее влияние на структуру корня по сравнению с другими (Gruber et al., 2013). Фосфатное голодание сильно стимулирует развитие корневых волосков (Péret et al., 2011), тогда как добавление нитратов вызывает увеличение плотности корневых волосков (Canales et al., 2017). Кроме того, дефицит фосфата вызывает значительное снижение роста первичных корней и стимулирует развитие боковых корней (Shahzad and Amtmann, 2017). Напротив, меньшая доступность нитратов увеличивает рост первичных корней и снижает развитие боковых корней. Эти противоположные эффекты на клеточную архитектуру корня предполагают наличие различных путей передачи сигналов для нитрата и фосфата, вероятно, связанных с электрическими сигналами разной природы, которые не обязательно распространяются одинаково или генерируются в одном и том же типе эпителиальных клеток.
Заключение
Изменения внутриклеточного кальция в корневых волосковых клетках эпидермиса растений генерируются механизмом, включающим локальные изменения мембранного потенциала, вызванные электрогенным транспортом или прямой активацией рецепторов растений внеклеточными лигандами. Эти локальные изменения мембранного напряжения будут обеспечивать усиление входного сигнала за счет увеличения кальциевой проницаемости эпителиальной клетки и дальнейшего запуска потенциалов действия за счет увеличения проницаемости проводимости хлоридов с последующей реполяризацией, управляемой калием / протонами.Эти электрические сигналы первоначально распространяются через клетки коры с помощью строго регулируемых сетей плазмодесм и, используя преимущества континуума апопластического пространства, до тех пор, пока они не достигнут ситовидной трубки флоэмы, где они получают доступ для передачи по всему телу растения. Физические барьеры, распределение ионных каналов и межклеточная связь в корне являются критическими аспектами, которые формируют электрические сигналы, генерируемые в сенсорной ткани. Все эти элементы находятся под клеточным контролем и могут обеспечивать пластичность реакции растений.Роль плазмодесм в распространении и настройке электрических сигналов в сенсорном эпителии растений является фундаментальной темой, которой несколько пренебрегают и, очевидно, требует большего внимания.
Взносы авторов
SB, JC и CH-V написали статью. SB и JC подготовили цифры.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
MiNICAD — это ядро тысячелетия, поддерживаемое Iniciativa Científica Milenio, Министерством экономики, развития и туризма Чили. Anillo Científico ACT-1401 поддерживает SB. SB является частью программы CISNe-UACh и UACh по клеточной биологии. JC поддерживается грантом FONDECYT 11150070. Институт интегративных систем и синтетической биологии «Миллениум» поддерживается Министерством экономики, развития и туризма Чили «Iniciativa Científica Milenio». Мы благодарим Шарлотту К.Коленсо за комментарии к рукописи.
Список литературы
Амтманн А., Джелитто Т. К. и Сандерс Д. (1999). K + -селективные внутренние каналы выпрямления и апопластный pH в корнях ячменя. Plant Physiol. 120, 331–338. DOI: 10.1104 / стр.120.1.331
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ариас-Дарраз, Л., Кабесас, Д., Коленсо, К. К., Алегрия-Аркос, М., Браво-Морага, Ф., Варас-Конча, И. и др. (2015). Транзиторный потенциальный ионный канал рецептора у хламидомонады имеет те же ключевые особенности, что и связанные с сенсорной трансдукцией каналы TRP у млекопитающих. Растительная клетка 27, 177–188. DOI: 10.1105 / tpc.114.131862
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Balagué, C., Lin, B., Alcon, C., Flottes, G., Malmström, S., Köhler, C., et al. (2003). HLM1, важный сигнальный компонент гиперчувствительного ответа, является членом семейства ионных каналов циклических нуклеотид-управляемых каналов. Растительная клетка 15, 365–379. DOI: 10.1105 / tpc.006999
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Болдуин, И.Т., Халичке Р., Пашольд А., фон Даль К. К. и Престон К. А. (2006). Изменчивая передача сигналов во взаимодействиях растений и растений: «говорящие деревья» в эпоху геномики. Наука 311, 812–815. DOI: 10.1126 / science.1118446
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Балуска, Ф., Манкузо, С., Фолькманн, Д., и Барлоу, П. (2004). Верхушки корня как командные центры растений: уникальный «мозговой» статус переходной зоны верхушки корня. Биология 59, 7–19.
Google Scholar
Балуска Ф., Фолькманн Д. и Барлоу П. В. (1996). Специализированные зоны развития в корнях: взгляд с клеточного уровня. Plant Physiol. 112, 3–4.
PubMed Аннотация | Google Scholar
Балушка Ф., Фолькманн Д. и Мензель Д. (2005). Синапсы растений: домены на основе актина для межклеточной коммуникации. Trends Plant Sci. 10, 106–111. DOI: 10.1016 / j.tplants.2005.01.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барбье-Брюгу, Х., De Angeli, A., Filleur, S., Frachisse, J.-M., Gambale, F., Thomine, S., et al. (2011). Анионные каналы / переносчики в растениях: от молекулярных основ до регуляторных сетей. Annu. Rev. Plant Biol. 62, 25–51. DOI: 10.1146 / annurev-arplant-042110-103741
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Becker, D., Geiger, D., Dunkel, M., Roller, A., Bertl, A., Latz, A., et al. (2004). AtTPK4, канал тандемной поры K + Arabidopsis , способный контролировать напряжение на мембране пыльцы в зависимости от pH и Ca 2+ . Proc. Natl. Акад. Sci. США . 101, 15621–15626. DOI: 10.1073 / pnas.0401502101
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беллоно, Н. В., Байрер, Дж. Р., Лейтч, Д. Б., Брайерли, С. М., Ингрэхэм, Х. А., Джулиус, Д., и др. (2017). Энтерохромаффинные клетки представляют собой хемосенсоры кишечника, которые соединяются с сенсорными нервными путями. Энтерохромаффинные клетки представляют собой хемосенсоры кишечника, которые соединяются с сенсорными нервными путями. Ячейка 170, 1–14. DOI: 10.1016 / j.ячейка.2017.05.034
CrossRef Полный текст | Google Scholar
Берриер, К., Пейроннет, Р., Беттон, Дж. М., Эфритихин, Г., Барбье-Брюгу, Х., Фракисс, Дж. М. и др. (2015). Канальные характеристики VDAC-3 из Arabidopsis thaliana . Biochem. Биофиз. Res. Коммуна . 459, 24–28. DOI: 10.1016 / j.bbrc.2015.02.034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бозе, Дж. К. (1907). Сравнительная электрофизиология, физико-физиологическое исследование. Лондон: Лонгманс.
Brault, M., Amiar, Z., Pennarun, A.-M., Monestiez, M., Zhang, Z., Cornel, D., et al. (2004). Деполяризация плазматической мембраны, индуцированная абсцизовой кислотой в суспензионных клетках Arabidopsis, включает уменьшение протонной перекачки в дополнение к активации анионных каналов, которые оба зависят от Ca2 +. Plant Physiol. 135, 231–243. DOI: 10.1104 / стр.103.039255
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бреннер, Э. Д., Стальберг, Р., Манкузо, С., Виванко, Дж., Балушка, Ф., и Ван Волкенбург, Э. (2006). Нейробиология растений: комплексный взгляд на передачу сигналов растений. Trends Plant Sci. 11, 413–419. DOI: 10.1016 / j.tplants.2006.06.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бюхнер П., Такахаши Х. и Хоуксфорд М. Дж. (2004). Переносчики сульфатов растений: координация поглощения, внутриклеточного и дальнего транспорта. J. Exp. Бот. 55, 1765–1773.DOI: 10.1093 / jxb / erh306
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берч-Смит, Т. М., и Замбрыски, П. К. (2012). Сдвиг парадигмы плазмодесм: регулирование извне по сравнению с внутренним. Annu. Rev. Plant Biol. 63, 239–260. DOI: 10.1146 / annurev-arplant-042811-105453
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каналес, Дж., Контрерас-Лопес, О., Альварес, Дж. М., и Гутьеррес, Р. А. (2017). Индукция нитратной плотности корневых волосков опосредуется факторами транскрипции TGA1 / TGA4 и CPC в Arabidopsis thaliana . Plant J. 92, 305–316. DOI: 10.1111 / TPJ.13656
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кандео А., Доккула Ф. Г., Валентини Г., Басси А. и Коста А. (2017). Световая флуоресцентная микроскопия позволяет количественно оценить колебания кальция в корневых волосках Arabidopsis thaliana . Physiol растительных клеток. 58, 1–12. DOI: 10.1093 / pcp / pcx045
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чин, К., Дефалко, Т.А., Мёдер, В., и Йошиока, К. (2013). Циклические нуклеотидно-управляемые ионные каналы Arabidopsis AtCNGC2 и AtCNGC4 работают в одном и том же сигнальном пути, регулируя защиту от патогенов и переход цветков. Plant Physiol. 163, 611–624. DOI: 10.1104 / стр.113.225680
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чой, В.-Г., Хиллири, Р., Суонсон, С.Дж., Ким, С.-Х., и Гилрой, С. (2016). Быстрая передача электрических и кальциевых сигналов на большие расстояния в растениях. Annu. Rev. Plant Biol. 67, 287–307. DOI: 10.1146 / annurev-arplant-043015-112130
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чой, В.-Г., Миллер, Г., Уоллес, И., Харпер, Дж., Миттлер, Р., и Гилрой, С. (2017). Организация быстрой передачи сигналов на большие расстояния в растениях с помощью Ca 2+ , ROS и электрических сигналов. Plant J. 90, 698–707. DOI: 10.1111 / tpj.13492
CrossRef Полный текст | Google Scholar
Клемменсен, К., Мюллер, Т. Д., Вудс, С. К., Берту, Х., Сили, Р. Дж., И Чёп, М. Х. (2017). Перекрестный разговор между кишечником и мозгом в метаболическом контроле. Cell 168, 758–774. DOI: 10.1016 / j.cell.2017.01.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Конн, А., Педмале, У. В., Чори, Дж., Стивенс, К. Ф., и Навлаха, С. (2017). Статистическое описание архитектуры побегов растений. Curr. Биол. 27, 2078.e3–2088.e3. DOI: 10.1016 / j.cub.2017.06.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дарвин, К.(1897). Насекомоядные растения . Эд Д. Эпплтон. Нью-Йорк, штат Нью-Йорк: Дж. Мюррей.
Дэвис, Э. (1987). Потенциалы действия как многофункциональные сигналы у растений: объединяющая гипотеза, объясняющая очевидно несопоставимые реакции на раны. Plant Cell Environ. 10, 623–631.
Google Scholar
Де Анджели, А., Моначелло, Д., Эфритикхин, Г., Фракисс, Дж. М., Томин, С., Гамбале, Ф., и др. (2006). Нитратный / протонный антипортер AtCLCa опосредует накопление нитратов в вакуолях растений. Природа . 442, 939–942. DOI: 10.1038 / nature05013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Де Анджели, А., Чжан, Дж., Мейер, С., и Мартиноя, Э. (2013). AtALMT9 представляет собой активируемый малатом вакуолярный хлоридный канал, необходимый для открытия устьиц у Arabidopsis. Нац Коммуна . 4, 1804. DOI: 10.1038 / ncomms2815
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дакетт, К. М., Опарка, К. Дж., Прайор, Д. А. М., Долан, Л.и Робертс К. (1994). Связывание красителя в корневом эпидермисе Arabidopsis постепенно снижается в процессе развития. Разработка 3255, 3247–3255.
Google Scholar
Данлоп Дж. И Градинер С. (1993). Поглощение фосфата, экструзия протонов и мембранные электропотенциалы фосфор-дефицитных Trifolium repens L. J. Exp. Бот. 44, 1801–1808. DOI: 10.1093 / jxb / 44.12.1801
CrossRef Полный текст | Google Scholar
Эдель, К.Х., Маршадье, Э., Браунли, К., Кудла, Дж., И Хетерингтон, А. М. (2017). Эволюция кальциевой передачи сигналов у растений. Curr. Биол. 27, R667 – R679. DOI: 10.1016 / j.cub.2017.05.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Инженер, К. Б., Хашимото-Сугимото, М., Неги, Дж., Исраэльссон-Нордстрём, М., Азулай-Шемер, Т., Раппель, В. Дж. И др. (2015). Зондирование CO2 и регуляция устьичной проводимости CO2: достижения и открытые вопросы. Trends Plant Sci. 21, 16–30. DOI: 10.1016 / j.tplants.2015.08.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фромм Дж. И Эшрих У. (1993). Электрические сигналы, исходящие от корней ивы ( Salix viminalis L.), изменяют транспирацию и фотосинтез. J. Plant Physiol. 141, 673–680. DOI: 10.1016 / S0176-1617 (11) 81573-7
CrossRef Полный текст | Google Scholar
Фромм, Дж., Хаджирезаи, М.-Р., Беккер, В.К., и Лотнер, С.(2013). Электрическая передача сигналов по флоэме и ее физиологические реакции в листе кукурузы. Фронт. Plant Sci. 4: 239. DOI: 10.3389 / fpls.2013.00239
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фромм, Дж., И Лотнер, С. (2012). «Электрофизиология растений: передача сигналов и реакции», в Электрофизиология растений , под ред. А.Г. Волкова (Берлин; Гейдельберг: Springer-Verlag), 1–377.
Фромм, Дж., Мейер, А. Дж., И Вайзензель, М.Х. (1997). На рост, мембранный потенциал и эндогенные ионные токи корней ивы ( Salix viminalis ) влияют абсцизовая кислота и спермин. Physiol. Растение. 99, 529–537. DOI: 10.1034 / j.1399-3054.1997.9.x
CrossRef Полный текст | Google Scholar
Габриэль Р. и Кессельмайер Дж. (1999). Апопластные концентрации растворенных органических кислот и минеральных питательных веществ в листьях некоторых грибных растений. Physiol растительных клеток. 40, 604–612.DOI: 10.1093 / oxfordjournals.pcp.a029583
CrossRef Полный текст | Google Scholar
Gajdanowicz, P., Michard, E., Sandmann, M., Rocha, M., Corrêa, L.G., Ramírez-Aguilar, S.J., et al. (2011). Градиенты калия (K +) служат мобильным источником энергии в тканях сосудов растений. Proc. Natl. Акад. Sci. США 108, 864–869. DOI: 10.1073 / pnas.1009777108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гальвани, Л. (1791). De viribus electricitatis in motu muscularis commentarius. Bononiensi Sci. Artium Inst. atque Acad. Комментарий. 7, 363–418. DOI: 10.1097 / 00000441-195402000-00023
CrossRef Полный текст
Geiger, D., Becker, D., Vosloh, D., Gambale, F., Palme, K., Rehers, M., et al. (2009). Гетеромерные каналы AtKC1 {средняя точка} AKT1 в корнях Arabidopsis способствуют росту в K + -лимитирующих условиях. J. Biol. Chem . 284, 21288–21295. DOI: 10.1074 / jbc.M109.017574
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стеклянная, А.Д., Шафф, Дж. Э. и Кочиан, Л. В. (1992). Исследования поглощения нитратов ячменем: IV. Электрофизиология. Plant Physiol. 99, 456–463.
PubMed Аннотация | Google Scholar
Гоберт А., Исаенков С., Фелькер К., Чемпински К. и Маатуис Ф. Дж. М. (2007). Двухпоровый канал , ген TPK1 кодирует вакуолярную проводимость K + и играет роль в гомеостазе K + . Proc. Natl. Акад. Sci. США . 104, 10726–10731.DOI: 10.1073 / pnas.0702595104
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Голдсуорси, А. (1983). Эволюция потенциалов действия растений. J. Theor. Биол. 103, 645–648.
граммов, Т. Э., Лотнер, С., Фелле, Х. Х., Матиссек, Р., и Фромм, Дж. (2009). Электрические сигналы, индуцированные нагреванием, влияют на цитоплазматический и апопластный pH, а также на фотосинтез во время прохождения через лист кукурузы. Plant Cell Environ. 32, 319–326.DOI: 10.1111 / j.1365-3040.2008.01922.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грубер Б. Д., Гиль Р. Ф. Х., Фридель С. и фон Вирен Н. (2013). Пластичность корневой системы арабидопсиса при недостатке питательных веществ. Plant Physiol. 163, 161–179. DOI: 10.1104 / стр.113.218453
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гунсе, Б., Пошенридер, К., Ранкл, С., Шредер, П., Родриго-Морено, А., и Барсело, Дж.(2016). Универсальная и легко настраиваемая система для электрофизиологии растений. Методы 3, 436–451. DOI: 10.1016 / j.mex.2016.05.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Го, Дж., Цзэн, В., Чен, К., Ли, К., Чен, Л., Ян, Ю. и др. (2016). Структура потенциалзависимого двухпорового канала TPC1 из Arabidopsis thaliana . Природа 531, 196–201. DOI: 10.1038 / природа16446
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хедрих, Р., Сальвадор-Рекатала, В., Дрейер, И. (2016). Электропроводка и междугородная заводская связь. Trends Plant Sci. 21, 376–387. DOI: 10.1016 / j.tplants.2016.01.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hegenauer, V., Fürst, U., Kaiser, B., Smoker, M., Zipfel, C., Felix, G., et al. (2016). Обнаружение паразита растений Cuscuta reflexa с помощью рецептора поверхности клетки томата. Наука 353, 478–481. DOI: 10.1126 / наука.aaf3919
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ходжкин, А. Л. (1937). Доказательства передачи электрического тока в нерве. J. Physiol. 90, 183–210.
Google Scholar
Ходжкин А. Л. и Хаксли А. Ф. (1952). Количественное описание мембранного тока и его применение к проводимости и возбуждению в нерве. J. Physiol. 117, 500–544.
PubMed Аннотация | Google Scholar
Надежда, А.Б. и Финдли Г. П. (1964). Потенциал действия в Чара . Physiol растительных клеток. 5, 377–380. DOI: 10.1093 / oxfordjournals.pcp.a079056
CrossRef Полный текст | Google Scholar
Хози, Э., Вавассер, А., Мулин, К., Дрейер, И., Гаймар, Ф., Порэ, Ф. и др. (2003). Канал Arabidopsis наружу K + канал GORK участвует в регуляции движений устьиц и транспирации растений. Proc. Natl. Акад. Sci.США . 100, 5549–5554. DOI: 10.1073 / pnas.0733970100
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джексон, М. Б. (2006). Молекулярная и клеточная биофизика . Кембридж: Издательство Кембриджского университета.
Google Scholar
Кандел, Э. Р. (2001). Молекулярная биология хранения памяти: диалог между геном и синапсами. Наука 294, 1030–1038. DOI: 10.1126 / science.1067020
CrossRef Полный текст | Google Scholar
Карбан, Р.(2015). Зондирование растений и коммуникация . Чикаго, Иллинойс: Издательство Чикагского университета.
Google Scholar
Киба Т., Кудо Т., Кодзима М. и Сакакибара Х. (2011). Гормональный контроль усвоения азота: роль ауксина, абсцизовой кислоты и цитокинина. J. Exp. Бот. 62, 1399–1409. DOI: 10.1093 / jxb / erq410
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ким, С. А., Квак, Дж. М., Джэ, С. К., Ван, М. Х. и Нам, Х.Г. (2001). Сверхэкспрессия гена AtGluR2 , кодирующего арабидопсис, гомолог глутаматных рецепторов млекопитающих, снижает утилизацию кальция и чувствительность к ионному стрессу у трансгенных растений. Физиология растительных клеток . 42, 74–84. DOI: 10.1093 / pcp / pce008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Киркби, Э. (2011). «Введение, определение и классификация питательных веществ», в Marschner’s Mineral Nutrition of High Plants, 3rd Edn ., изд П. Маршнер (Лондон: Elsevier), 3–5. DOI: 10.1016 / B978-0-12-384905-2.00001-7
CrossRef Полный текст | Google Scholar
Котула, Дж. У., Кернс, С. Дж., Шакет, Л. А., Сирадж, Л., Коллинз, Дж. Дж., Уэй, Дж. К. и др. (2014). Программируемые бактерии обнаруживают и записывают сигнал окружающей среды в кишечнике млекопитающих. Proc. Natl. Акад. Sci. США 111, 4838–4843. DOI: 10.1073 / pnas.1321321111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Курусу, Т., Хамада, Х., Кояно, Т., и Кучицу, К. (2012). Внутриклеточная локализация и физиологическая функция риса Ca 2+ -проницаемого канала OsTPC1. Завод Сигнал. Behav. 7, 1428–1430. DOI: 10.4161 / psb.22086
CrossRef Полный текст | Google Scholar
Лаанеметс, К., Брандт, Б., Ли, Дж., Мерило, Э., Ван, Й.-Ф., Кешвани, М. М. и др. (2013). Кальций-зависимая и независимая устьичная сигнальная сеть и компенсаторная обратная связь, управление открытием устьиц с помощью прайминга чувствительности к Ca2 +. Plant Physiol. 163, 504–513. DOI: 10.1104 / стр.113.220343
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ленг, К., Мерсье, Р. В., Хуа, Б.-Г., Фромм, Х., Берковиц, Г. А. (2002). Электрофизиологический анализ клонированных циклических нуклеотид-управляемых ионных каналов. Физиология растений . 128, 400–410. DOI: 10.1104 / pp.010832
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ленг, К., Мерсье, Р. В., Яо, В., и Берковиц, Г.А. (1999). Клонирование и первая функциональная характеристика катионного канала, управляемого циклическими нуклеотидами. Физиология растений . 121, 753–761.
PubMed Аннотация | Google Scholar
Li, Z. Y., Xu, Z. S., He, G. Y., Yang, G. X., Chen, M., Li, L. C., et al. (2013). Напряжение-зависимый анионный канал 1 (AtVDAC1) отрицательно регулирует реакцию растений на холод во время прорастания и развития проростков Arabidopsis и взаимодействует с сенсором кальцияCBL1. Внутр. J. Mol.Sci . 14, 701–713. DOI: 10.3390 / ijms14010701
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю К. Х., Ню Й., Кониси М., Ву Й., Ду Х., Сун Чунг Х. и др. (2017). Открытие передачи сигналов нитрат-CPK-NLP в центральных сетях роста питательных веществ. Природа 545, 311–316. DOI: 10.1038 / nature22077
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лопес-Миллан, А. Ф., Моралес, Ф., Абадия, А., и Абадия, Дж. (2001).Связанные с дефицитом железа изменения в составе апопластной жидкости листьев полевых деревьев груши ( Pyrus communis L.). J. Exp. Бот. 52, 1489–1498. DOI: 10.1093 / jexbot / 52.360.1489
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ма, В., Смигель, А., Уокер, Р. К., Мёдер, В., Йошиока, К., и Берковиц, Г. А. (2010). Передача сигналов о старении листьев: Ca2 + -проводящий канал 2, управляемый циклическими нуклеотидами Arabidopsis, действует через оксид азота, подавляя программирование старения. Физиология растений . 154, 733–743. DOI: 10.1104 / стр.110.161356
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Malcheska, F., Ahmad, A., Batool, S., Müller, H.M., Ludwig-Müller, J., Kreuzwieser, J., et al. (2017). Усиленный засухой сульфат ксилемного сока закрывает устьица, влияя на ALMT12 и синтез АБК замыкающих клеток. Plant Physiol. 174, 798–814. DOI: 10.1104 / стр. 16.01784
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартинац, Б.(2008). Зондирование с помощью ионных каналов . изд Б. Мартинак. Берлин; Гейдельберг: Springer.
Google Scholar
Маси, Э., Цисак, М., Компарини, Д., Монетти, Э., Пандольфи, К., Аццарелло, Э. и др. (2015). Электрическая сеть верхушки корня кукурузы зависит от силы тяжести. Sci. Отчет 5: 7730. DOI: 10.1038 / srep07730
PubMed Аннотация | CrossRef Полный текст | Google Scholar
МакКлюр, П. Р., Кочиан, Л. В., Спансвик, Р. М., и Шафф, Дж.Э. (1990). Доказательства совместного переноса нитратов и протонов в корнях кукурузы: I. Влияние нитратов на мембранный потенциал. Plant Physiol. 93, 281–289.
PubMed Аннотация | Google Scholar
Мехарг, А.А., Блатт, М.Р. (1995). Транспорт NO3 через плазматическую мембрану корневых волосков Arabidopsis thaliana : кинетический контроль с помощью pH и мембранного напряжения. J. Membr. Биол. 145, 49–66. DOI: 10.1007 / BF00233306
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Купец, С.С., Прочник, С. Е., Валлон, О., Харрис, Э. Х., Карпович, С. Дж., Витман, Г. Б. и др. (2007). Геном хламидомонады показывает эволюцию ключевых функций животных и растений. Наука 318, 245–250. DOI: 10.1126 / science.1143609
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мейер, С., Мумм, П., Имес, Д., Эндлер, А., Ведер, Б., Аль-Рашейд, К. А. и др. (2010). AtALMT12 представляет собой анионный канал R-типа, необходимый для движения устьиц в замыкающих клетках арабидопсиса. Завод J . 63, 1054–1062. DOI: 10.1111 / j.1365-313X.2010.04302.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мулин К., Вери А.-А., Гаймар Ф., Бушерез Дж., Пилот Г., Девич М. и др. (2002). Развитие пыльцевых трубок и конкурентоспособность ухудшаются из-за нарушения канала Shaker K + в Arabidopsis . Genes Dev . 16, 339–350. DOI: 10.1101 / gad.213902
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мусави, С.А., Шовен, А., Паско, Ф., Келленбергер, С., и Фармер, Э. Э. (2013). Гены, подобные ГЛУТАМАТНЫМ РЕЦЕПТОРАМ, опосредуют передачу сигналов от раны от листа к листу. Природа 500, 422–426. DOI: 10.1038 / природа12478
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мучхал, США, и Рагхотама, К.Г. (1999). Транскрипционная регуляция переносчиков фосфатов растений. Proc. Natl. Акад. Sci. США 96, 5868–5872.
PubMed Аннотация | Google Scholar
Наврат, К., Шрайбер, Л., Франке, Р. Б., Гелднер, Н., Рейна-Пинто, Дж. Дж., И Кунст, Л. (2013). Барьеры апопластной диффузии у арабидопсиса. Араб. Б. 11: е0167. DOI: 10.1199 / tab.0167
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Окубо Ю., Танака М., Табата Р., Огава-Охниши М. и Мацубаяси Ю. (2017). Мобильные полипептиды от побегов к корням участвуют в системной регуляции усвоения азота. Nat. Растения 3, 17029. doi: 10.1038 / nplants.2017.29
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Peiter, E., Maathuis, F. J. M., Mills, L. N., Knight, H., Pelloux, J., Hetherington, A.M, et al. (2005). Вакуолярный Ca 2+ -активированный канал TPC1 регулирует прорастание и движение устьиц. Природа 434, 404–408. DOI: 10.1038 / nature03381
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перет Б., Клеман М., Нуссом Л. и Деснос Т. (2011).Адаптация развития корней к фосфатному голоданию: лучше перестраховаться. Trends Plant Sci. 16, 442–450. DOI: 10.1016 / j.tplants.2011.05.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Pietruszka, M., Stolarek, J. A. N., and Pazurkiewicz-kocot, K. (1997). Эволюция во времени потенциала действия в клетках растений. J. Biol. Phys. 23, 219–232.
PubMed Аннотация | Google Scholar
Приндл, А., Лю, Дж., Асалли, М., Ли, С., Гарсия-Ойалво, Дж., Сюэль, Г. М. и др. (2015). Ионные каналы обеспечивают электрическую связь в бактериальных сообществах. Природа 527, 59–63. DOI: 10.1038 / природа15709
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Риверас, Э., Альварес, Дж. М., Видаль, Э. А., Осес, К., Вега, А., и Гутьеррес, Р. А. (2015). Ион кальция является вторым посредником в сигнальном пути нитратов Arabidopsis. Plant Physiol. 169, 1397–1404. DOI: 10.1104 / стр. 15.00961
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rocchetti, A., Sharma, T., Wulfetange, C., Scholz-Starke, J., Grippa, A., Carpaneto, A., et al. (2012). Предполагаемая субъединица K (+) канала AtKCO3 образует стабильные димеры у Arabidopsis. Фасадный завод Sci . 3: 251. DOI: 10.3389 / fpls.2012.00251
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ронзье, Э., Корратже-Фэйли, К., Санчес, Ф., Прадо, К., Бриер, К., Леонхардт Н. и др. (2014). CPK13, неканоническая Са2 + -зависимая протеинкиназа, специфически ингибирует K + каналы KAT2 и KAT1shaker и уменьшает открытие устьиц. Физиология растений . 166, 314–326. DOI: 10.1104 / стр.114.240226
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рубио, В., Линьярес, Ф., Солано, Р., Мартин, А.С., Иглесиас, Дж., Лейва, А. и др. (2001). Консервативный фактор транскрипции MYB, участвующий в передаче сигналов фосфатного голодания как у сосудистых растений, так и у одноклеточных водорослей. Genes Dev. 15, 2122–2133. DOI: 10.1101 / gad.204401
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сакано, К. (1990). Протонно-фосфатная стехиометрия поглощения неорганического фосфата культивируемыми клетками Catharanthus roseus (L.) G. Don. Plant Physiol. 93, 479–83.
PubMed Аннотация | Google Scholar
Salvador-Recatalà, V. (2016a). Новые роли генов, подобных рецепторам глутамата 3.3, 3.5 и 3.6, как переключателей включения / выключения системных электрических сигналов, вызванных раной. Завод Сигнал. Behav. 11: e1161879. DOI: 10.1080 / 155.2016.1161879
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Salvador-Recatalà, V. (2016b). Калиевый канал AKT2 обеспечивает деполяризацию, вызванную NaCl, в корне Arabidopsis thaliana . Завод Сигнал. Behav. 11: e1165381. DOI: 10.1080 / 155.2016.1165381
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сальвадор-Рекатала, V., Tjallingii, W. F., and Farmer, E. E. (2014). В реальном времени, in vivo внутриклеточных записей вызванных гусеницами волн деполяризации в элементах сита с использованием тлейных электродов. New Phytol. 203, 674–684. DOI: 10.1111 / Nph.12807
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сандерсон, Дж. Б. (1872). Обратите внимание на электрические явления, которые сопровождают раздражение листа Dionaea muscipula . Proc. R Soc. Лондон. 21, 495–496.DOI: 10.1098 / RSPL.1872.0092
CrossRef Полный текст
Саттельмахер Б. (2001). Апопласт и его значение для минерального питания растений. New Phytol. 149, 167–192. DOI: 10.1046 / j.1469-8137.2001.00034.x
CrossRef Полный текст | Google Scholar
Саттельмахер Б. и Хорст В. Дж. (2007). Апопласт высших растений: отделение хранения, транспорта и реакций: значение апопласта для минерального питания высших растений. Дордрехт: Спрингер.
Google Scholar
Шредер Дж. И., Хедрих Р. и Фернандес Дж. М. (1984). Калий-селективные одиночные каналы в протопластах замыкающих клеток Vicia faba . Природа 312, 361–362. DOI: 10.1038 / 312361a0
CrossRef Полный текст | Google Scholar
Спансвик, Р. М., и Костертон, Дж. У. Ф. (1967). Plasmodesmata in Nitella translucens : структура и электрическое сопротивление. J. Cell Sci. 2, 451–464.
PubMed Аннотация | Google Scholar
Широки А., Ивашикина Н., Дитрих П., Роэльфсема М. Р. Г., Аче П., Рейнтанц Б. и др. (2001). KAT1 не важен для открытия устьиц. Proc. Natl. Акад. Sci. США . 98, 2917–2921. DOI: 10.1073 / pnas.051616698
CrossRef Полный текст | Google Scholar
Татеда, К., Ватанабе, К., Кусано, Т., и Такахаши, Ю. (2011). Молекулярная и генетическая характеристика семейства генов, кодирующих потенциал-зависимый анионный канал у Arabidopsis. J. Exp. Бот . 62, 4773–4785. DOI: 10.1093 / jxb / err113
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тилснер, Дж., Николас, В., Росадо, А., и Байер, Э. М. (2016). Сохранение герметичности: места контакта плазмодесмальной мембраны и контроль межклеточной связи у растений. Annu. Rev. Plant Biol. 67, 1–28. DOI: 10.1146 / annurev-arplant-043015-111840
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Требач, К.и Завадски Т. (1985). Световые потенциалы действия в печеночнике Conocephalum conicum . Physiol. Растение. 64, 482–486. DOI: 10.1111 / j.1399-3054.1985.tb08526.x
CrossRef Полный текст | Google Scholar
Ullrich-Eberius, C. I., Novacky, A., Fischer, E., and Lüttge, U. (1981). Связь между энергозависимым поглощением фосфата и электрическим мембранным потенциалом у lemna gibba G1. Plant Physiol. 67, 797–801.
PubMed Аннотация | Google Scholar
Видаль, Э.A., Araus, V., Lu, C., Parry, G., Green, P.J., Coruzzi, G.M., et al. (2010). Нитрат-чувствительный регуляторный модуль miR393 / AFB3 контролирует архитектуру корневой системы у Arabidopsis thaliana . Proc. Natl. Акад. Sci. США 107, 4477–4482. DOI: 10.1073 / pnas.01107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фёлькер, К., Шмидт, Д., Мюллер-Робер, Б., и Чемпински, К. (2006). Члены семейства Arabidopsis AtTPK / KCO образуют гомомерные вакуолярные каналы in planta . Завод J . 48, 296–306. DOI: 10.1111 / j.1365-313X.2006.02868.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Waese, J., Fan, J., Pasha, A., Yu, H., Fucile, G., Shi, R., et al. (2017). ePlant: визуализация и изучение нескольких уровней данных для генерации гипотез в биологии растений. Растительная клетка 29, 1806–1821. DOI: 10.1105 / tpc.17.00073
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, Р. и Кроуфорд, Н.М. (1996). Генетическая идентификация гена, участвующего в конститутивном высокоаффинном транспорте нитратов у высших растений. Proc. Natl. Акад. Sci. США 93, 9297–9301.
PubMed Аннотация | Google Scholar
Ван Р., Лю Д. и Кроуфорд Н. М. (1998). Белок CHL1 Arabidopsis играет важную роль в захвате высокоаффинных нитратов. Proc. Natl. Акад. Sci. U.S. A. 95, 15134–15139.
PubMed Аннотация | Google Scholar
Ван, Р., Окамото, М., Син, X., и Кроуфорд, Н. М. (2003). Микроматричный анализ нитратного ответа в корнях и побегах арабидопсиса выявил более 1000 быстро реагирующих генов и новые связи с метаболизмом глюкозы, трегалозо-6-фосфата, железа и сульфата. Plant Physiol. 132, 556–567. DOI: 10.1104 / стр.103.021253
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уорд, Дж. М., Мазер, П., Шредер, Дж. И. (2009). Ионные каналы растений: семейства генов, физиология и анализ функциональной геномики. Annu. Rev. Physiol. 71, 59–82. DOI: 10.1146 / annurev.physiol.010908.163204
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Xu, J., Li, H.-D., Chen, L.-Q., Wang, Y., Liu, L.-L., He, L., et al. (2006). Протеинкиназа, взаимодействуя с двумя белками, подобными кальциневрину B, регулирует транспортер K + AKT1 в Arabidopsis . Cell 125, 1347–1360. DOI: 10.1016 / j.cell.2006.06.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ян, Дж., He, H., Tong, S., Zhang, W., Wang, J., Li, X., et al. (2009). Напряжение-зависимый анионный канал 2 Arabidopsis thaliana (AtVDAC2) участвует в АБК-опосредованном раннем развитии проростков. Внутр. J. Mol. Sci . 10, 2476–2486. DOI: 10.3390 / ijms10062476
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yang, W., Nagasawa, K., Münch, C., Xu, Y., Satterstrom, K., Jeong, S., et al. (2016). Митохондриальная сиртуиновая сеть обнаруживает динамическое SIRT3-зависимое деацетилирование в ответ на деполяризацию мембраны. Cell 0, 14447–14452. DOI: 10.1016 / j.cell.2016.10.016
CrossRef Полный текст | Google Scholar
Ян, X. Y., Chen, Z. W., Xu, T., Qu, Z., Pan, X. D., Qin, X. H., et al. (2011). Кинезин KP1 арабидопсиса специфически взаимодействует с VDAC3, митохондриальным белком, и регулирует дыхание во время прорастания семян при низкой температуре. Растительная клетка 23, 1093–1106. DOI: 10.1105 / tpc.110.082420
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Завадски, Т., Дэвис, Э., Бзюбинска, Х., Хткбач, К., и Характеристики, К. (1991). Характеристика потенциалов действия у Helianthus annuus . Physiol. Растение. 83, 601–604.
Google Scholar
Зебело, С. А., Мацуи, К., Одзава, Р., и Маффеи, М. Е. (2012). Деполяризация потенциала плазматической мембраны и цитозольный поток кальция являются ранними событиями, участвующими в коммуникации растений томатов ( Solanum lycopersicon ). Plant Sci. 196, 93–100.DOI: 10.1016 / j.plantsci.2012.08.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhang, A., Ren, H.-M., Tan, Y.-Q., Qi, G.-N., Yao, F.-Y., Wu, G.-L., et al. (2016). Анионные каналы S-типа SLAC1 и SLAh4 функционируют как важные негативные регуляторы внутренних каналов K + и устьичных отверстий у Arabidopsis . Растительная клетка 28, 949–965. DOI: 10.1105 / tpc.16.01050
CrossRef Полный текст | Google Scholar
Чжан, Дж., Мартиноя, Э., и Де Анджели, А. (2014). Цитозольные нуклеотиды блокируют и регулируют канал вакуолярного аниона Arabidopsis AtALMT9. J. Biol. Chem. 289, 25581–25589. DOI: 10.1074 / jbc.M114.576108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжэн, X., Хе, К., Клейст, Т., Чен, Ф., и Луан, С. (2015). Анионный канал SLAh4 участвует в нитратзависимом снижении токсичности аммония у Arabidopsis. Среда растительных клеток . 38, 474–486.DOI: 10.1111 / pce.12389
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжоу В., Брокмеллер Т., Линг З., Омдал А., Болдуин И. Т. и Сюй С. (2016). Эволюция ранней защиты, индуцированной травоядными животными, была сформирована у Nicotiana благодаря дупликации по всему геному. eLife 5: e19531. DOI: 10.7554 / eLife.19531
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Циммерманн, М. Р., Майшак, Х., Митхёфер, А., Боланд, В., и Фелле, Х.Х. (2009). Системные потенциалы, новый электрический сигнал апопласта на большие расстояния у растений, вызванный ранением. Plant Physiol. 149, 1593–1600. DOI: 10.1104 / стр.108.133884
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Биоэлектрические сигналы как новые биометрические параметры: проблемы и проблемы
В этой статье представлена эффективность биоэлектрических сигналов, таких как электрокардиограмма (ЭКГ) и электроэнцефалограмма (ЭЭГ), для биометрических приложений.Исследования показывают, что импульсы сердечного ритма и электрической активности мозга регистрируются на ЭКГ и ЭЭГ соответственно; обладают уникальными особенностями среди людей, поэтому их можно предложить использовать в качестве биометрических данных для проверки личности. Благоприятные характеристики для использования сигналов ЭКГ или ЭЭГ в качестве биометрических включают универсальность, измеримость, уникальность и надежность. Кроме того, им присуща жизненная сила, которая означает жизненные знаки, обеспечивающие надежную защиту от атак подделки.В отличие от традиционной биометрии, ЭКГ или ЭЭГ являются строго конфиденциальными и безопасными для человека, которые трудно подделать. Мы представляем обзор методов, используемых для ЭКГ и ЭЭГ в качестве биометрии для индивидуальной аутентификации, и сравниваем их эффективность на наборах данных и условиях тестирования, которые они использовали. Мы проиллюстрируем проблемы, связанные с использованием ЭКГ или ЭЭГ в качестве биометрических данных, в первую очередь из-за наличия значительных вариаций сбора данных и отсутствия стандартизации характеристик сигнала.Еще одна проблема, которую предстоит решить, — это определить масштабную производительность, индивидуальность ЭКГ или ЭЭГ.
1. Введение
1.1. Биоэлектрические сигналы
Биоэлектрические сигналы — это электрические сигналы очень низкой амплитуды и низкой частоты, которые могут быть измерены от биологических существ, например людей. Биоэлектрические сигналы генерируются сложной системой саморегулирования, и их можно измерить по изменениям электрического потенциала в клетке или органе.К интересующим нас биоэлектрическим сигналам относятся, в частности, электрокардиограмма (ЭКГ) и электроэнцефалограмма (ЭЭГ). ЭКГ измеряет электрическое проявление ионного потенциала сердца, в то время как ЭЭГ измеряет электрическую активность, вызванную вдоль скальпа мозга. ЭКГ и ЭЭГ записываются с использованием стандартного оборудования неинвазивным способом. Исследователи из разных дисциплин проявили больший интерес к анализу ЭКГ и ЭЭГ, чтобы понять, какие высокоуровневые характеристики создает человек.Однако междисциплинарный анализ биоэлектрических сигналов не только помогает в оценке состояния здоровья людей, но также предполагает, что биоэлектрические сигналы могут использоваться в качестве кандидата биометрических данных для проверки личности.
1.2. Биометрические характеристики биоэлектрических сигналов
Биометрия призвана облегчить систему управления идентификацией для достижения более высокого уровня точности, в то время как она использует анатомические и поведенческие характеристики людей, которые являются уникальными и измеримыми.Анатомические части тела и методы передачи сигналов включают лицо, отпечатки пальцев, руки, глаза, уши, вены и голос, в то время как поведенческие характеристики включают рукописную подпись, нажатие клавиш и походку [1]. Ограничения использования обычных биометрических данных включают то, что они являются уникальными идентификаторами, но они не являются конфиденциальными и не являются секретными для отдельного человека. Например, люди оставляют отпечатки пальцев на всем, к чему прикасаются, узоры радужной оболочки можно наблюдать везде, куда бы они ни посмотрели, лица видны, а голоса записываются.Наличие биометрических отпечатков публично, предлагая злоумышленникам поднять эти отпечатки и скопировать их как настоящие, таким образом подделывает систему. Использование биоэлектрических сигналов в качестве биометрии дает системе управления идентификацией ряд преимуществ. Помимо своей уникальности, биоэлектрические сигналы конфиденциальны и безопасны для человека. Их трудно подражать и копировать. Таким образом, личность человека вряд ли будет подделана, что позволит сохранить секретность и конфиденциальность пользователей.
Биоэлектрические сигналы, особенно ЭКГ и ЭЭГ, являются новыми биометрическими идентичностями [2–17]. В отличие от анатомических биометрических идентификаторов, которые имеют двумерное представление данных, ЭКГ или ЭЭГ — это физиологически низкочастотные сигналы, которые имеют одномерное представление данных. Использование сигналов ЭКГ или ЭЭГ в качестве биометрических может предложить следующие характеристики: универсальность, измеримость, уникальность и надежность. Универсальность означает, что каждый (живой) человек должен обладать сигналами ЭКГ или ЭЭГ.Измеримость означает, что сигналы ЭКГ или ЭЭГ могут быть записаны с помощью электродов, размещенных на поверхности тела рядом с конкретным органом (например, грудная клетка, руки и ноги для ЭКГ и вдоль кожи головы для ЭЭГ).
Биологическая информация человека, генетически регулируемая белками дезоксирибонуклеиновой кислоты (ДНК) или рибонуклеиновой кислоты (РНК) [18, 19]. В конце концов, белки ответственны за существование уникальности в определенных частях тела [1]. Точно так же такие органы, как сердце и мозг, состоят из белковых тканей, называемых миокардом и глиальными клетками соответственно.Следовательно, электрические сигналы, исходящие от этих органов, у разных людей уникальны [20]. И последнее, но не менее важное: воспроизведение или воспроизведение сигналов ЭКГ или ЭЭГ очень сложно до тех пор, пока один и тот же человек не будет вызван на повторную регистрацию. Таким образом, предлагаемые методы, использующие ЭКГ или ЭЭГ в качестве биометрических данных, достаточно неуязвимы для атак подделки. Следовательно, факт использования сигналов ЭКГ или ЭЭГ в качестве биометрических дает уверенность в том, что биометрические данные поступают от законного человека, который действительно присутствует во время регистрации.Это необходимое условие для безупречной работы практичной биометрической системы.
1.3. Краткое содержание статьи
В этой статье мы представляем обзор эффективности ЭКГ и сигналов ЭКГ для биометрических приложений. Критически анализируются различные методы, которые показывают перспективность ЭКГ или ЭЭГ в качестве кандидата на биометрические данные. Кроме того, обсуждаются вопросы и проблемы использования ЭКГ или ЭЭГ в качестве биометрических. Статья построена следующим образом. В разделе 2 описывается потенциал сигнала ЭКГ как биометрического параметра для индивидуальной аутентификации.Дан обзор сигнала ЭКГ и факторов, поддерживающих его использование в качестве биометрических данных. Представлен обзор существующих методов, которые исследуют возможность использования сигнала ЭКГ как биометрического, и нового метода, подтверждающего перспективность ЭКГ как биометрического. Выделены проблемы и проблемы при разработке биометрической системы с поддержкой ЭКГ, которая выполняет задачу аутентификации в широком диапазоне условий. В разделе 3 исследуется потенциал ЭЭГ с биометрической точки зрения, где представлен обзор ЭЭГ и основные особенности, позволяющие использовать ее в качестве биометрических данных.Дан обзор существующих методов, демонстрирующих лабораторные результаты в поддержку ЭЭГ как кандидата биометрических данных. Обобщены проблемы с точки зрения операционной жизнеспособности ЭЭГ как биометрического метода. Раздел 4 определяет некоторые из перспективных направлений будущих исследований для успешного использования сигналов ЭКГ или ЭЭГ для биометрических приложений.
2. Биометрическая ЭКГ
2.1. Преамбула ЭКГ
ЭКГ — это неинвазивный инструмент, используемый для регистрации электрических проявлений сократительной и расслабляющей активности сердца.Нобелевский лауреат Виллем Эйнтховен был первым, кто записал ЭКГ в 1903 году [21]. Его можно зарегистрировать с помощью поверхностных электродов, размещенных на конечностях и груди. Устройства ЭКГ используют различное количество электродов от 3 до 12 для регистрации сигнала, в то время как система, использующая большее количество электродов от 12 до 120, также доступна [22]. Каждый нормальный цикл сигнала ЭКГ содержит зубцы P, QRS и T (например, см. Рисунок 1). Зубец P представляет собой сокращение предсердной мышцы и имеет продолжительность 60–100 миллисекунд (мс).Он имеет низкоамплитудную морфологию 0,1–0,25 милливольт (мВ) и обычно обнаруживается в начале сердцебиения. Комплекс QRS является результатом деполяризации беспорядочных желудочков. Это резкая двухфазная или трехфазная волна длительностью 80–120 мс, демонстрирующая значительное отклонение амплитуды, которое варьируется от человека к человеку. Время, необходимое для распространения ионного потенциала от синусового узла через мышцу предсердия и попадания в желудочки, составляет 120–200 мс и известно как интервал PR. Желудочки имеют относительно большую длительность ионного потенциала 300–420 мс, известную как интервал QT.Плато ионного потенциала составляет 80–120 мс после QRS и известно как сегмент ST. Возврат желудочковой мышцы в его ионное состояние покоя вызывает зубец Т, который имеет амплитуду 0,1–0,5 мВ и длительность 120–180 мс. Продолжительность от покоя желудочков до начала следующего цикла сокращения предсердий известна как сегмент TP, который представляет собой длинную часть плато с незначительным подъемом.
Сигналы ЭКГ, полученные от разных людей, имеют неоднородные характеристики.Разнородность была отмечена в исследованиях, проведенных для диагностики аритмий, присутствующих в функции сердца [23]. Отличительные особенности сигналов ЭКГ обычно возникают из-за изменения ионного потенциала, времени распространения ионного потенциала из разных частей сердечной мышцы, уровней электролитов в плазме (например, калия, кальция и магния и т. Д.) И ритмических различий. Эти различия отражаются в различных формах, таких как изменение морфологии, разница в амплитудах и изменение временных интервалов доминирующих реперных точек сердечных сокращений.Различия в структуре сердца, такой как геометрия грудной клетки, положение, размер и физическое состояние, у разных людей также проявляют уникальные характеристики в их ритме сердцебиения.
2.2. Поддерживающие факторы
В литературе были предложены различные методы в поддержку использования сигнала ЭКГ в качестве кандидата биометрических данных для проверки личности [2–10]. В отличие от других традиционных биометрических данных (например, лица и отпечатков пальцев), сигнал ЭКГ обеспечивает обратную связь жизнеспособности в реальном времени.Биометрическая идентичность с активностью означает признаки жизни. Это гарантирует, что биометрический образец будет взят у живого человека для аутентификации. Таким образом, он предлагает веский аргумент против атак с подделкой на биометрическую систему безопасности. Кроме того, ЭКГ часто используется врачами для диагностики сердечных и других заболеваний. Благодаря развертыванию биометрической системы с поддержкой ЭКГ личность людей может быть проверена онлайн во время мониторинга ЭКГ или офлайн с помощью их медицинских записей.Его можно дополнительно дополнить существующими современными системами, такими как мобильный мониторинг здоровья или сенсорная сеть в области тела. Этот способ проверки личности гораздо более полезен для защиты личности и защиты сердечно-сосудистых заболеваний, особенно для пациентов с сердечно-сосудистыми заболеваниями [24]. Эта частная медицинская информация строго защищена в соответствии со стандартизацией регулирования конфиденциальности Закона о переносимости и подотчетности медицинского страхования (HIPAA) 1996 г. [25].
Хотя методы, использующие ЭКГ в качестве биометрических данных, могут не обеспечивать адекватной точности [26, 27], информация ЭКГ может помочь в повышении безопасности системы с другими биометрическими данными в мультибиометрической структуре.Singh et al. [27] предложили мультибиометрическую систему, которая получается путем объединения сигнала ЭКГ с биометрическими данными лица и отпечатка пальца для надежной аутентификации человека. Производительность предложенной системы была оптимальной, и было обнаружено, что равный коэффициент ошибок (EER) объединенной системы составляет 0,2% по сравнению с отдельной системой ЭКГ, которая имеет ERR 10,8%, в то время как EER лицо и системы отпечатков пальцев — 4,5% и 2,1% соответственно. Они пришли к выводу, что объединенная система может работать так, как это воспринимается пользователем, так что данные ЭКГ человека могут быть получены одновременно с другими биометрическими данными, то есть отпечатками пальцев и получением изображения лица.
Так как белки ДНК и РНК в конечном итоге ответственны за существование уникальности в идентичности лиц, отпечатков пальцев и других следов тела. Поэтому обязательно отметить, что биометрические параметры, такие как геометрия лица, текстура пальцев и электрическое проявление, вызванное сердцем, ортогональны друг другу. В конце концов, существует корреляция между функцией сердца (ЭКГ) и характеристиками других биометрических данных. При объединении ЭКГ с другими биометрическими данными, если ЭКГ может сузить область поиска до половины (50%), тогда шансы на успех достичь нужного человека из оставшейся популяции будут удвоены (с использованием других биометрических данных).Таким образом, менее эффективная биометрическая информация может значительно оптимизировать пространство поиска, чтобы дополнить другие биометрические параметры для удовлетворения требований приложений строгой безопасности.
Помимо генетических причин, ЭКГ как биометрический показатель подтверждается тем фактом, что физиологические и геометрические различия между людьми обуславливают определенную степень уникальности их сердцебиения [28]. Хотя поддерживающие факторы, обсужденные до сих пор, предлагают выбор в пользу выбора ЭКГ в качестве биометрических данных, основанных на лабораторной демонстрации, они очень многообещающие.Следовательно, пришло время разработать биометрическую систему с поддержкой ЭКГ, которую можно использовать для конкретного приложения мониторинга пациента, и после этого возможность применения системы может быть проверена для расширенной области приложений.
2.3. Связанные работы
Анализ ЭКГ и интерпретация моделей ЭКГ как инструмент клинической диагностики — активные области исследований. Используя преимущества исследования, в недавнем прошлом было проведено несколько исследований, изучающих возможность использования ЭКГ в качестве нового кандидата биометрических данных для проверки личности.
Israel et al. [2] показали, что ЭКГ индивидуума демонстрирует уникальный паттерн. Они выполнили обработку ЭКГ для проверки качества, и предложены количественные показатели для классификации сердечных сокращений среди людей. Они извлекли из каждого биения сердца стабильные черты, характеризующие уникальность человека. Эксперимент был протестирован на 29 людях от мужчин и женщин в возрасте 22 и 48 лет, где собирались данные о различных состояниях тревоги и на разных сессиях.Из каждого сердечного сокращения было извлечено в общей сложности 15 внутриудаленных характеристик, основанных на физиологии сердца, и классификация выполняется с использованием линейного дискриминантного анализа. Они показали, что извлеченные признаки не зависят от положения электродов (например, вокруг груди и шеи), инвариантны к состоянию тревоги и уникальны для каждого человека. Проблемы, например, масштабируемость функций ЭКГ для аутентификации большой совокупности и неизменность функций в течение более длительных периодов времени, исследуются на большом наборе данных.
Анализ собственного пространства записи ЭКГ для идентификации человека был представлен в [3]. Подобно тому, как традиционные биометрические методы (например, лица, отпечатков пальцев и радужной оболочки глаза) использовали метод анализа основных компонентов (PCA) для использования образцов изображений, декомпозиция сердцебиения по собственному алгоритму дала более надежный подход к идентификации человека. Методология проверки личности с использованием ЭКГ, которая определяет минимальное количество сердечных сокращений, необходимых для аутентификации человека, обсуждалась в [4].Подход основан на статистической теории последовательных процедур, которые извлекают характеристики сердцебиения для вычисления статистики теста. Решение о том, что полученная ЭКГ соответствует сохраненным учетным данным заявленной личности, было принято с использованием непрерывной выборки сердечных сокращений.
Одно из самых ранних исследований, продемонстрировавших возможное использование сигнала ЭКГ для биометрических приложений, было проведено Biel et al. [5]. Они провели биометрический эксперимент на группе из 20 человек, включая мужчин и женщин, каждый из которых относится к возрасту 20–55 лет.Для каждого субъекта данные ЭКГ были собраны за период времени в шесть недель, и в общей сложности было извлечено 30 характеристик из каждого сердцебиения. Все эти извлеченные признаки обычно используются при клинической диагностике сердечной аритмии. Чтобы уменьшить количество признаков, признаки с относительно высокой корреляцией с другими признаками отбрасываются, и, наконец, для классификации было выбрано 12 признаков. Для классификации использовался метод, основанный на многомерном анализе; тем не менее, для интерпретации сходства и различий сердечных сокращений среди людей использовался график оценки основных компонентов (PCA).
Возможность использования ЭКГ в качестве новой биометрии для проверки личности была показана Shen et al. [6]. Они провели эксперимент по внешнему виду и характеристикам сердцебиения во временной области. Однако большинство функций было извлечено из комплекса QRS, который стабилен при изменении частоты сердечных сокращений. Одна особенность — интервал QT, который меняется с изменением частоты сердечных сокращений, нормализовался. Подходы нейронной сети на основе сопоставления шаблонов и принятия решений использовались для количественной оценки скорости проверки личности, которая, как сообщается, составляет 95% и 80% соответственно.После объединения подходов к классификации результат проверки личности оказался 100% от группы из 20 человек.
Wang et al. [7] представили двухэтапную структуру определения реперных точек, которая включает в себя аналитические и основанные на внешнем виде функции из сердцебиения. Аналитические функции собирают локальную информацию, которая объединяет временные и амплитудные характеристики, а функции, основанные на внешнем виде, фиксируют целостные модели сердцебиения. Чтобы лучше использовать дополнительные характеристики аналитических функций и функций внешнего вида, была представлена иерархическая схема интеграции данных.Метод, используемый для выделения признаков, основан на комбинации автокорреляции (AC) и дискретного косинусного преобразования (DCT). Метод AC / DCT работает с оконными сегментами ЭКГ и поэтому не требует синхронизации импульсов. Эффективность распознавания, достигнутая методом AC / DCT, составляет 94,47% и 97,8% соответственно для баз данных PTB и MIT-BIH [29].
Недавно Сингх и Гупта использовали возможность сигнала ЭКГ для помощи в идентификации человека [8–10]. Для определения кривых ЭКГ использовались методы обработки сигналов.Результаты разграничения оказались оптимальными и стабильными по сравнению с другими опубликованными результатами. Эти разграничители использовались вместе с комплексом QRS для извлечения различных характеристик классов: временного интервала, амплитуды и угла из клинически доминирующих реперных точек на каждом сердечном сокращении электрокардиограммы. Тестовый набор из 250 записей ЭКГ, подготовленный из PhysioNet [29], оценивался с помощью предложенной системы идентификации, разработанной на сопоставлении с шаблоном и адаптивном установлении порогов. Решения о сопоставлении принимаются на основе корреляции между сохраненными учетными данными и данными тестовой ЭКГ.В результате люди классифицируются с оптимальными показателями.
2.4. Биометрическая система с поддержкой ЭКГ
Схематическое описание системы биометрической аутентификации с поддержкой ЭКГ показано на рисунке 2. Во-первых, сигнал ЭКГ получается от людей; затем для проверки качества сигнал предварительно обрабатывается, что включает в себя коррекцию сигнала от шума и артефактов. Обозначение ЭКГ включает обнаружение зубцов P, Q, R., S и T и их доминирующих реперных точек в каждом сердечном сокращении.Извлечение признаков включает определение характеристик различных классов, таких как временной интервал, амплитуда и угол, из очерченных форм сигналов. Наконец, выполняется сопоставление функций сохраненного шаблона и образца запроса для проверки личности.
Сердцебиение по записи ЭКГ обнаруживается с использованием метода [30] с некоторыми улучшениями. Он использует цифровой анализ наклона, амплитуды и ширины кривых ЭКГ. Реперные точки комплекса QRS разграничиваются в соответствии с расположением и выпуклостью пика R.Как только сердцебиение обнаружено, определяются временные окна до и после реперных точек комплекса QRS для поиска границ зубцов P и T для каждого удара ЭКГ. Подход с производной по времени и адаптивным пороговым значением используется для определения зубца P [31], в то время как зубец T определяется с использованием анализа кривизны его формы волны для каждого сердечного сокращения [32].
Для проведения биометрического эксперимента с использованием сигнала ЭКГ готовится набор функций из извлеченных реперных точек зубцов P, Q, R, S и T для каждого сердечного сокращения [10].Он содержит атрибуты интервальных, амплитудных и угловых характеристик, которые показаны на рисунках 3 (a), 3 (b) и 3 (c) соответственно. Кроме того, атрибуты, связанные с деполяризацией предсердий и реполяризацией желудочков, нормализуются [11] путем деления длины сокращений для устранения изменений, происходящих из-за изменения частоты сердечных сокращений.
Мы применяем подход статистической структуры для генерации оценок соответствия на основе сравнения векторов признаков между запросом и образцами шаблонов.Вычислительная процедура для генерации оценок соответствия на основе сравнения выборки запроса с шаблоном базы данных характеристик ЭКГ описывается следующим образом. Прежде всего, проиллюстрирована процедура создания шаблонов баз данных для разных лиц. Предположим, у человека есть набор данных ЭКГ из 𝑡 единицы времени. 𝑚 наборов подданных длины 𝑠 единицы времени (<𝑡) произвольно выбираются из полной записи ЭКГ. Затем из каждого набора подданных извлекается вектор 𝑑-признаков.Пусть 𝑃 (𝑖) - матрица паттернов, состоящая из 𝑚 векторов отдельных 𝑖 размера 𝑚 × 𝑑, которую можно определить как 𝑃 (𝑖) = ⎛⎜⎜⎜⎜⎜⎜⎝𝑓1,1𝑓1,2 ⋯ 𝑓1, 𝑑𝑓2,1𝑓2,2 ⋯ 𝑓2, 𝑑𝑓 ⋮⋮⋮ 𝑚, 1𝑓𝑚, 2 ⋯ 𝑓𝑚, 𝑑⎞⎟⎟⎟⎟⎟⎟⎠ , (1) где элемент 𝑓𝑗, 𝑘 представляет-ю характеристику 𝑗-го набора подданных. Целью произвольного выбора набора дополнительных данных является статистический анализ вариаций, присутствующих в различных сердцебиениях отдельной ЭКГ. Предположим, размер популяции составляет, поэтому существует 𝑛 различных наборов данных ЭКГ. Таким образом, в базе данных генерируется 𝑛 различных шаблонных матриц 𝑃 (𝑖), где 𝑖 = 1,2,… 𝑛.
Пусть у человека есть образец запроса 𝑄, который генерирует вектор признаков 𝑓 = ⟨𝑓1, 𝑓2,…, 𝑓𝑑⟩. Статистически расстояние между атрибутами выборки запроса и векторами признаков матрицы шаблонов индивидуума 𝑖 вычисляется с использованием евклидова расстояния следующим образом:
𝑑𝑗 (𝑖) = || 𝑓𝑗, 1 − 𝑓1 |||| 𝑓𝑗, 2 − 𝑓2 || ⋯ || 𝑓𝑗, 𝑑 − 𝑓𝑑 || , (2)
где 𝑗 = 1,2,… 𝑚. Сумма евклидовых расстояний между атрибутами векторов признаков дает оценку расстояния для отдельного 𝑖, как
𝑠𝑗 (𝑖) = 𝑑𝑘 = 1 || 𝑓𝑗, 𝑘 − 𝑓𝑘 || (3)
для всех наборов подданных 𝑗 = 1,2… 𝑚.Чтобы признать (включить) вариации, присутствующие в наборе данных ЭКГ отдельного человека, можно вычислить среднее значение оценок расстояния, обозначенное как 𝑠 (𝑖), и определить его следующим образом.
𝑠 (𝑖) = 1𝑚𝑚𝑗 = 1𝑠𝑗 (𝑖). (4)
Подлинная оценка соответствия получается при сравнении двух векторов признаков, соответствующих одному и тому же человеку, а оценка соответствия самозванца получается при сравнении векторов признаков от двух разных людей. Меньшее значение оценки расстояния указывает на хорошее совпадение, а более высокое значение оценки расстояния указывает на плохое совпадение.
2.4.1. Результаты экспериментов
Мы оценили производительность системы аутентификации с поддержкой ЭКГ в базе данных ЭКГ, подготовленной из PhysioBank [29]. База данных содержит сигналы ЭКГ 78 человек, полученные из европейской базы данных ST-T, базы данных нормального синусового ритма MIT-BIH, базы данных аритмий MIT-BIH и базы данных QT PhysioBank. В базе данных у каждого человека есть только одна запись ЭКГ; поэтому мы разделили каждую запись сигнала ЭКГ на две половины. Первая половина записей ЭКГ для каждого субъекта помечена как цель, а другая половина записей ЭКГ помечена как запрос.Производительность системы аутентификации анализируется с помощью равного коэффициента ошибок (EER). EER — это коэффициент ошибок, при котором частота мошеннических совпадений (FMR) и частота несовпадений лиц, которые должны быть правильно проверены (FNMR), принимают одно и то же значение. EER можно использовать как уникальную меру для характеристики стандарта безопасности биометрической системы. Результат EER системы аутентификации с включенной ЭКГ составляет 10,8%. Отображаются результат EER и кривая компромисса ошибок обнаружения (DET) (см.g., рисунок 4), что производительность аутентификации биометрической системы с ЭКГ низкая и вряд ли может конкурировать с традиционными биометрическими данными, такими как лицо и отпечаток пальца.
Однако сигнал ЭКГ можно комбинировать с другими биометрическими данными (например, с изображением лица и отпечатка пальца) и обеспечивать отличный источник дополнительной информации в мультибиометрической структуре. Singh et al. [27] предложили мультибиометрическую систему, которая эффективно объединяет сигнал ЭКГ с биометрическими данными лица и отпечатков пальцев.Метод слияния оценок на основе преобразования используется для оценки эффективности вышеупомянутой системы. Результаты EER унимодальных систем биометрии лица и отпечатков пальцев составляют 4,5% и 2,1%. Кривая DET, относящаяся к лучшим показателям аутентификации, достижимым с использованием мультибиометрической системы, которая объединяет сигнал ЭКГ с биометрическими данными лица и отпечатков пальцев, показана на рисунке 4. Вышеупомянутая система достигла EER 0,2%. Эти результаты подтверждают, что сигнал ЭКГ можно эффективно объединить с биометрическими данными лица и отпечатков пальцев для индивидуальной аутентификации.Хотя результат EER для сигнала ЭКГ как биометрических данных немного хуже, чем биометрические параметры лица и отпечатков пальцев, объединенная система показывает значительное улучшение результата EER.
2,5. Оценка эффективности биометрических методов с использованием ЭКГ
Методы, использующие сигнал ЭКГ в качестве биометрических данных для проверки личности, тестируются на различных наборах данных и условиях тестирования. Лабораторная демонстрация цитируемых методов показывает, что точность классификации варьируется от одного метода к другому, в то время как эффективность методов проверяется на ограниченном объеме данных.Оценки эффективности различных методов, использующих ЭКГ в качестве биометрических данных при различных условиях проверки и испытаний, приведены в таблице 1. Однако сложнее сделать какой-либо вывод об эффективности одного метода, использующего ЭКГ в качестве биометрического, по сравнению с другими методами.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2.6. Проблемы и проблемы
Биометрические методы с поддержкой ЭКГ обеспечивали индивидуальную аутентификацию посредством статистического анализа сигнала ЭКГ и выполнения количественного сравнения сигнала запроса с зарегистрированными сигналами.Эффективность предложенных методов сильно зависит от задачи представления данных, то есть от того, насколько эффективно выполняется выделение ЭКГ и определение ее формы волны. Результаты, представленные большинством методов, показали уникальность ЭКГ среди людей и поэтому могут рассматриваться как кандидат биометрических данных. Но большинство экспериментов проводится на скромных наборах данных, которые получены в контролируемых условиях в лабораторных условиях. Чтобы быть осуществимым, биометрическая система с поддержкой ЭКГ должна выполнять задачу аутентификации в широком диапазоне условий, при этом необходимо решить следующие проблемы: (1) отсутствие стандартизации функций ЭКГ, (2) вариабельность функций ЭКГ, ( 3) индивидуальность паттернов ЭКГ, (4) наследуемость волн ЭКГ, (5) настройка биометрической системы с поддержкой ЭКГ.
2.6.1. Отсутствие стандартизации характеристик ЭКГ
В большинстве методов описано, что ЭКГ как биометрический основан на временном выборе границ волн, эвристически по времени и амплитудной области. Используя информацию о локальных максимумах, минимумах и пересечениях нуля в различных масштабах, методы определяют важные реперные точки кривых ЭКГ. Эффективность этих методов зависит от точности обнаруженных реперных точек. Это открытая проблема из-за отсутствия какого-либо стандартизированного определения эффективной локализации границ волны ЭКГ.
Используя ЭКГ в качестве биометрических данных, исследователи используют знания, основанные на медицинских выводах информации ЭКГ, и они проанализировали ЭКГ на приблизительных позициях доминирующих с медицинской точки зрения реперных точек. Хотя приблизительной локализации реперных точек может быть достаточно для клинической диагностики, этого недостаточно для принятия биометрических решений. Потому что применение ЭКГ в качестве биометрических данных предполагает точную локализацию ее реперных точек волны. Небольшое изменение в локализации его реперных точек волны может вызвать неправильную классификацию больших наборов данных.
2.6.2. Вариабельность модели ЭКГ
Другой проблемой, которая ставит под сомнение использование ЭКГ в качестве потенциального кандидата биометрических данных, является вариабельность сердечного ритма. Частота сердечных сокращений зависит от физиологического и психического состояния человека. Стресс, возбуждение, физические упражнения и другие рабочие нагрузки могут влиять на частоту сердечных сокращений и повышаться. Следует отметить, что кроме изменений ритма сердцебиения, морфология ЭКГ остается неизменной. Следовательно, изменения частоты сердечных сокращений изменяют сегменты сердцебиения, такие как интервал RR, интервал PR и интервал QT.Эти функции тщательно трансформируются и освобождают от воздействия изменяющейся частоты сердечных сокращений, прежде чем использовать их в биометрических приложениях [10].
Другие факторы, такие как наличие артефакта и появление нерегулярных сокращений, могут вызывать изменения морфологии сердцебиения. Преобладающая частота помех от электросети 50–60 Гц может исказить морфологию ЭКГ, в то время как наличие эктопических или преждевременных сокращений может затруднить индивидуальную классификацию. Хотя большинство методов тестируется на ограниченном классе паттернов ЭКГ, поэтому необходимо, чтобы метод мог быть расширен на любое количество морфологически различных форм сигналов ЭКГ для жизнеспособной биометрической системы с поддержкой ЭКГ.
2.6.3. Индивидуальность рисунков ЭКГ
Проблема биометрической индивидуальности является серьезной проблемой при оценке производительности биометрической системы. Индивидуальность биометрических данных относится к вероятности межклассовой изменчивости и внутриклассового сходства шаблонов тестирования, наблюдаемых в целевой популяции. Например, индивидуальность ЭКГ указывает на то, до какой степени модели ЭКГ можно масштабировать в целевой популяции достаточно большого размера. Статистически проблема индивидуальности ЭКГ еще не решена.
В отличие от этого, возможность использования ЭКГ в качестве кандидата биометрических данных для индивидуальной аутентификации, исследованных большинством методов, была проверена на ограниченном объеме данных. Чтобы оценить крупномасштабную производительность, методы необходимо протестировать на сравнительно больших объемах данных.
2.6.4. Наследственность волн ЭКГ
Анализ наследуемости ЭКГ показал внутриклассовую корреляцию между взрослыми близнецами мужского пола (монозиготными и дизиготными). В частности, реполяризация желудочков (интервал QT) и частота сердечных сокращений (интервал RR) являются важными наследственными компонентами, в то время как комплекс QRS не показывает значимого наследственного компонента [34].
2.6.5. Настройка биометрической системы с поддержкой ЭКГ
Индивидуальная биометрическая система с поддержкой ЭКГ, аналогичная анализатору ЭКГ, который диагностирует сердечные заболевания в режиме онлайн, необходима для выполнения задачи аутентификации в различных условиях. Такое устройство размером с наручные часы или мобильный телефон, который можно закрепить или держать в руках, можно было бы использовать для регистрации и выполнения задачи аутентификации во время индивидуальной нормальной деятельности.
3. ЭЭГ как биометрический
3.1. Преамбула ЭЭГ
Электроэнцефалограмма (ЭЭГ) — это запись электрической активности мозга, которая спонтанно меняется в течение короткого периода времени. Мозг состоит из двух типов клеток: нейронов и глии [35]. Нейроны — это поляризованные белковые молекулы, переносящие заряды через мембраны мозга. ЭЭГ измеряет изменение напряжения в результате ионного потока внутри нейронов. Ганс Бергер показал, что электрическая активность мозга может быть записана неинвазивно с помощью электродов, помещенных на поверхность кожи головы [36] и названных альфа-волной.Частотный диапазон альфа — 8–12 Гц. Он возникает при закрытии глаз и при расслаблении, а ослабевает при открытии глаз или умственном напряжении. Бета-ритмы представляют собой волны низкой амплитуды с различной частотой от 12 до 30 Гц и часто связаны с активным или тревожным мышлением. Волновые структуры альфа- и бета-ритмов показаны на рисунках 5 (а) и 5 (б) соответственно.
Автоматический анализ ЭЭГ имеет множество применений в клинической нейрофизиологии. ЭЭГ — это неинвазивный инструмент, используемый для измерения вызванного потенциала и колебательных функций человеческого мозга.Вызванные потенциалы — это переходные формы волны или кратковременные нарушения текущей активности. Осцилляторные особенности ЭЭГ возникают в ответ на определенные события, которые обычно изучаются с помощью спектрального анализа. Используя способ мониторинга активности мозга, он применяется в интерфейсах мозга и компьютера с целью улучшения коммуникативных и управляющих способностей людей с двигательными нарушениями [37]. Однако извлеченные характеристики ЭЭГ можно использовать для мониторинга неврологического статуса головного мозга, в частности для выявления церебрального повреждения и ведения пациентов с неврологической травмой в отделениях интенсивной терапии.
3.2. Поддерживающие факторы
Исследования показывают, что электрическая активность мозга человека уникальна, и ЭЭГ можно использовать в качестве нового биометрического метода для проверки личности людей [12–17]. Хотя получение данных ЭЭГ несколько затруднительно. Записанный сигнал содержит шум и артефакты. Поэтому прямое использование сигнала для представления характеристик затруднено. В [16] авторы провели эксперимент для проверки людей с помощью паттернов их мозговых волн.Авторы предложили статистическую основу, основанную на моделях смеси Гаусса и максимальной апостериорной адаптации модели [38], которые используются в других традиционных биометрических системах. Они показали, что есть некоторые умственные задачи, которые больше подходят для проверки личности, чем другие.
Авторы считают, что использование ЭЭГ в качестве биометрических дает несколько преимуществ. Во-первых, это конфиденциально для человека, потому что волны ЭЭГ соответствуют его / ее умственной задаче. ЭЭГ очень сложно имитировать и практически невозможно скопировать.ЭЭГ — это отражение индивидуальных зависимых внутренних умственных задач, а аналогичные умственные задачи зависят от человека и не могут быть воспроизведены другими. Это очень безопасно, поэтому его невозможно украсть. Деятельность мозга чувствительна к стрессу и настроению человека, и нельзя заставить человека воспроизвести его / ее мысленную парольную фразу.
3.3. Связанные работы
Более раннее исследование, изучающее взаимосвязь между ЭЭГ и генетической уникальностью людей, было проведено Poulos et al.[12]. Они использовали модель авторегрессии (AR) для извлечения параметров из ЭЭГ и описали альфа-ритмы. Параметры модели AR были позже использованы в качестве функций в процессе классификации, который выполняется с использованием линейного векторного квантователя и адаптации нейронной сети. Эффективность используемой модели для идентификации людей была протестирована на наборе из 255 записей ЭЭГ, сделанных четырьмя людьми. Они сообщили о точности классификации от 72% до 80%. Paranjape et al.[13] также исследовали 349 эпох ЭЭГ, полученных от 40 человек, с использованием модели AR. Используя дискриминантный анализ, они обнаружили, что точность классификации составляет около 80%.
Паланиаппан и Мандич [14] исследовали энергию потенциалов мозга, возникающих при обработке визуальных стимулов для индивидуальной идентификации. После исследования, проведенного в [15], они проанализировали потенциал доминирующих частотных мощностей в сигналах визуального вызванного потенциала (ЗВП) гамма-диапазона как биометрических. Гамма-диапазон — это образец нервных колебаний, который может быть связан с повышенной активностью мозга, например, сознанием и остротой ума.Они выполнили преобразование сигнала с использованием фильтра с конечной импульсной характеристикой по сумме и разности и этого гамма-диапазона. Сигналы VEP извлекаются в диапазоне 3 дБ от 32 до 48 Гц. Чтобы уменьшить внутриклассовую дисперсию, присутствующую в сигнале, среднее значение фильтрованных сигналов VEP из всех каналов вычисляется с использованием следующего уравнения пространственного усреднения:
𝑥𝑥𝑛𝑇 = 𝑧𝑛𝑇 − 16161𝑖 = 1𝑧𝑖𝑛𝑇, (5)
где 𝑥𝑛𝑇 представляет собой выборку данных размера в дискретный момент времени 𝑇 и 𝑦𝑛𝑇 и 𝑧𝑛𝑇, соответственно, являются выходными сигналами фильтров суммы и разности, определяемых следующим образом:
𝑦𝑛𝑇 = 𝑀𝑗 = 0𝑀! 𝑗! (𝑀 − 𝑗)! ⋅𝑥 (𝑛 − 𝑗) 𝑇, 𝑧𝑛𝑇 = 𝑀𝑗 = 0 (−1) 𝑗𝑁! 𝑗! (𝑁 − 𝑗)! ⋅𝑦 (𝑛 −𝑗) 𝑇, (6)
где 𝑀 и 𝑁 — параметры длины фильтра, которые задаются во время эксперимента.
После предварительной обработки сигналов VEP функции извлекаются с использованием метода подпространства, называемого классификацией множественных сигналов. Наконец, извлеченные признаки классифицируются с использованием методов, основанных на k-ближайших соседях (kNN), классификаторах нейронной сети Элмана (ENN) и 10-кратной перекрестной проверочной классификации. В эксперименте было проведено в общей сложности 3560 сигналов VEP, полученных от 102 субъектов, из которых минимум 10 и максимум 50 сигналов VEP без моргания использовались для каждого субъекта.Сообщается, что точность классификации классификатора ENN является максимальной и составляет 98,12% ± 1,26% по сравнению с классификационной характеристикой kNN, которая составляет 96,13% ± 1,03%. Они утверждали, что предложенная структура четко указывает на потенциал электрической активности мозга как биометрической идентификации для проверки личности.
Марсель и Миллан [16] предложили статистическую основу, основанную на моделях смеси Гаусса (GMM) и максимальной апостериорной модели для проверки личности людей с помощью ЭЭГ.Они утверждали, что потенциал модели мозговых волн для человека является результатом строгих протоколов проверки и тестирования. Пусть 𝐱𝑇 = [𝑥1, 𝑥2,… 𝑥𝐾] будет вектором признаков, подготовленным из биометрических данных ЭЭГ, где 𝑥𝑖∈ℝ𝐾 и 𝐾 — количество признаков в каждом векторе. Пусть 𝜓𝐾∑) (𝐱; 𝜇, —вариативная гауссова функция плотности со средним и диагональной ковариационной матрицей ∑, т. Е.
𝜓𝐾 = 1𝐱; 𝜇, (2𝜋) 𝐾 / 2 || ∑ || 1 / 2 − 1exp2 (𝑥 − 𝜇) 𝑇 − 1. (𝑥 − 𝜇) (7)
С учетом GMM набора параметров (𝜆) и компонентов смеси (𝑁), оценки плотности 𝑃 (𝑥∣𝜆) могут быть получены как смесь гауссиана следующим образом:
𝑃 (𝑥∣𝜆) = 𝑁𝑗 = 1𝑤𝑗𝜓𝐾𝑥; 𝜇𝑗, 𝑗, (8)
где 𝜆 = (𝑤𝑗, 𝜇𝑗, ∑𝑗) для всех 𝑗 = 1,2,… 𝑁, 𝑤𝑗 — вес, присвоенный 𝑗-м компонентам смеси, ∑𝑁𝑗 = 1𝑤𝑗 = 1 и 𝑤𝑗≥0.Вероятность набора из 𝑛 векторов признаков 𝑋 = [𝐱𝑖] 𝑛𝑖 = 1 может быть получена как
𝑃 (𝑋𝜆) = 𝑛𝑖 = 1𝑃𝐱𝑖𝜆. (9)
Максимальное апостериорное обучение используется для оценки параметров модели (𝜆), 𝜆MAP следующим образом:
𝜆MAP = argmax𝜆𝑃 (𝑋𝜆) 𝑃 (𝜆), (10)
где 𝑃 (𝜆) — априорная плотность вероятности. Наконец, решение об аутентификации Φ (𝑋) принимается на основании вероятности требования, исходящего от подлинного пользователя 𝑃 (𝑋∣𝜆𝐺), или вероятности требования, исходящего от самозванца 𝑃 (𝑋∣𝜆𝐼), следующим образом:
Φ (𝑋) = 1, если log𝑃𝑋∣𝜆𝐺 − log𝑃𝑋∣𝜆𝐼≥𝜏0, в противном случае (11)
для некоторого порога 𝜏.
Производительность системы аутентификации измеряется на уровне половины общего коэффициента ошибок (HTER). Он определяется как среднее значение коэффициента ложного принятия (FAR) и коэффициента ложного отклонения (FRR). Для оценки возможностей предложенного метода эксперимент был проведен на наборах данных ЭЭГ, полученных от 9 здоровых испытуемых в течение 12 сеансов без обратной связи в течение трех дней. Результаты моделирования были получены на разных обучающих или проверочных наборах по различным экспериментальным протоколам.Эффективность с точки зрения HTER составляет от 6,6% до 36,2%. Наконец, они пришли к выводу, что есть некоторые умственные задачи, которые больше подходят для аутентификации личности, чем другие.
3.4. Оценка эффективности биометрических методов с использованием ЭЭГ
Большинство предлагаемых методов, которые использовали ЭЭГ в качестве биометрических, находятся на стадии лабораторной демонстрации, и их эксперименты проводятся в основном со здоровыми людьми. Лабораторная демонстрация цитируемых методов показывает, что точность классификации людей зависит от паттернов их мозговых волн, таких как альфа- и бета-волны.Оценки эффективности цитируемых методов, использующих ЭЭГ в качестве биометрических данных при различных проверках и тестовых сценариях, приведены в таблице 2. Как и в случае с другими биоэлектрическими сигналами, слишком рано делать какие-либо выводы об использовании ЭЭГ в качестве биометрических данных для людей. аутентификация. Требуется работа по разработке такой системы, которая охватывала бы людей с клиническими нарушениями, в частности людей с ограниченными физическими возможностями, для проверки их личности.
| ||||||||||||||||||||||||||||||||||||||||
3.5. Проблемы и проблемы
Обобщая результаты, полученные с помощью указанных методов, можно сделать вывод, что ЭЭГ содержит генетическую информацию, которая предлагает выбор для использования в качестве биометрических данных для индивидуальной аутентификации.Однако для получения статистически значимых результатов для биометрических приложений требуется обширный анализ ЭЭГ. Для работы жизнеспособной системы, использующей ЭЭГ в качестве биометрических данных, необходимо решить следующие ключевые вопросы: (1) размещение электродов на скальпе и сбор данных, (2) вариабельность волновых паттернов мозга, ( 3) масштабируемость для большей популяции, (4) наследуемость паттернов ЭЭГ.
3.5.1. Размещение электродов на коже черепа и сбор данных
ЭЭГ обычно включает запись с электродов на коже черепа.Основным источником ЭЭГ является синхронная активность миллионов нейронов, имеющих схожую пространственную ориентацию. В стандартной клинической практике 19 электродов равномерно размещают на коже черепа; однако количество электродов можно увеличить до 256 для получения подробной информации об источниках мозга. Поскольку ЭЭГ имеет значительно меньшее пространственное разрешение; поэтому выбор множества электродов и их размещение требует особой осторожности для записи данных с высоким пространственным разрешением, поскольку несоответствующее пространственное разделение электродов вызывает искажение предполагаемого потенциала на поверхности мозга.Кроме того, любой одиночный электрод может быть выбран в качестве опорного, а потенциал в другом месте регистрируется относительно этого опорного электрода. Следовательно, неправильный выбор ссылки часто приводит к неправильной интерпретации источника мозга.
ЭЭГ характеризуется малой амплитудой (~ мкм В) зашумленным сигналом, который находится в диапазоне 1–20 Гц. Артефакты ЭЭГ в основном генерируются телом, например, миганием глаз, движениями глаз, активацией сердца и активацией мышц. Движение объекта и размещение электродов могут вызвать появление толчков на электродах, которые возникают при мгновенном изменении.Плохое заземление электродов может привести к появлению артефактов с высокой частотой (50–60 Гц), которые трудно удалить из необработанных данных ЭЭГ. По мере увеличения уровня артефакта межклассовая вариация увеличивается (внутриклассовая вариация уменьшается), что приводит к ложному несоответствию (ложному совпадению). Полная процедура получения данных ЭЭГ требует абсолютной серьезности и полного участия испытуемых.
3.5.2. Вариабельность паттернов мозговых волн
ЭЭГ варьируется в зависимости от состояний мозга.Например, разные стадии сна, такие как быстрое движение глаз (REM) и небыстрое движение глаз (NREM), ходьба, визуализация и мышление, приводят к разной ЭЭГ. Генерируемые сигналы зрительного вызванного потенциала (ЗВП), которые, как известно, уникальны для людей, связаны с сознанием и остротой интеллекта, которые контролируются двигательной активностью экстраокулярных мышц. Следовательно, люди с тяжелыми нервно-мышечными нарушениями, у которых может отсутствовать экстраокулярная мышца, не подходят для проверки личности с использованием их паттернов ЭЭГ.
Нормальная ЭЭГ зависит от возраста. ЭЭГ новорожденных сильно отличается от ЭЭГ взрослых. ЭЭГ в детстве обычно показывает более медленные колебания частоты, чем ЭЭГ взрослых [39]. В данных, проанализированных большинством предложенных методов, отсутствует информация о разнице во времени между обучающим и тестовым наборами. Некоторые из них варьируются от секунд до пары дней. Но никаких долгосрочных различий не изучалось. Следовательно, периодическая повторная регистрация отдельной ЭЭГ может быть одной из стратегий для отслеживания долгосрочных изменений.
3.5.3. Масштабируемость для большей популяции
Потенциал электрической активности мозга как биометрического метода, исследуемый большинством методов, не соответствует размеру выборки, необходимому для определения крупномасштабных показателей. Поэтому сложно сделать однозначный вывод о том, как использовать ЭЭГ в качестве биометрических данных для проверки личности. Однако следующим шагом является проверка жизнеспособности ЭЭГ как биометрических данных на более крупных наборах данных, где необходимо исследовать различные психические состояния при разных сценариях.Хотя в исследовании [14] было экспериментально проведено в общей сложности 3560 сигналов VEP, собранных у 102 субъектов, не имеющих артефактов, индуцированных глазами, выполнимость данных ЭЭГ, собранных при нескольких состояниях мозга, включая быстрый и медленный сон, для оценки производительности каждого отдельного человека. проверка подлинности больших наборов данных еще не решена.
3.5.4. Наследуемость паттернов ЭЭГ
Спектральный анализ ЭЭГ близнецов не показал достоверных различий [40]. Однако корреляция спектров близнецов (монозиготных и дизиготных) показала внутри-парное сходство, которое значительно больше, чем межиндивидуальное сходство между неродственными людьми [41].
4. Обсуждение и будущая работа
В этой статье исследуется эффективность биоэлектрических сигналов, особенно ЭКГ и ЭЭГ, как биометрических параметров для индивидуальной аутентификации. Мы сообщили о проблемах исследования, связанных с включением ЭКГ или ЭЭГ в область биометрических приложений. Включение сигналов ЭКГ или ЭЭГ для использования в качестве биометрических данных дает несколько преимуществ. ЭКГ или ЭЭГ конфиденциальны для человека, поэтому их сложнее имитировать. Они надежно защищены, поэтому украсть их практически невозможно.Используя обычные биометрические идентификаторы, действительно сложнее разработать безопасную и надежную систему аутентификации из компонентов системы, которые не являются ни секретными, ни отзываемыми. Характеристики сигналов ЭКГ или ЭЭГ создают отличие от традиционной биометрии и предлагают альтернативу для изучения их осуществимости при разработке безопасной системы проверки личности. В качестве меры противодействия ложным атакам сигналы ЭКГ или ЭЭГ обладают унаследованной особенностью жизнеспособности. Это означает, что физиологические характеристики ЭКГ или ЭЭГ не могут быть приобретены, если человек не жив.
Хотя лабораторная демонстрация использования сигналов ЭКГ или ЭЭГ для аутентификации людей показывает умеренную производительность, они могут дополнять другие биометрические данные в мультибиометрической структуре для надежной аутентификации. В частности, мы показали, что сигнал ЭКГ можно комбинировать с биометрической информацией о лице и отпечатках пальцев с оптимальной производительностью системы. В заключение, плодотворные направления дальнейших исследований могут включать следующее. (1) Чтобы быть осуществимым, система, использующая биоэлектрические сигналы в качестве биометрии, должна выполнять задачу аутентификации в широком диапазоне условий и в большей популяции, тогда как данные собираются в большие временные интервалы.(2) Эффективность большинства методов, использующих ЭКГ или ЭЭГ в качестве биометрических данных, была проверена на здоровых людях. Следовательно, следует предпринять усилия для изучения возможности распространения предлагаемых схем на нездоровых субъектов. (3) Важно выяснить, до какой степени ЭКГ или ЭЭГ варьируются в зависимости от эмоций или различных состояний ума. Частота сердечных сокращений варьируется в зависимости от различных раздражителей. Характер мозговых волн варьируется в зависимости от состояний нейронов. Исследование устойчивости к субъектам с различными психическими и эмоциональными состояниями необходимо для подтверждения результатов большинства методов.(4) Сбор данных о биоэлектрических сигналах, в частности, ЭЭГ, является сложной задачей. Размещение электродов в правильном положении, например неравномерное или несоответствующее пространственное разделение, может вызвать искажение полученного сигнала. Следовательно, правильное размещение электродов требует дополнительных знаний и понимания физиологии мозга и сердца для записи сигналов ЭЭГ и ЭКГ соответственно. (5) Методы анализа сигналов ЭКГ или ЭЭГ все еще находятся в зачаточном состоянии. Необходимо исследование альтернативных методов классификации, которые достаточно надежны для обработки вариаций, присутствующих в функциях.Потребуются особые усилия, чтобы проверить качество внезапно полученных данных, что может быть связано с отказом от сотрудничества со стороны пользователя или наличием некоторых артефактов. Далее требуются методы представления данных, которые будут работать в рамках нестандартных функций. (6) Чтобы отнести сигналы ЭКГ или ЭЭГ к категории менее навязчивой биометрии, для регистрации и аутентификации требуется настраиваемая система размером с мобильный телефон. . Это упростит не только процесс аутентификации, но и выполнение функций во время обычных действий людей.
Электрические сигналы, которые делают человеческий мозг уникальным
Нейроны в человеческом мозге получают электрические сигналы от тысяч других клеток, и длинные нейронные отростки, называемые дендритами, играют решающую роль во включении всей этой информации, чтобы клетки могли реагировать соответствующим образом.
Используя труднодоступные образцы ткани человеческого мозга, нейробиологи Массачусетского технологического института обнаружили, что дендриты человека обладают электрическими свойствами, отличными от дендритов других видов.Их исследования показывают, что электрические сигналы ослабевают сильнее, когда они проходят по дендритам человека, что приводит к более высокой степени электрической компартментализации, а это означает, что небольшие участки дендритов могут вести себя независимо от остальной части нейрона.
Эти различия могут способствовать увеличению вычислительной мощности человеческого мозга, говорят исследователи.
«Люди умны не только потому, что у нас больше нейронов и крупнее кора головного мозга. Снизу вверх нейроны ведут себя по-разному, — говорит Марк Харнетт, доцент кафедры мозговых и когнитивных наук Фреда и Кэрол Миддлтон.«В нейронах человека больше электрического разделения, и это позволяет этим единицам быть немного более независимыми, что потенциально приводит к увеличению вычислительных возможностей отдельных нейронов».
Харнетт, который также является членом Института исследований мозга Макговерна Массачусетского технологического института, и Сидней Кэш, доцент кафедры неврологии Гарвардской медицинской школы и Массачусетской больницы общего профиля, являются старшими авторами исследования, которое будет опубликовано 18 октября. выпуск Cell. Ведущий автор статьи — Лу Больё-Ларош, аспирант факультета мозга и когнитивных наук Массачусетского технологического института.
Нейронные вычисления
Дендриты можно рассматривать как аналог транзисторов в компьютере, выполняющих простые операции с использованием электрических сигналов. Дендриты получают входные данные от многих других нейронов и передают эти сигналы телу клетки. При достаточной стимуляции нейрон генерирует потенциал действия — электрический импульс, который затем стимулирует другие нейроны. Большие сети этих нейронов взаимодействуют друг с другом, чтобы генерировать мысли и поведение.
Структура отдельного нейрона часто напоминает дерево, со множеством ветвей, несущих информацию, которая поступает далеко от тела клетки.Предыдущие исследования показали, что сила электрических сигналов, поступающих в тело клетки, частично зависит от того, как далеко они проходят по дендриту, чтобы добраться туда. По мере распространения сигналов они ослабевают, поэтому сигнал, поступающий далеко от тела клетки, оказывает меньшее влияние, чем сигнал, поступающий рядом с телом клетки.
Дендриты в коре головного мозга человека намного длиннее, чем у крыс и большинства других видов, потому что кора головного мозга человека стала намного толще, чем у других видов.У людей кора головного мозга составляет около 75 процентов от общего объема мозга по сравнению с примерно 30 процентами в мозге крысы.
Хотя кора головного мозга человека в два-три раза толще, чем у крыс, она поддерживает ту же общую организацию, состоящую из шести отличительных слоев нейронов. Нейроны из слоя 5 имеют дендриты, достаточно длинные, чтобы достичь уровня 1, а это означает, что человеческие дендриты должны были удлиняться по мере развития человеческого мозга, а электрические сигналы должны проходить гораздо дальше.
В новом исследовании команда Массачусетского технологического института хотела изучить, как эти различия в длине могут повлиять на электрические свойства дендритов. Они смогли сравнить электрическую активность дендритов крысы и человека, используя небольшие кусочки мозговой ткани, взятые у пациентов с эпилепсией, перенесших хирургическое удаление части височной доли. Чтобы добраться до пораженной части мозга, хирурги также должны удалить небольшой кусок передней височной доли.
С помощью сотрудников MGH Кэша, Мэтью Фроша, Зива Уильямса и Эмада Эскандара лаборатория Харнетта смогла получить образцы передней височной доли, каждая размером с ноготь.
По словам Харнетта, данные свидетельствуют о том, что передняя височная доля не поражена эпилепсией, и ткань кажется нормальной при исследовании с помощью нейропатологических методов. Эта часть мозга, по-видимому, участвует во множестве функций, включая язык и визуальную обработку, но не является критичной для какой-либо одной функции; пациенты могут нормально функционировать после его удаления.
После удаления ткани исследователи поместили ее в раствор, очень похожий на спинномозговую жидкость, с проходящим через него кислородом.Это позволило им сохранить живую ткань до 48 часов. В то время они использовали методику, известную как электрофизиология патч-зажима, чтобы измерить, как электрические сигналы проходят по дендритам пирамидных нейронов, которые являются наиболее распространенным типом возбуждающих нейронов в коре головного мозга.
Эти эксперименты были выполнены в основном Больё-Ларош. Лаборатория Харнетта (и другие) ранее проводила подобные эксперименты на дендритах грызунов, но его команда первой проанализировала электрические свойства дендритов человека.
Уникальные особенности
Исследователи обнаружили, что, поскольку человеческие дендриты покрывают большие расстояния, сигнал, проходящий по дендриту человека от слоя 1 к телу клетки в слое 5, намного слабее, чем сигнал, проходящий по дендриту крысы из слоя 1 к слою 5.
Они также показали, что дендриты человека и крысы имеют одинаковое количество ионных каналов, которые регулируют ток, но эти каналы имеют более низкую плотность в дендритах человека в результате удлинения дендритов.Они также разработали подробную биофизическую модель, которая показывает, что это изменение плотности может объяснить некоторые различия в электрической активности дендритов человека и крысы, говорит Харнетт.
Нельсон Спрастон, старший директор научных программ Медицинского института Говарда Хьюза в исследовательском кампусе Джанелии, назвал проведенный исследователями анализ человеческих дендритов «замечательным достижением».
«На сегодняшний день это наиболее подробные измерения физиологических свойств человеческих нейронов», — говорит Спрастон, не принимавший участия в исследовании.«Подобные эксперименты технически сложны даже на мышах и крысах, поэтому с технической точки зрения довольно удивительно, что они сделали это на людях».
Остается вопрос, как эти различия влияют на интеллект человека? Гипотеза Харнетта заключается в том, что из-за этих различий, которые позволяют большему количеству участков дендрита влиять на силу входящего сигнала, отдельные нейроны могут выполнять более сложные вычисления с информацией.
«Если у вас есть кортикальный столбик, в котором есть часть коры головного мозга человека или грызунов, вы сможете быстрее выполнять больше вычислений с архитектурой человека по сравнению с архитектурой грызунов», — говорит он.
Есть много других различий между человеческими нейронами и нейронами других видов, добавляет Харнетт, что затрудняет выявление эффектов электрических свойств дендритов. В будущих исследованиях он надеется дополнительно изучить точное влияние этих электрических свойств и то, как они взаимодействуют с другими уникальными особенностями человеческих нейронов для увеличения вычислительной мощности.
Статья переиздана по материалам MIT. Примечание: материал мог быть отредактирован по объему и содержанию.Для получения дополнительной информации, пожалуйста, свяжитесь с цитируемым источником.
Ссылка: Beaulieu-Laroche, L., Toloza, E.H.S., Goes, M.-S. ван дер, Лафуркад, М., Барнаджан, Д., Уильямс, З. М.,… Харнетт, М. Т. (2018). Повышенная дендритная компартментализация в корковых нейронах человека. Ячейка, 175 (3), 643-651.e14. https://doi.org/10.1016/j.cell.2018.08.045
Понимание электрических сигналов из-под поверхности Земли
Ученым давно известно, что различные типы горных пород по-разному проводят ток и что эти различия становятся еще более заметными по мере того, как температура и давление возрастают под поверхностью Земли.Они также знают, что необычные изменения в электропроводности могут сигнализировать об активности внизу, например, о миграции магмы или выбросе захваченных жидкостей.
Таким образом, электрические измерения могут раскрыть ключи к разгадке событий, которые вызывают землетрясения и извержения вулканов здесь, на поверхности. Они также могут дать ключ к разгадке структуры и динамики мантии. Однако интерпретировать эти сигналы далеко не так просто.
Земные ученые все чаще используют электрические наблюдения, сделанные в полевых условиях, для получения изображений земной коры и мантии, в частности, в зонах субдукции и срединно-океанических хребтах.Эффективные средства интерпретации этих электрических изображений и помещения их в контекст с другими геологическими наблюдениями являются ключом к преобразованию исходных данных в полезные знания. Такие знания включают оценку потенциальных опасностей путем исследования, например, связей между выбросом флюидов и генерацией землетрясений или производством и переносом магматического расплава от области его источника до возможного извержения.
SIGMELTS — это бесплатное приложение, которое помогает характеризовать электропроводящие или резистивные элементы, обнаруженные на глубине, с помощью электромагнитных наблюдений.Цель этого веб-приложения — облегчить разработку моделей электрических свойств материалов коры и мантии, которые, в свою очередь, используются для улучшения интерпретации полевых электромагнитных наблюдений. Доступна новая версия SIGMELTS.
От электрических токов к свойствам материалов
Земные ученые могут использовать SIGMELTS, чтобы связать свойства проводимости с известными свойствами различных горных пород, минералов, водных флюидов и магматических расплавов.Способность различных типов горных пород и минералов проводить электрические токи зависит от их химических и физических свойств. Таким образом, изменения электропроводности могут быть индикаторами изменений минерального состава от одной области к другой или того, является ли материал в данной области твердым или полностью или частично расплавленным. Ученые-геологи могут использовать SIGMELTS для обработки и интерпретации необработанных данных о проводимости с месторождения, связывая свойства проводимости с известными свойствами различных горных пород, минералов, водных флюидов и магматических расплавов.
Выходные данные SIGMELTS помогают пользователям интерпретировать аномалии электропроводности, наблюдаемые в данных, собранных в результате магнитотеллурических исследований и электромагнитных исследований с контролируемым источником в различных тектонических условиях, таких как срединно-океанические хребты и зоны субдукции. Эти аномалии обычно согласуются с результатами петрологических исследований, которые предсказывают выделение водных флюидов или присутствие частичного таяния.
Новые функции и возможности
SIGMELTS 2.0 имеет значительно расширенные возможности по сравнению с SIGMELTS 1.0 [ Pommier and LeTrong, 2011]. В SIGMELTS 1.0 отсутствовали важные функции, в том числе возможность построения и экспорта вычисленных данных. В дополнение к новой, более удобной компоновке и обновленной и расширенной базе данных по электричеству, SIGMELTS 2.0 теперь предлагает пользователю возможность визуализировать вычисленные значения проводимости на графике и экспортировать результаты (в формате CSV) для использования вне приложения ( Фигура 1).
Мы также добавили обучающую вкладку, чтобы предоставить пользователям некоторые инструкции по использованию SIGMELTS.Кроме того, каждая вкладка на веб-сайте содержит четкое изложение экспериментальной ошибки, полученной как в результате полевых, так и лабораторных измерений и расчетов. Пользователям настоятельно рекомендуется обращаться к источникам литературы, перечисленным в нижней части вкладок и используемым в базе данных SIGMELTS.
Новая вкладка «мантия» позволяет пользователям легко вычислять электрическую проводимость объемной, многофазной, возможно, частично расплавленной мантийной породы, минералогия которой определяется пользователем (рис. 1). Эта опция позволяет пользователям вычислять любые расчеты электропроводности для реалистичных композиций.При желании пользователи могут архивировать свои расчеты для доступа других пользователей приложения. Такой краудсорсинговый подход должен помочь улучшить интерпретацию электрических аномалий, обнаруженных в верхней мантии.
Рис. 1. Веб-интерфейс SIGMELTS 2.0. В текстовом поле ввода отображаются параметры ввода для расчета проводимости объемной мантии. Расчетный график помогает визуализировать вычисления и может быть загружен.
Не только оливин
Включение реальных минеральных составов во вкладку мантии — это один из способов, которым SIGMELTS 2.0 преодолевает существенные ограничения предыдущих методов моделирования. Модели проводимости верхней мантии часто основывались на минералогии только оливина, хотя недавние исследования показывают, что оливин, возможно, не является самым распространенным минералом в некоторых регионах верхней мантии. Модели только для оливина предполагают, что вкладом других минералов (таких как пироксены и гранат) в электрический отклик мантии можно пренебречь, но такое упрощение может привести к значительной неопределенности в оценках температуры мантии, полученных на основе проводимости.
В случае сухой мантии (с небольшим количеством воды или без нее) разница в расчетной проводимости между мантией, состоящей только из оливина, и трехфазной мантией (перидотит, порода, которая обычно содержит в основном оливин, с меньшими количествами пироксены и гранат) в верхней мантии невелика, но эта разница увеличивается с температурой, которая увеличивается с глубиной (рис. 2, зеленая и оранжевая кривые). Это связано с тем, что гранат, один из минералов перидотита, является лучшим проводником, чем оливин.
Рис. 2. Влияние химического состава мантии и распределения воды на электропроводность (в сименсах на метр) при давлении около 2 гигапаскалей, что соответствует глубине 60 километров. Порода верхней мантии (перидотит) соответствует смеси оливина (ol), пироксена (pyr) и граната (gt). Коэффициент распределения воды D между пироксеном и оливином взят из Hauri et al. [2006]. Пунктирными линиями показаны различные оценки температуры, полученные в результате рассмотрения различного распределения воды между минералами мантии.Для влажной мантии наблюдаются сильные различия проводимости, в основном зависящие от распределения воды. Расчеты для объемного содержания воды 200 частей на миллион показывают, что проводимость водного оливина аналогична проводимости водного перидотита, если эта вода остается в основном в фазе оливина (рис. 2, синяя и пурпурная кривые).
Однако, если мы учитываем более реалистичное разделение воды, рассчитанная нами объемная электропроводность уменьшается примерно в 2 раза в исследуемом диапазоне температур (рис. 2, красная кривая).Это приводит к значительному изменению оценки температуры мантии. Например, предположение о величине объемной проводимости 0,03 симена на метр в составе мантии, состоящей только из водного оливина или водного перидотита с водой, содержащейся только в оливине, дает расчетную температуру мантии около 1220–1230 ° C (рис. 2, синие и фиолетовые кривые). Учет распределения воды между оливином и пироксеном дает значение 1330 ° C (рис. 2, красная кривая).
В качестве альтернативы, если мы предположим, что нам известна температура мантии, то значение проводимости можно использовать для оценки степени гидратации мантии.Например, учитывая температуру мантии 1300 ° C, значение объемной проводимости более 0,01 симена на метр предполагает гидратацию мантии (рис. 2).
Эти простые вычисления подчеркивают важность сочетания геохимических ограничений и петрологических знаний с электрическими измерениями.
Использование SIGMELTS 2.0
Пользователи должны понимать петрологию и тектонику интересующей области, потому что значение аномалии проводимости редко может быть однозначно интерпретировано только с точки зрения проводимости.Пользователи SIGMELTS должны понимать петрологию и тектонику интересующей области, потому что значение аномалии проводимости редко может быть однозначно интерпретировано только с точки зрения проводимости. Например, различия в проводимости частичных расплавов и флюидов в значительной степени зависят от их температуры и состава.
Например, в зоне субдукции Каскадия вдоль западного побережья Северной Америки рядом с плитой на глубине 80 км наблюдается аномалия проводимости ~ 1 симен на метр.Эта аномалия согласуется с высвобождением водных флюидов из плиты и образованием расплава [ McGary et al., 2014].
Однако выяснение влияния температуры и состава на объемную проводимость водных флюидов и расплавов требует включения информации о составе флюидной фазы из петрологических исследований и температурного поля из численных моделей [ Pommier and Evans, 2017]. Знание термической структуры рассматриваемой области поля часто имеет решающее значение для интерпретации электрических аномалий, поскольку проводимость очень чувствительна к температуре.Более того, стабильность минералов, а также начало плавления сильно зависят от теплового состояния.
Пример места, где исследования включают термическую структуру в интерпретацию электрических аномалий, находится вдоль Восточно-Тихоокеанского поднятия. Петрологические исследования (рис. 3, в центре) дают представление о различных материалах, ожидаемых ниже срединно-океанического хребта. Вычисленные значения проводимости (рис. 3, справа) для этих материалов хорошо согласуются с электрическим профилем, измеренным в полевых условиях (рис. 3, слева).
Короче говоря, SIGMELTS 2.0 упрощает интерпретацию данных о проводимости и интеграцию результатов с результатами петрологических исследований и компьютерного моделирования. В результате земные ученые могут построить более подробное и точное представление геологических процессов далеко под поверхностью Земли.
Рис. 3. Пример интерпретации полевого электрического изображения: (слева) электрическое изображение ниже Восточно-Тихоокеанского поднятия, полученное по магнитотеллурическим данным морского дна [ Key et al. , 2013], (в центре) петрологический вид срединно-океанического хребта [по Dasgupta et al., 2013], на котором показаны ожидаемые материалы на разных глубинах, и (справа) вид на срединно-океанический хребет, рассчитанный на основе лабораторных измерений с использованием SIGMELTS 2.0. Химический состав материалов и температурные ограничения взяты из петрологических исследований.
Благодарности
SIGMELTS 2.0 размещается и обслуживается в Институте геофизики и планетной физики (IGPP), Институте океанографии Скриппса. Те, кто хочет узнать больше о SIGMELTS, могут связаться с нами по адресу [адрес электронной почты защищен].Мы благодарим Роба Эванса, Микаэля Лаумонье и Стива Констебля за отзывы во время разработки SIGMELTS 2.0, а также Керри Ки и анонимного рецензента за полезные комментарии. Эта работа поддержана IGPP Green Foundation и Национальным научным фондом (EAR-1461594, EAR-1551200).
Что такое сигнал? Типы сигналов, их свойства, действие и применение
Что такое сигнал, классификация сигналов и роль сигналов в цифровой связи
Что такое сигнал?
Сигнал определяется как любое физическое или виртуальное количество , которое изменяется в зависимости от времени или пространства или любой другой независимой переменной или переменных.
Графически независимая переменная представлена горизонтальной осью , или осью x. А зависимая переменная представлена вертикальной осью , или осью y.
Математически сигнал является функцией одной или нескольких независимых переменных.
Сигнал с одной переменной
Зависит от одной независимой переменной. Он изменяется линейно или нелинейно в зависимости от выражения сигнала.Примеры сигнала с одной переменной:
S (x) = x + 5
S (x) = x 2 +5 Где x — это переменная
S (t) = cos (wt + ϴ) Где t — переменная
Сигнал с двумя переменными
Сигнал с двумя переменными изменяется с изменением двух независимых переменных. Пример сигнала с двумя переменными:
S (x, y) = 2x + 5y
Характеристики сигнала
Сигнал определяется своими характеристиками.Он показывает характер сигнала. Эти характеристики приведены ниже:
Амплитуда
Амплитуда — это сила или высота формы сигнала. Визуально это высота сигнала от его центральной линии или оси x. Ось Y формы сигнала показывает амплитуду сигнала. Амплитуда сигнала меняется со временем.
Например, амплитуда синусоидальной волны — это максимальная высота формы волны по оси Y.
Уровень сигнала обычно измеряется в децибел db .
Частота
Частота — это частота повторения формы волны сигнала в секунду.
Периодические сигналы повторяют свой цикл через некоторое время. Число циклов в секунду известно как Частота . Единица частоты — герц (Гц) и один герц равен один цикл в секунду. Он измеряется по оси x формы сигнала.
Например, синусоида 5 герц завершит свои 5 циклов за одну секунду .
Время Период
Период времени сигнала — это время, за которое он завершает свой полный цикл. Единица периода — Секунда . Период времени обозначается ‘T’ , и это , инверсия частоты . Т.е.
T = 1 / F
Например, синусоидальная волна с периодом времени 10 секунд завершит свое за один полный цикл за 10 секунд .
Фаза
Фаза синусоидального сигнала — это сдвиг или сдвиг в его исходной или начальной точке. Фазовый сдвиг может быть с отставанием или с опережением . Обычно исходные синусоидальные сигналы имеют фазу 0 ° градусов и начинаются с нулевой амплитуды, но сдвиг по фазе сдвигает его начальную амплитуду на значение, отличное от 0.
Пример сдвига фазы 45 ° приведен ниже.Сигнал остается прежним, но его источник смещен на 45 ° .
Фазовый сдвиг может составлять от 0 ° до 360 ° с шагом градусов или 0 до 2π с шагом радиан . 360 ° градусов или 2π радиан — это один полный период.
Связанное сообщение: Что такое GSM и как он работает?
Размер сигнала
Размер сигнала — это число, которое показывает мощность или ширину этого сигнала. Как мы знаем, амплитуда сигнала меняется во времени.Из-за этого изменения мы не можем сказать, что его амплитуда может быть его размером. Чтобы измерить размер сигнала, мы должны принять во внимание область , покрытую , по амплитуде сигнала в пределах временной длительности.
В зависимости от размера сигнала есть два параметра.
Энергия сигнала
Энергия сигнала — это площадь сигнала под его кривой. Но сигнал может быть как в положительной, так и в отрицательной области. Из-за этого они будут нейтрализовать влияние друг друга, что приведет к уменьшению сигнала.Чтобы устранить эту проблему, мы берем квадратов амплитуды сигнала , которая всегда положительна.
Для сигнала g (t) площадь под g 2 (t) известна как энергия сигнала .
Единица энергии сигнала
Эта энергия не принимается в обычном понимании, но показывает размер сигнала. Следовательно, его единица измерения — не джоуль. Единица энергии зависит от сигнала .Если это сигнал напряжения , то его единица будет вольт 2 / сек .
Ограничение
Энергию сигнала можно измерить, только если сигнал имеет конечное значение . Бесконечный сигнал будет иметь бесконечную энергию , что абсурдно. Амплитуда конечного сигнала переходит в 0 , когда время (t) приближается к бесконечности (∞).
Итак, необходимо , чтобы сигнал был конечным сигналом , если вы хотите измерить его энергию.
Мощность сигнала
Если сигнал представляет собой бесконечный сигнал , то есть его амплитуда не переходит в 0 , когда время t приближается к ∞ , мы не можем измерить его энергию. В таком случае мы берем среднее по времени ( период времени ) энергии сигнала в качестве мощности сигнала.
Единица мощности
Подобно энергии сигнала, эта мощность также не принимается в общепринятом смысле.Также будет зависеть от от измеряемого сигнала . Если сигнал сигнал напряжения , тогда мощность будет в вольт 2 .
Ограничение
Как и в случае с энергией сигнала, измерение мощности сигнала также имеет некоторые ограничения: сигнал должен иметь периодическую природу . бесконечный и непериодический сигнал не имеют ни энергии , ни мощности .
Классификация сигналов
Сигналы классифицируются по различным категориям в зависимости от их характеристик. Некоторые из этих категорий приведены ниже.
Аналог. Цифровой сигнал
Сигнал можно разделить на аналоговую или цифровую категорию на основе их амплитуды . Эта классификация основана только на вертикальной оси (амплитуда) сигнала. И это не имеет никакого отношения к горизонтальной оси (оси времени).
Амплитуда аналогового сигнала может иметь любое значение (включая доли) в любой момент времени. Это означает, что аналоговый сигнал имеет бесконечных значений .
Однако амплитуда цифрового сигнала может иметь только конечных и дискретных значений.
Особый случай цифрового сигнала , имеющий двух дискретных значений , известен как двоичный сигнал .Однако количество значений амплитуды в цифровом сигнале составляет , не ограничивается , до только двумя .
Аналоговый сигнал преобразуется в цифровой с помощью аналого-цифрового преобразователя (АЦП) .
Сигнал непрерывного и дискретного времени
Эта классификация основана на горизонтальной оси (оси времени) сигнала.
Непрерывные и дискретные временные сигналы не следует путать с аналоговыми и цифровыми сигналами соответственно.
Сигнал непрерывного времени — это сигнал, значение (амплитуда) которого существует для каждой дроби из времени t.
Сигнал дискретного времени существует только для дискретного значения из времени t .
Помните , нет ограничения на амплитуды сигнала. Поэтому его не следует путать с аналоговым или цифровым сигналом.
Энергия противСигнал мощности
Сигнал Сигнал энергии , если его амплитуда переходит в 0 , когда время приближается к ∞ . Энергетические сигналы имеют конечную энергию.
Аналогично, сигнал с конечной мощностью известен как сигнал мощности. Сигнал мощности является периодическим сигналом , то есть имеет период времени.
Сигнал Energy имеет конечной энергии , но нулевой мощности .А сигнал мощности имеет конечной мощности , но бесконечной энергии . Таким образом, сигнал может быть энергетическим сигналом или энергетическим сигналом , но не может быть одновременно .
Бесконечный сигнал , не имеющий периодической природы не является ни , ни сигналом мощности .
Периодические Vs. Апериодический сигнал
Периодический сигнал — это сигнал, который заставляет повторять свой шаблон после минимального фиксированного времени .Это время известно как Период времени «T» этого сигнала. Периодический сигнал не изменяется, если он сдвинут во времени на любое кратное временному периоду «T».
Математическое выражение для периодического сигнала g (t) :
T 0 — это период времени сигнала g (t) .
Периодический сигнал начинается с t = -∞ и продолжается до t = + ∞ . Сигнал, который начинается с t = 0 , не будет таким же сигналом, если он сдвинут по времени на + T, потому что он не существовал для отрицательных t.
Апериодический или непериодический сигнал — это сигнал, который не повторяет сам по себе через определенное время. У этих сигналов нет повторов ни одного паттерна.
Детерминированный и случайный сигнал
Сигнал, который может быть представлен в математической или графической форме , называется детерминированным сигналом . Детерминированные сигналы имеют заданную амплитуду , частоту и т. Д.Их легко обрабатывать, поскольку они определены в течение длительного периода времени, и мы можем Оценить его результат , если они применяются к конкретной системе на основе ее выражения.
Сигнал random или , недетерминированный — это сигнал, который может быть представлен только в вероятностном выражении , а не в его полном математическом выражении. Каждый сигнал, имеющий некоторую неопределенность , является случайным сигналом . Шумовой сигнал — лучший пример случайного сигнала.
Как правило, каждый сигнал сообщения является случайным сигналом, потому что мы не уверены в том, какая информация будет передана другой стороне.
Операция сигнала
Некоторые основные операции сигналов приведены ниже
Сдвиг по времени
Сдвиг по времени означает перемещение сигнала по оси времени ( горизонтальная ось ).Временной сдвиг в сигнале не изменяет сам сигнал, а только сдвигает источник сигнала от его исходной точки по оси времени.
По сути, сложение по времени — это сдвиг во времени. Для сдвига по времени сигнал g (t) , t следует заменить на (t-T) , где T — секунды сдвига во времени . Следовательно, g (t-T) — это сигнал со сдвигом во времени на T секунд.
Временной сдвиг может быть сдвиг вправо (задержка) или сдвиг влево (вперед).
Если сдвиг по времени T равен положительным , то сигнал сместится на вправо (задержка). Например, сигнал g (t-4) — это сдвинутая версия g (t) с задержкой 4 секунд .
Если временной сдвиг T равен отрицательным , то сигнал сместится на влево (вперед). Сигнал g (t + 4) — это сдвинутая версия g (t) с 4 секунд до слева .
Масштабирование времени
Масштабирование времени сигнала означает сжатие или расширение сигнала. Это достигается умножением временной переменной сигнала на коэффициент . Сигнал расширяется или сжимается в зависимости от фактора.
Предположим, что сигнал g (t) , тогда его масштабированная версия составляет g (at) .
Если коэффициент a> 1 , то сигнал сжимает .И операция называется сжатием сигнала . Сжатие сигнала сделает сигнал быстрым , поскольку он станет меньше и его временная длительность станет меньше.
Если a <1 , то сигнал расширится . И операция называется расширением сигнала .
После масштабирования исходная точка сигнала остается неизменной. Расширение сигнала приведет к замедлению сигнала , поскольку он становится шире и покрывает большую временную продолжительность.
Инверсия времени
При инверсии времени сигнал перевернут вокруг оси y (вертикальная ось). Результирующий сигнал представляет собой зеркальное отображение исходного сигнала.
Инверсия времени — это частный случай масштабирования времени , в котором коэффициент a = -1 . Поэтому, чтобы инвертировать сигнал , мы заменяем (t) на (-t) .
Математически инверсия времени сигнала g (t) равна g (-t).
Похожие сообщения
Аналоговые и цифровые сигналы: использование, преимущества и недостатки | Статья
.
Получайте ценные ресурсы прямо на ваш почтовый ящик — рассылается раз в месяц
Мы ценим вашу конфиденциальность
Сигнал — это электромагнитный или электрический ток, по которому данные передаются из одной системы или сети в другую. В электронике сигнал часто представляет собой изменяющееся во времени напряжение, которое также является электромагнитной волной, несущей информацию, хотя он может принимать другие формы, например ток.В электронике используются два основных типа сигналов: аналоговые и цифровые. В этой статье обсуждаются соответствующие характеристики, использование, преимущества и недостатки, а также типичные применения аналоговых и цифровых сигналов.
Аналоговый сигнал
Аналоговый сигнал изменяется во времени и обычно привязан к диапазону (например, от +12 В до -12 В), но в этом непрерывном диапазоне существует бесконечное количество значений. Аналоговый сигнал использует данное свойство среды для передачи информации о сигнале, например, электричество, движущееся по проводу.В электрическом сигнале напряжение, ток или частота сигнала могут изменяться для представления информации. Аналоговые сигналы часто представляют собой рассчитанные реакции на изменения света, звука, температуры, положения, давления или других физических явлений.
При построении графика зависимости напряжения от времени аналоговый сигнал должен давать плавную и непрерывную кривую. Никаких дискретных изменений значения не должно быть (см. Рисунок 1) .
Рисунок 1: Аналоговый сигнал
Цифровой сигнал
Цифровой сигнал — это сигнал, который представляет данные как последовательность дискретных значений.Цифровой сигнал может принимать только одно значение из конечного набора возможных значений в данный момент времени. В случае цифровых сигналов физическая величина, представляющая информацию, может быть различной:
- Переменный электрический ток или напряжение
- Фаза или поляризация электромагнитного поля
- Акустическое давление
- Намагничивание магнитного носителя информации
Цифровые сигналы используются во всей цифровой электронике, включая вычислительное оборудование и устройства передачи данных.При нанесении на график зависимости напряжения от времени цифровые сигналы имеют одно из двух значений и обычно находятся в диапазоне от 0 В до VCC (обычно 1,8 В, 3,3 В или 5 В) (см. Рисунок 2) .
Аналоговая электроника
Большинство основных электронных компонентов — резисторы, конденсаторы, катушки индуктивности, диоды, транзисторы и операционные усилители (операционные усилители) — по своей сути являются аналоговыми компонентами. Схемы, построенные из комбинации этих компонентов, представляют собой аналоговые схемы (см. Рисунок 3) .
Рисунок 3: Аналоговая схема
Аналоговые схемы могут иметь сложную конструкцию с несколькими компонентами или могут быть простыми, например, два резистора, образующие делитель напряжения. Как правило, аналоговые схемы проектировать сложнее, чем цифровые схемы, выполняющие ту же задачу. Для разработки аналогового радиоприемника или аналогового зарядного устройства для батареи потребуется проектировщик, знакомый с аналоговыми схемами, поскольку для упрощения этих конструкций были приняты цифровые компоненты.
Аналоговые схемы обычно более восприимчивы к шуму, а «шум» — это любые небольшие нежелательные изменения напряжения.Небольшие изменения уровня напряжения аналогового сигнала могут привести к значительным ошибкам при обработке.
Аналоговые сигналы обычно используются в системах связи, которые передают голос, данные, изображение, сигнал или видеоинформацию с использованием непрерывного сигнала. Существует два основных типа аналоговой передачи, которые основаны на том, как они адаптируют данные для комбинирования входного сигнала с сигналом несущей. Двумя методами являются амплитудная модуляция и частотная модуляция. Амплитудная модуляция (AM) регулирует амплитуду несущего сигнала.Частотная модуляция (FM) регулирует частоту несущего сигнала. Аналоговая передача может быть достигнута многими способами:
- Через витую пару или коаксиальный кабель
- Через оптоволоконный кабель
- По радио
- Через воду
Подобно тому, как человеческое тело использует глаза и уши для захвата сенсорной информации, аналоговые схемы используют эти методологии для взаимодействия с реальным миром, а также для точного захвата и обработки этих сигналов в электронике.
MPS производит множество аналоговых ИС и компонентов, таких как MP2322, синхронный понижающий преобразователь с низким I Q в крошечном корпусе QFN размером 1,5 x 2 мм.
Цифровая электроника
Цифровые схемы реализуют такие компоненты, как логические вентили или более сложные цифровые ИС. Такие ИС представлены прямоугольниками с выходящими из них выводами (см. Рисунок 4) .
Рисунок 4: Цифровая схема
Цифровые схемы обычно используют двоичную схему.Хотя значения данных представлены только двумя состояниями (0 и 1), более крупные значения могут быть представлены группами двоичных битов. Например, в 1-битной системе 0 представляет значение данных 0, а 1 представляет значение данных 1. Однако в 2-битной системе 00 представляет 0, 01 представляет 1, 10 представляет 2, а 11 представляет 3. В 16-битной системе наибольшее число, которое может быть представлено, равно 216 или 65 536. Эти группы битов могут быть захвачены либо как последовательность последовательных битов, либо через параллельную шину.Это позволяет легко обрабатывать большие потоки данных.
В отличие от аналоговых схем, наиболее полезные цифровые схемы являются синхронными, что означает наличие эталонных часов для координации работы схемных блоков, поэтому они работают предсказуемым образом. Аналоговая электроника работает асинхронно, то есть обрабатывает сигнал по мере его поступления на вход.
Большинство цифровых схем используют цифровой процессор для обработки данных. Это может быть простой микроконтроллер (MCU) или более сложный процессор цифровых сигналов (DSP), который может фильтровать и обрабатывать большие потоки данных, например видео.
Цифровые сигналы обычно используются в системах связи, где цифровая передача может передавать данные по каналам передачи «точка-точка» или «точка-многоточка», таким как медные провода, оптические волокна, средства беспроводной связи, носители данных или компьютерные шины. Передаваемые данные представлены в виде электромагнитного сигнала, например микроволнового, радиоволны, электрического напряжения или инфракрасного сигнала.
В общем, цифровые схемы легче спроектировать, но они часто стоят дороже, чем аналоговые схемы, предназначенные для тех же задач.
Каталог цифровых компонентов
MPS включает MP2886A, цифровой многофазный ШИМ-контроллер с интерфейсом PWM-VID, совместимый со спецификацией NVIDIA Open VReg.
Преобразование аналогово-цифрового (АЦП) и цифро-аналогового (ЦАП) сигнала
Многие системы должны обрабатывать как аналоговые, так и цифровые сигналы. Во многих системах связи обычно используется аналоговый сигнал, который действует как интерфейс для среды передачи для передачи и приема информации.Эти аналоговые сигналы преобразуются в цифровые, которые фильтруют, обрабатывают и сохраняют информацию.
Рисунок 5 показывает общую архитектуру, в которой аналоговый РЧ-интерфейс (AFE) состоит из всех аналоговых блоков для усиления, фильтрации и усиления аналогового сигнала. Между тем, секция процессора цифровых сигналов (DSP) фильтрует и обрабатывает информацию. Для преобразования сигналов из аналоговой подсистемы в цифровую подсистему в тракте приема (RX) используется аналого-цифровой преобразователь (ADC).Для преобразования сигналов из цифровой подсистемы в аналоговую подсистему в тракте передачи (TX) используется цифро-аналоговый преобразователь (DAC).
Рисунок 5: Система связи с аналоговыми и цифровыми подсистемами
Цифровой сигнальный процессор (DSP) — это специализированный микропроцессорный чип, который выполняет операции цифровой обработки сигналов. DSP изготавливаются на интегральных схемах MOSFET и широко используются в обработке аудиосигналов, телекоммуникациях, цифровой обработке изображений, телевизионных продуктах высокой четкости, обычных бытовых электронных устройствах, таких как мобильные телефоны, и во многих других важных приложениях.
DSP используется для измерения, фильтрации или сжатия непрерывных реальных аналоговых сигналов. Выделенные DSP часто имеют более высокую энергоэффективность, что делает их пригодными в портативных устройствах из-за ограничений по энергопотреблению. Большинство микропроцессоров общего назначения также могут выполнять алгоритмы цифровой обработки сигналов.
Работа АЦП
Рисунок 6 показывает работу АЦП. На вход поступает аналоговый сигнал, который обрабатывается схемой удержания выборки (S / H) для создания приближенного цифрового представления сигнала.Амплитуда больше не имеет бесконечных значений и была «квантована» до дискретных значений в зависимости от разрешения АЦП. АЦП с более высоким разрешением будет иметь более мелкие размеры шага и более точно представлять входной аналоговый сигнал. Последний каскад АЦП кодирует оцифрованный сигнал в двоичный поток битов, который представляет амплитуду аналогового сигнала. Цифровой вывод теперь можно обрабатывать в цифровом формате.
Рисунок 6: Типичная архитектура АЦП для преобразования аналогового сигнала в цифровой
Работа ЦАП
ЦАП обеспечивает обратную работу.Вход DAC представляет собой двоичный поток данных из цифровой подсистемы и выводит дискретное значение, которое аппроксимируется как аналоговый сигнал. По мере увеличения разрешения ЦАП выходной сигнал все больше приближается к истинному плавному и непрерывному аналоговому сигналу (см. Рисунок 7). Обычно в цепи аналогового сигнала есть постфильтр для дальнейшего сглаживания формы волны.
Рисунок 7: 6-битный ЦАП для цифро-аналогового преобразования сигнала
Как упоминалось ранее, многие системы, используемые сегодня, представляют собой «смешанные сигналы», что означает, что они полагаются как на аналоговые, так и на цифровые подсистемы.Эти решения требуют, чтобы АЦП и ЦАП преобразовывали информацию между двумя доменами.
Сравнение цифровых сигналов и аналоговых сигналов: преимущества и недостатки
Как и в большинстве инженерных тем, у аналоговых и цифровых сигналов есть свои плюсы и минусы. Конкретное приложение, требования к производительности, среда передачи и операционная среда могут определять, следует ли использовать аналоговую или цифровую сигнализацию (или их комбинацию).
Цифровые сигналы: преимущества и недостатки
Преимущества использования цифровых сигналов, включая цифровую обработку сигналов (DSP) и системы связи, включают следующее:
- Цифровые сигналы могут передавать информацию с меньшим шумом, искажениями и помехами.
- Цифровые схемы можно легко воспроизводить в массовых количествах при сравнительно небольших затратах.
- Цифровая обработка сигналов более гибкая, поскольку операции DSP можно изменять с помощью систем с цифровым программированием.
- Цифровая обработка сигналов более безопасна, поскольку цифровую информацию можно легко зашифровать и сжать.
- Цифровые системы более точны, и вероятность появления ошибок можно снизить за счет использования кодов обнаружения и исправления ошибок.
- Цифровые сигналы могут быть легко сохранены на любых магнитных или оптических носителях с использованием полупроводниковых микросхем.
- Цифровые сигналы могут передаваться на большие расстояния.
Недостатки использования цифровых сигналов, включая системы цифровой обработки сигналов (DSP) и системы связи, включают следующее:
- Для цифровой связи требуется более широкая полоса пропускания по сравнению с аналоговой передачей той же информации.
- DSP обрабатывает сигнал на высоких скоростях и содержит больше внутренних аппаратных ресурсов.Это приводит к более высокому рассеянию мощности по сравнению с обработкой аналогового сигнала, которая включает пассивные компоненты, потребляющие меньше энергии.
- Цифровые системы и обработка обычно более сложные.
Аналоговые сигналы: преимущества и недостатки
Преимущества использования аналоговых сигналов, включая системы обработки аналоговых сигналов (ASP) и связи, включают следующее:
- Аналоговые сигналы легче обрабатывать.
- Аналоговые сигналы лучше всего подходят для передачи аудио и видео.
- Аналоговые сигналы имеют гораздо более высокую плотность и могут предоставлять более точную информацию.
- Аналоговые сигналы используют меньшую полосу пропускания, чем цифровые сигналы.
- Аналоговые сигналы обеспечивают более точное представление изменений физических явлений, таких как звук, свет, температура, положение или давление.
- Аналоговые системы связи менее чувствительны с точки зрения электрических допусков.
Недостатки использования аналоговых сигналов, включая системы обработки аналоговых сигналов (ASP) и системы связи, включают следующее:
- Передача данных на большие расстояния может привести к нежелательным помехам сигнала.
- Аналоговые сигналы склонны к потере генерации.
- Аналоговые сигналы подвержены шумам и искажениям, в отличие от цифровых сигналов, которые имеют гораздо более высокую помехоустойчивость.
- Аналоговые сигналы обычно имеют более низкое качество, чем цифровые сигналы.
Аналоговые и цифровые сигналы: системы и приложения
В традиционных аудиосистемах и системах связи используются аналоговые сигналы. Однако с развитием технологий обработки кремния, возможностей цифровой обработки сигналов, алгоритмов кодирования и требований к шифрованию — в дополнение к увеличению эффективности использования полосы пропускания — многие из этих систем стали цифровыми.Они по-прежнему являются некоторыми приложениями, в которых аналоговые сигналы имеют устаревшее использование или преимущества. Большинство систем, которые взаимодействуют с реальными сигналами (такими как звук, свет, температура и давление), используют аналоговый интерфейс для захвата или передачи информации. Несколько приложений аналогового сигнала перечислены ниже:
- Запись и воспроизведение звука
- Датчики температуры
- Датчики изображения
- Радиосигналы
- Телефоны
- Системы управления
MPS предлагает широкий ассортимент аналоговых компонентов, включая MP2322, MP8714, MP2145 и MP8712.
Хотя многие оригинальные системы связи использовали аналоговую сигнализацию (телефоны), современные технологии используют цифровые сигналы из-за их преимуществ в отношении помехоустойчивости, шифрования, эффективности использования полосы пропускания и возможности использования ретрансляторов для передачи на большие расстояния. Несколько приложений цифрового сигнала перечислены ниже:
- Системы связи (широкополосная, сотовая)
- Сеть и передача данных
- Цифровые интерфейсы для программирования
Посетите наш веб-сайт, чтобы узнать больше о цифровых компонентах MPS, таких как MP2886A, MP8847, MP8868, MP8869S и MP5416.
Заключение
В этой статье представлены некоторые основные концепции аналоговых и цифровых сигналов и их использование в электронике. У каждой технологии есть явные преимущества и недостатки, и знание потребностей вашего приложения и требований к производительности поможет вам определить, какой сигнал (ы) выбрать.
_________________________
Вам это показалось интересным? Получайте ценные ресурсы прямо на свой почтовый ящик — рассылайте их раз в месяц!
Получить техническую поддержку
.