
Зависимость сопротивления от температуры
Опыт в соответствии с общими соображениями § 46 показывает, что сопротивление проводника зависит также и от его температуры.
Намотаем в виде спирали несколько метров тонкой (диаметра 0,1-0,2 мм) железной проволоки 1 и включим ее в цепь, содержащую батарею гальванических элементов 2 и амперметр 3 (рис. 81). Сопротивление этой проволоки подберем таким, чтобы при комнатной температуре стрелка амперметра отклонялась почти на всю шкалу. Отметив показания амперметра, сильно нагреем проволоку при помощи горелки. Мы увидим, что по мере нагревания ток в цепи уменьшается, а значит, сопротивление проволоки при нагревании увеличивается. Такой результат получается не только с железом, но и со всеми другими металлами. При повышении температуры сопротивление металлов увеличивается. У некоторых металлов это увеличение значительно: у чистых металлов при нагревании на 100°С оно достигает 40-50%; у сплавов оно обычно бывает меньше. Есть специальные сплавы, у которых сопротивление почти не меняется при повышении температуры; таковы, например, константан (от латинского слова constans – постоянный) и манганин.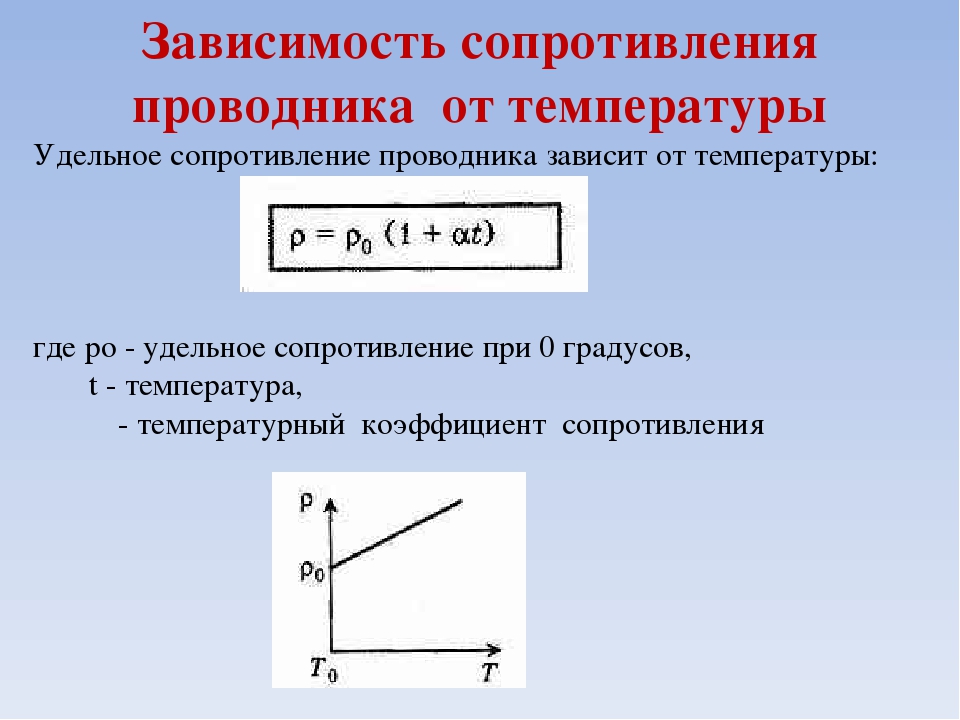 Константан употребляется для изготовления некоторых измерительные приборов.
Константан употребляется для изготовления некоторых измерительные приборов.
Рис. 81. Опыт, показывающий зависимость сопротивления проволоки от температуры. При нагревании сопротивление проволоки увеличивается: 1 – проволока, 2 – батарея гальванических элементов, 3 – амперметр
Иначе меняется при нагревании сопротивление электролитов. Повторим описанный опыт, но введем в цепь вместо железной проволоки какой-нибудь электролит (рис. 82). Мы увидим, что показания амперметра при нагревании электролита все время увеличиваются, а значит, сопротивление электролитов при повышении температуры уменьшается. Отметим, что сопротивление угля и некоторых других материалов также уменьшается при нагревании.
Рис. 82. Опыт, показывающий зависимость сопротивления электролита от температуры. При нагревании сопротивление электролита уменьшается: 1 – электролит, 2 – батарея гальванических элементов, 3 – амперметр
Зависимость сопротивления металлов от температуры используется для устройства термометров сопротивления. В простейшем виде это – намотанная на слюдяную пластинку тонкая платиновая проволока (рис. 83), сопротивление которой при различных температурах хорошо известно. Термометр сопротивления помещают внутрь тела, температуру которого желают измерить (например, в печь), а концы обмотки включают в цепь. Измеряя сопротивление обмотки, можно определить температуру. Такие термометры часто применяются для измерения очень высоких и очень низких температур, при которых ртутные термометры уже неприменимы.
В простейшем виде это – намотанная на слюдяную пластинку тонкая платиновая проволока (рис. 83), сопротивление которой при различных температурах хорошо известно. Термометр сопротивления помещают внутрь тела, температуру которого желают измерить (например, в печь), а концы обмотки включают в цепь. Измеряя сопротивление обмотки, можно определить температуру. Такие термометры часто применяются для измерения очень высоких и очень низких температур, при которых ртутные термометры уже неприменимы.
Рис. 83. Термометр сопротивления
Приращение сопротивления проводника при его нагревании на 1°С, разделенное на первоначальное сопротивление, называется температурным коэффициентом сопротивления и обычно обозначается буквой
. Вообще говоря, температурный коэффициент сопротивления сам зависит от температуры. Величина
имеет одно значение, например, если мы будем повышать температуру от 20 до 21°С, и другое при повышении температуры от 200 до 201°С. Но во многих случаях изменение
Но во многих случаях изменение
в довольно широком интервале температур незначительно, и можно пользоваться средним значением
в этом интервале. Если сопротивление проводника при температуре
равно
, а при температуре
равно
, то среднее значение
. (48.1)
Обычно в качестве
принимают сопротивление при температуре 0°С.
Таблица 3. Среднее значение температурного коэффициента сопротивления некоторых проводников (в интервале от 0 до 100 °С)
| Вещество | Вещество | ||
| Железо | 6,6 | Ртуть | 0,88 |
| Вольфрам | 4,8 | Никелин | 0,30 |
| Медь | 4,3 | Нихром | 0,13 |
| Серебро | 4,1 | Константан | 0,04 |
| Платина | 3,9 | Манганин | 0,02 |
В табл.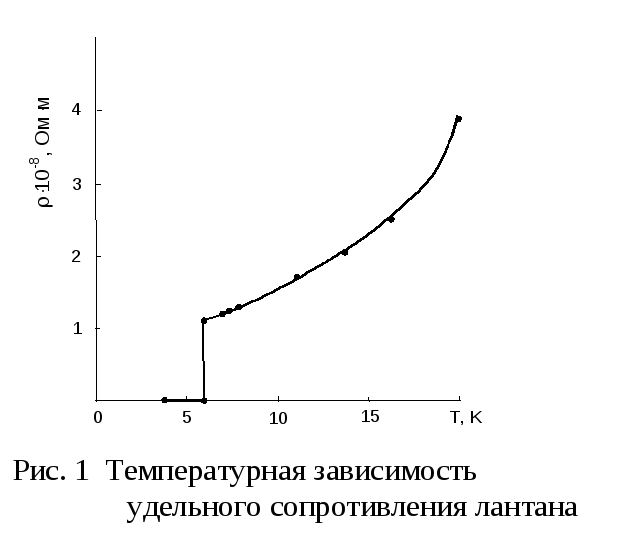 3 приведены значения
3 приведены значения
для некоторых проводников.
48.1.
При включении электрической лампочки сила тока в цепи в первый момент отличается от силы тока, который течет после того, как лампочка начнет светиться. Как изменяется ток в цепи с угольной лампочкой и лампочкой, имеющей металлическую нить накаливания?
48.2.
Сопротивление выключенной электрической лампочки накаливания с вольфрамовой нитью равно 60 Ом. При полном накале сопротивление лампочки возрастает до 636 Ом. Какова температура накаленной нити? Воспользуйтесь табл. 3.
48.3.
Сопротивление электрической печи с никелиновой обмоткой в ненагретом состоянии равно 10 Ом. Каково будет сопротивление этой печи, когда обмотка ее нагреется до 700°С? Воспользуйтесь табл. 3.
Зависимость сопротивления проводника от температуры.
Если
пропустить ток от аккумулятора через
стальную спираль, то амперметр покажет
уменьшение силы тока. Это означает, что
Это означает, что
с сопротивлением температуры сопротивление
проводника меняется.
Если
при температуре, равной 0С,
сопротивление проводника равно R0,
а при температуре t оно равно R, то
относительное изменение сопротивления,
как показывает опыт, прямо пропорционально
изменению температуры t:
(1)
Коэффициент
называетсятемпературным
коэффициентом сопротивления.
Он характеризует зависимость сопротивления
вещества от температуры.
Температурный
коэффициент сопротивления численно
равен относительному изменению
сопротивления проводника при повышении
температура на 1 К.
Для
всех металлов
>0
и незначительно меняется с изменением
температуры. У растворов электролитов
сопротивление с ростом температуры не
уменьшается, а увеличивается. Для них<0.
При
нагревании проводника его геометрические
размеры меняются незначительно.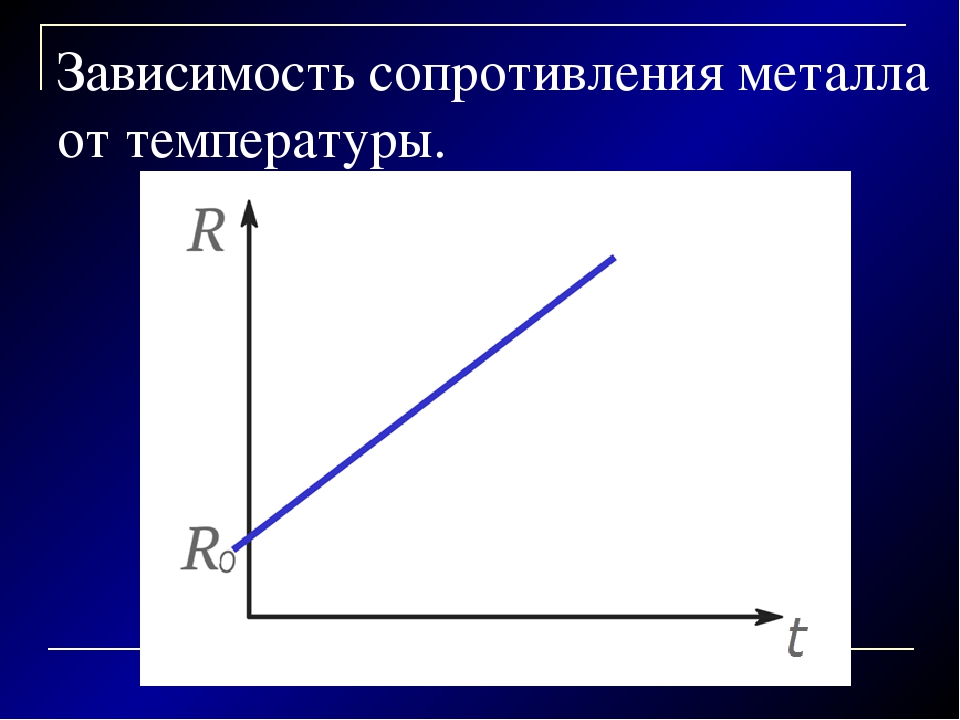
Сопротивление проводника меняется в
основном за счет изменения его удельного
сопротивления. Можно найти зависимость
этого удельного сопротивления от
температуры, если в формулу (1) подставить
значения
и
:
Так
как
мало меняется при изменении температуры,
то можно считать, что удельное
сопротивление проводника линейно
зависит от температуры.
С
приближением температуры к абсолютному
нулю удельное сопротивление монокристаллов
становится очень малым. Этот факт
свидетельствует о том, что в идеальной
кристаллической решетке металла
электроны перемещаются под действием
электрического поля, не взаимодействуя
с ионами решетки. Электроны взаимодействуют
лишь с ионами, не находящимися в узлах
кристаллической решетки.
При
повышении температуры возрастает число
дефектов кристаллической решетки из-за
тепловых колебаний ионов, – и это
приводит к возрастанию удельного
сопротивления кристалла.
Сверхпроводимость
В
1911 г. нидерландский ученый
Камерлинг-Оннес обнаружил, что при
понижении температуры ртути до 4,1 К
ее удельное сопротивление скачком
уменьшается до нуля. Явление уменьшения
удельного сопротивления до нуля при
температуре, отличной от абсолютного
нуля, называется сверхпроводимостью.
Материалы, обнаруживающие способность
переходить при некоторых температурах,
отличных от абсолютного нуля, в
сверхпроводящее состояние, называются
сверхпроводниками.
Прохождение тока
в сверхпроводнике происходит без потерь
энергии, поэтому однажды возбужденный
в сверхпроводящем кольце электрический
ток может существовать неограниченно
долго без изменения.
Сверхпроводящие
вещества уже используются в электромагнитах.
Однако получить сколь угодно сильное
магнитное поле с помощью сверхпроводящего
магнита нельзя, т. к. очень сильное
магнитное поле разрушает сверхпроводящее
состояние. Поэтому для каждого проводника
в сверхпроводящем состоянии существует
критическое значение силы тока, превзойти
которое, не нарушая сверхпроводящего
состояния, нельзя.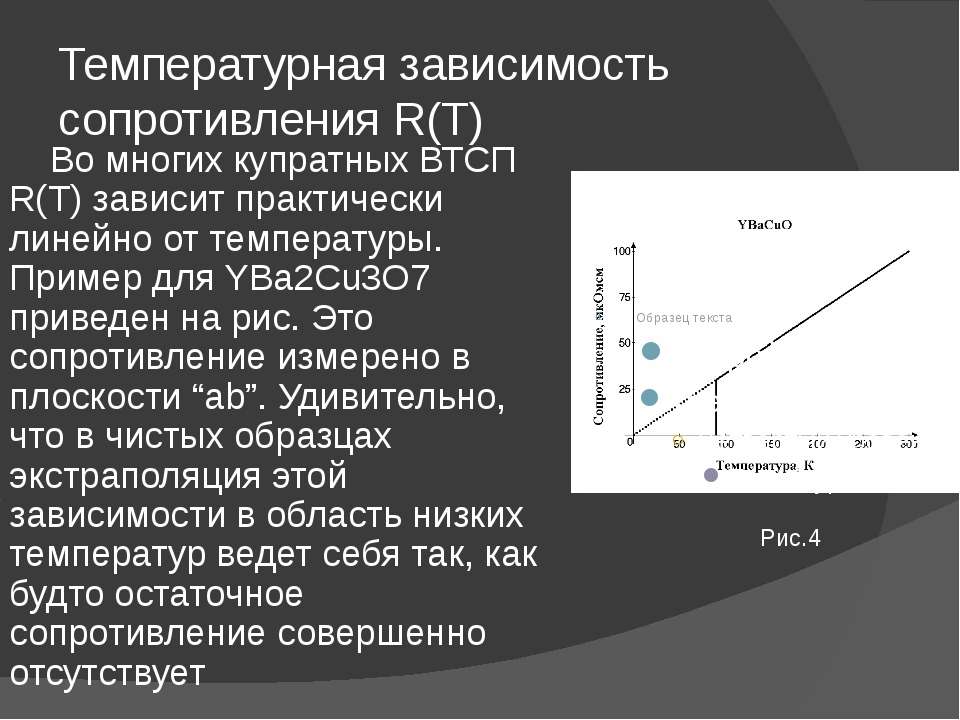
Объяснение
сверхпроводимости возможно только на
основе квантовой теории. Оно было дано
лишь в 1957 г.
В
1986 г. была открыта высокотемпературная
сверхпроводимость керамик – соединений
лантана, бария, меди и кислорода.
Сверхпроводимость таких керамик
сохраняется до температур около 100 К.
4. Зависимость сопротивления металлов и полупроводников от температуры
Для
характеристики температурной зависимости
сопротивления проводников вводится
температурный коэффициент сопротивления
,
который по определению равен:
(5)
Температурный
коэффициент сопротивления металлов —
это число, которое показывает, на
сколько изменится каждая единица
сопротивления проводника при изменении
температуры на 1°С (от 00С)
(6)
где
R0
— сопротивление данного проводника при
00
С; R
— сопротивление этого проводника при
t°С.
так
как
R0
неизвестно,
обычно вычисляют по .двум сопротивлениям:
R1=R0
(1+t1),
R2=R0()
откуда
(7)
Для
металлов
очень слабо зависит от температуры, но
для полупроводников дело обстоит иначе.
Электрическое
сопротивление полупроводников можно
выразить следующим образом:
(8)
где
— удельное сопротивление,- длина иS
– сечение полупроводника, А =.
Обозначив= В, получим
(9)
где
А — константа, пропорциональная «холодному»
сопротивлению полупроводника (обычно
при 20Со).
Постоянная
В является одной из важнейших характеристик
полупроводника, так как она определяет
его коэффициент сопротивления
.
Действительно
из выражений (5) и (9) находим:
(10)
Из
формулы (10) следует существенная
зависимость -а у полупроводников от
температуры.
Рис.6. Зависимости
удельных сопротивлений от температуры
для: а) металлов, б) диэлектриков,
в) полупроводников.
В
данной работе сравниваются температурные
зависимости сопротивления металлов
и полупроводников, вычисляются ,
температурный коэффициент сопротивления
металлов и ширина запрещенной зоны
полупроводника.
Ширина
запрещенной зоны ∆Е можно определить,
измерив экспериментально сопротивление
полупроводника при различных
температурах. Для этого приведем формулу
(8) к виду:
ℓgR=ℓgA+
(11)
где множители 103
введены для удобства дальнейших
вычислений. Эта зависимость в координатах
ℓgR,
при ΔЕ =const
представляет собой уравнение прямой,
тангенс угла наклона которой выражается
равенством
.-Отсюда
следует, что для определения 1 величины
ΔЕ графическим методом нужно
экспериментально измеренную зависимость
сопротивления полупроводника от
температуры пересчитать в зависимость:
(12)
и,
отложив по оси абсцисс
,
а по оси ординат,
определить тангенс угла наклона линейного
участка полученного графика и вычислить
значениепо формуле:
(13)
5. Описание установки
Описание установки
 Описание установки
Описание установки Внешний
вид установки представлен на рис.7.
Исследуемые образцы — медная проволока
и полупроводник (термосопротивление
ММТ-4) помещены в нагреватель, представляющий
собой проволочное остеклованное
сопротивление. Температура образцов
определяется ртутным термометром 2 с
пределом измерения от 0° до 150°С. В
зависимости от положения переключателя
(Rп
– Rм)
можно подсоединять к измерителю
сопротивления по желанию
полупроводниковый Rn
или металлический образец RM
. Нагреватель включается в сеть 220В
тумблером «Вкл» 3. Сопротивления
образцов с точностью ±1% измеряются с
помощью цифрового комбинированного
прибора ВК7-10А. Показания прибора в КΩ
непосредственно отсчитываются с
цифрового табло.
Рис.6.
Зависимость сопротивления от температуры
-
Автомобили
-
Бизнес
-
Дом и семья
-
Домашний уют
-
Духовное развитие
-
Еда и напитки
-
Закон
-
Здоровье
-
Интернет
-
Искусство и развлечения
-
Карьера
-
Компьютеры
-
Красота
-
Маркетинг
-
Мода
-
Новости и общество
-
Образование
-
Отношения
-
Публикации и написание статей
-
Путешествия
-
Реклама
-
Самосовершенствование
-
Спорт и Фитнес
-
Технологии
-
Финансы
-
Хобби
Рубрики
-
О проекте -
Реклама на сайте -
Условия -
Конфиденциальность -
Вопросы и ответы
FB
Войти
Немецкая овчарка Мэйджор станет первой собакой из приюта в Белом Доме Фланелевая рубашка помогла украсить дно старого деревянного сундучка
Температурное изменение сопротивления | Электрический ток, сопротивление и закон Ома
Температурное изменение сопротивления
Удельное сопротивление всех материалов зависит от температуры. Некоторые даже становятся сверхпроводниками (нулевое сопротивление) при очень низких температурах. (См. Этот рисунок.) И наоборот, удельное сопротивление проводников увеличивается с увеличением температуры. Поскольку атомы колеблются быстрее и на больших расстояниях при более высоких температурах, электроны, движущиеся через металл, совершают больше столкновений, эффективно увеличивая удельное сопротивление.При относительно небольших изменениях температуры (примерно \ (\ text {100º} \ text {C} \) или меньше) удельное сопротивление \ (\ rho \) изменяется с изменением температуры \ (\ Delta T \), как выражается в следующем уравнении
Некоторые даже становятся сверхпроводниками (нулевое сопротивление) при очень низких температурах. (См. Этот рисунок.) И наоборот, удельное сопротивление проводников увеличивается с увеличением температуры. Поскольку атомы колеблются быстрее и на больших расстояниях при более высоких температурах, электроны, движущиеся через металл, совершают больше столкновений, эффективно увеличивая удельное сопротивление.При относительно небольших изменениях температуры (примерно \ (\ text {100º} \ text {C} \) или меньше) удельное сопротивление \ (\ rho \) изменяется с изменением температуры \ (\ Delta T \), как выражается в следующем уравнении
\ (\ rho = {\ rho} _ {0} (\ text {1} + \ alpha \ Delta T) \ text {,} \)
где \ ({\ rho} _ {0} \) — исходное удельное сопротивление, а \ (\ alpha \) — температурный коэффициент удельного сопротивления . (См. Значения \ (\ alpha \) в этой таблице ниже.) Для более значительных изменений температуры \ (\ alpha \) может измениться, или может потребоваться нелинейное уравнение для нахождения \ (\ rho \). Обратите внимание, что \ (\ alpha \) положительно для металлов, что означает, что их удельное сопротивление увеличивается с температурой. Некоторые сплавы были разработаны специально, чтобы иметь небольшую температурную зависимость. Например, манганин (который состоит из меди, марганца и никеля) имеет значение \ (\ alpha \), близкое к нулю (к трем цифрам на шкале в этой таблице), поэтому его удельное сопротивление незначительно изменяется в зависимости от температуры. Это полезно, например, для создания не зависящего от температуры эталона сопротивления.
Обратите внимание, что \ (\ alpha \) положительно для металлов, что означает, что их удельное сопротивление увеличивается с температурой. Некоторые сплавы были разработаны специально, чтобы иметь небольшую температурную зависимость. Например, манганин (который состоит из меди, марганца и никеля) имеет значение \ (\ alpha \), близкое к нулю (к трем цифрам на шкале в этой таблице), поэтому его удельное сопротивление незначительно изменяется в зависимости от температуры. Это полезно, например, для создания не зависящего от температуры эталона сопротивления.
Сопротивление образца ртути равно нулю при очень низких температурах — это сверхпроводник примерно до 4.{-3} \)
Обратите внимание, что \ (\ alpha \) отрицателен для полупроводников, перечисленных в этой таблице, что означает, что их удельное сопротивление уменьшается с увеличением температуры. Они становятся лучшими проводниками при более высоких температурах, потому что повышенное тепловое перемешивание увеличивает количество свободных зарядов, доступных для переноса тока.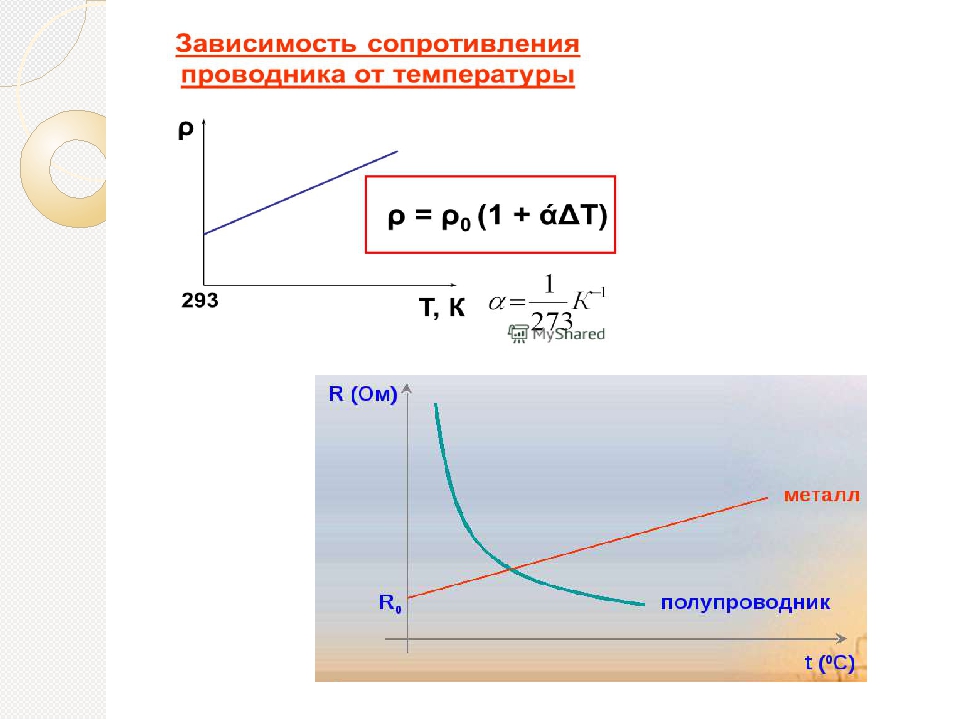 Это свойство уменьшения \ (\ rho \) с температурой также связано с типом и количеством примесей, присутствующих в полупроводниках.
Это свойство уменьшения \ (\ rho \) с температурой также связано с типом и количеством примесей, присутствующих в полупроводниках.
Сопротивление объекта также зависит от температуры, поскольку \ ({R} _ {0} \) прямо пропорционально \ (\ rho \).Для цилиндра мы знаем \ (R = \ mathrm {\ rho L} / A \), и поэтому, если \ (L \) и \ (A \) не сильно изменяются с температурой, \ (R \) будет иметь та же температурная зависимость, что и \ (\ rho \). (Исследование коэффициентов линейного расширения показывает, что они примерно на два порядка меньше типичных температурных коэффициентов удельного сопротивления, и поэтому влияние температуры на \ (L \) и \ (A \) примерно на два порядка меньше чем на \ (\ rho \).) Таким образом,
\ (R = {R} _ {0} (\ text {1} + \ alpha \ Delta T) \)
— это температурная зависимость сопротивления объект, где \ ({R} _ {0} \) — исходное сопротивление, а \ (R \) — сопротивление после изменения температуры \ (\ Delta T \).Многие термометры основаны на влиянии температуры на сопротивление. (См. Этот рисунок.) Одним из наиболее распространенных является термистор, полупроводниковый кристалл с сильной температурной зависимостью, сопротивление которого измеряется для определения его температуры. Устройство небольшое, поэтому быстро приходит в тепловое равновесие с той частью человека, к которой прикасается.
(См. Этот рисунок.) Одним из наиболее распространенных является термистор, полупроводниковый кристалл с сильной температурной зависимостью, сопротивление которого измеряется для определения его температуры. Устройство небольшое, поэтому быстро приходит в тепловое равновесие с той частью человека, к которой прикасается.
Эти знакомые термометры основаны на автоматическом измерении сопротивления термистора в зависимости от температуры.(кредит: Biol, Wikimedia Commons)
Пример: Расчет сопротивления: сопротивление горячей нити
Хотя при применении \ (\ rho = {\ rho} _ {0} (\ text {1} + \ alpha \ Delta T) \) и \ (R = {R} _ {0} (\ text {1} + \ alpha \ Delta T) \) для изменений температуры больше, чем \ (\ text {100º} \ text {C}. \) для вольфрама уравнения достаточно хорошо работают при очень больших изменениях температуры. Каково же тогда сопротивление вольфрамовой нити в предыдущем примере, если ее температура повышается с комнатной (C \ (\ text {20ºC} \)) до типичной рабочей температуры \ (\ text {2850º} \ text {C} \)?
Стратегия
Это прямое применение \ (R = {R} _ {0} (\ text {1} + \ alpha \ Delta T) \), поскольку исходное сопротивление нити накала было задано быть \ ({R} _ {0} = 0 \ text {. {–3} / \ text {ºC}) (\ text {2830º} \ text {C})] \\ & = & \ text {4.8 Ω.} \ End {array} \)
{–3} / \ text {ºC}) (\ text {2830º} \ text {C})] \\ & = & \ text {4.8 Ω.} \ End {array} \)
Обсуждение
Это значение согласуется с примером сопротивления фары в Законе Ома: сопротивление и простые цепи.
Исследования PhET: сопротивление в проводе
Узнайте о физике сопротивления в проводе. Измените его удельное сопротивление, длину и площадь, чтобы увидеть, как они влияют на сопротивление провода. Размеры символов в уравнении меняются вместе со схемой провода.
Изменение температуры и сопротивление | IOPSpark
Температурная зависимость сопротивления
Электричество и магнетизм
Изменение температуры и сопротивление
Практическая деятельность
для 14-16
Практический класс
Исследование изменения сопротивления провода при нагревании.
Аппаратура и материалы
- Источник питания, низкое напряжение, постоянный ток, например 12 В, 4 А не менее
- Реостат (10-20 Ом) на 5 А минимум
- Алюминиевый контейнер (например, одноразовый контейнер для пищевых продуктов)
- Амперметр, от 0 до 5 А, постоянный ток
- Вольтметр, от 0 до 10 В, постоянный ток
- Выводы, 4 мм, 6
- Зажимы типа Crocodile, 2
- Катушка медной проволоки (см. Техническое примечание)
Примечания по охране труда и технике безопасности
Проволока с полиуретановым покрытием может выделять опасные пары при перегреве.Окна лаборатории следует открывать, чтобы снизить риск.
Прочтите наше стандартное руководство по охране труда
Катушка должна быть свободно намотана из 1 м медной проволоки с полиуретановым покрытием (30 или 33 SWG). Снимите покрытие с концов, чтобы обеспечить электрический контакт через зажимы типа «крокодил».
Процедура
- Создайте простую последовательную цепь с длинными выводами к свободно намотанной катушке медного провода.
- Отрегулируйте источник питания, чтобы в катушке был ток около 4-5 ампер.Как можно скорее выключите цепь.
- Примерно через минуту змеевик остынет до комнатной температуры. Снова включите цепь. Снимите показания амперметра и вольтметра несколько раз в течение следующих полминуты или около того. За это время катушка нагревается, и ток меняется довольно быстро.
- Повторите эксперимент с катушкой из медной проволоки, подвешенной в воде в контейнере. Воду следует держать очень хорошо перемешанной. Будьте осторожны, чтобы избежать короткого замыкания змеевика, используя деревянную палочку на палочке или лопатку в качестве мешалки.
Учебные заметки
- Учащиеся записывают пары значений тока и разности потенциалов, когда катушка находится в воздухе, а затем строят график характеристики разности тока / разности потенциалов. Это не прямая линия, показывающая постоянное сопротивление, а скорее кривая, показывающая, что сопротивление провода увеличивается с температурой.
- Когда эксперимент повторяется с змеевиком в водяной бане, чтобы его температура оставалась постоянной, характеристический график представляет собой прямую линию, показывающую, что сопротивление остается постоянным.Чистые металлы подчиняются закону Ома, если их температура остается постоянной. Провода, изготовленные из сплавов, таких как проволока Constantan или Eureka (состоящая из 60% меди и 40% никеля), имеют очень малый температурный коэффициент удельного сопротивления. Поэтому их не нужно помещать в ванну с постоянной температурой, чтобы показать омическое поведение.
- Этот эксперимент может быть использован, чтобы научить понимать достоверность научных результатов. Результаты могут быть признаны недействительными, если на результаты влияет неконтролируемый фактор. В этом случае температура провода является фактором, влияющим на измерения его сопротивления.
- Обсудите, как это можно учесть. Один из подходов (как обсуждалось выше) состоит в том, чтобы поддерживать постоянную температуру проволоки, погружая ее в водяную баню. Альтернативой может быть использование приведенного выше графика p.d./current, чтобы найти сопротивление провода, когда через него не течет ток, потому что тогда нет эффекта нагрева. Объясните, что сопротивление провода равно p.d. делится на ток; я.е. это градиент линии от начала координат до точки на графике. Поместите линейку на график через начало координат и через самую высокую точку графика. Линейка имеет большой уклон. Двигайтесь вниз по графику от точки к точке, показывая, что градиент уменьшается. Ближе к началу координат график почти прямой (или вы можете использовать идею касательной к графику). Таким образом можно определить сопротивление провода, когда он не нагревается током.
- Этот эксперимент можно расширить, включив в него исследование влияния температуры на сопротивление в диапазоне от 0 ° C до 100 ° C с использованием водяной бани. Если студенты знакомы с экспериментальным наблюдением закона Чарльза, вы можете попросить их экстраполировать график зависимости сопротивления от температуры, чтобы найти приблизительную температуру, при которой сопротивление провода будет равно нулю. Для чистого металла сопротивление уменьшается примерно линейно в направлении температуры, близкой к 0 К. (Температурный коэффициент сопротивления многих чистых металлов близок к 0,004 K-1, поэтому график сопротивления / температуры будет экстраполирован обратно до 1 / 0,004 = 250. К.) Вы можете связать это с идеей о том, что сопротивление чистого металла при комнатной температуре зависит от вибрации ионов, и это снизится до нуля близко к 0 К.
 Это не прямая линия, показывающая постоянное сопротивление, а скорее кривая, показывающая, что сопротивление провода увеличивается с температурой.
Это не прямая линия, показывающая постоянное сопротивление, а скорее кривая, показывающая, что сопротивление провода увеличивается с температурой.Как работает наука? Добавочный номер:
 В этом случае температура провода является фактором, влияющим на измерения его сопротивления.
В этом случае температура провода является фактором, влияющим на измерения его сопротивления. Если студенты знакомы с экспериментальным наблюдением закона Чарльза, вы можете попросить их экстраполировать график зависимости сопротивления от температуры, чтобы найти приблизительную температуру, при которой сопротивление провода будет равно нулю. Для чистого металла сопротивление уменьшается примерно линейно в направлении температуры, близкой к 0 К. (Температурный коэффициент сопротивления многих чистых металлов близок к 0,004 K-1, поэтому график сопротивления / температуры будет экстраполирован обратно до 1 / 0,004 = 250. К.) Вы можете связать это с идеей о том, что сопротивление чистого металла при комнатной температуре зависит от вибрации ионов, и это снизится до нуля близко к 0 К.
Если студенты знакомы с экспериментальным наблюдением закона Чарльза, вы можете попросить их экстраполировать график зависимости сопротивления от температуры, чтобы найти приблизительную температуру, при которой сопротивление провода будет равно нулю. Для чистого металла сопротивление уменьшается примерно линейно в направлении температуры, близкой к 0 К. (Температурный коэффициент сопротивления многих чистых металлов близок к 0,004 K-1, поэтому график сопротивления / температуры будет экстраполирован обратно до 1 / 0,004 = 250. К.) Вы можете связать это с идеей о том, что сопротивление чистого металла при комнатной температуре зависит от вибрации ионов, и это снизится до нуля близко к 0 К.Этот эксперимент прошел испытания на безопасность в октябре 2006 г.
Недостатки резистивных датчиков температуры (RTD)
Датчик температуры сопротивления ( RTD ) — это датчики, используемые для измерения температуры. Многие элементы RTD состоят из отрезка тонкого провода, намотанного на керамический или стеклянный сердечник, но используются и другие конструкции.
Многие элементы RTD состоят из отрезка тонкого провода, намотанного на керамический или стеклянный сердечник, но используются и другие конструкции.
Провод RTD — это чистый материал, обычно платина, никель или медь.
Материал имеет точное соотношение сопротивления / температуры, которое используется для определения температуры. Поскольку элементы RTD хрупкие, их часто помещают в защитные зонды.
Ниже перечислены основные недостатки датчиков температуры сопротивления:
Самонагревание
Тепловая энергия генерируется при подаче тока для возбуждения элемента RTD для измерения его сигнала.
Возникающий самонагрев приведет к ошибке измерения температуры. Поскольку RTD изменяет свое сопротивление в зависимости от температуры, наиболее практичный способ его измерения — пропустить через него ток и измерить результирующее падение напряжения.
К сожалению, этот ток возбуждения, проходящий через сопротивление элемента, повышает температуру элемента, поскольку он пытается рассеять эту электрическую энергию за счет тепла, добавляя погрешность к нашему измерению температуры.
Способ борьбы с положительным сдвигом, вызванным самонагревом, заключается в увеличении теплового контакта с материалом, который мы ощущаем, и / или уменьшении тока возбуждения.
Самонагрев датчика RTD чаще всего выражается в мВт / ° C, что означает мощность, необходимую для повышения температуры внутреннего элемента на 1 ° C. Таким образом, чем выше этот показатель, тем меньше будет самонагрев.
Например, предположим, что ток возбуждения 2 мА используется для управления платиновым резистивным датчиком температуры 100 Ом при 100 ° C. Это дает сопротивление датчика 138,5 Ом. Его характеристики самонагрева составляют 50 мВт / ° C в воде, движущейся со скоростью 1 м / с.
Таким образом, количество тепла, выделяемого этой конфигурацией, составляет 1000 мВт / Вт * I 2 * R = 1000 * (0.002A) 2 * 138,5 Ом = 0,55 мВт.
Это приводит к ошибке самонагрева только (0,55 мВт) / (50 мВт / ° C) = 0,01 ° C.
Важно отметить, что эффективное самонагревание элемента сильно зависит от среды, в которую он погружен.
Например, RTD может самонагреваться в неподвижном воздухе в 100 раз сильнее, чем в движущейся воде, к которой применяются эти спецификации.
Поскольку мы измеряем сопротивление RTD, протягивая через него ток, мощность I 2 R, рассеиваемая RTD, вызывает самонагревание элемента.
Самонагрев приведет к изменению сопротивления RTD и увеличению погрешности измерения.
Негативный эффект самонагрева можно свести к минимуму, подавая меньший ток возбуждения.
Некоторые приборы используют токи возбуждения RTD до 0,1 мА, чтобы минимизировать эту ошибку.
В приведенном выше примере это снизит самонагревание до ~ 0,001 мВт / 50 мВт / ° C = 0,00003 ° C, незначительное количество даже в неподвижном воздухе.
Величина этой ошибки обратно пропорциональна способности чувствительного элемента рассеивать тепло.Это продукт материалов, конструкции и окружающей среды.
Элементы RTD с малым корпусом будут иметь более высокий эффект самонагрева, так как они имеют меньшую площадь поверхности для рассеивания тепла.
Возможно, наихудшим вариантом будет тонкопленочный RTD, который обычно будет иметь высокое тепловое сопротивление и соответствующую небольшую площадь поверхности для рассеивания тепла.
Обычно постоянная рассеяния указывается в технических характеристиках датчика RTD. Это число относится к мощности, необходимой для повышения температуры RTD на один градус температуры.
Таким образом, постоянная рассеяния 25 мВт / ° C показывает, что если потери мощности I 2 R в RTD равны 25 мВт, то RTD будет нагреваться на 1 ° C.
Константа рассеяния обычно указывается при двух условиях: свободный воздух и хорошо перемешиваемая масляная баня. Это происходит из-за разной способности среды отводить тепло от устройства.
Повышение температуры самонагрева можно определить по мощности, рассеиваемой RTD, и постоянной рассеиваемой энергии с помощью следующего:
ΔT = P / PD
где ΔT = повышение температуры из-за самонагрева в ° C; P = мощность, рассеиваемая в RTD из цепи в Вт; и PD = постоянная рассеяния RTD в Вт / ° C.
Итоги:
Ошибки самонагрева вызваны неспособностью элемента RTD отводить тепло, выделяемое требуемой мощностью, подаваемой через измерительный ток.
Стандарт ASTM требует, чтобы погрешность составляла максимум 1 ° C при подаче 33 мВт в воде 25 ° C, IEC требует максимальной погрешности 0,05 ° C в воде 25 ° C при приложении максимального рабочего тока.
Эти методы испытаний являются хорошими методами лабораторного сравнения.Для ПТС, установленных с надлежащим погружением в технологический процесс, рабочий ток составляет 1 мА или меньше, поэтому мощность (I 2 R) для ПТС на 100 Ом также мала (0.02–0,39 мВт).
Ошибки лагера могут возникать в датчиках с сопротивлением в диапазоне 500–1000 Ом, или когда процесс демонстрирует плохие условия теплопередачи, такие как неподвижный воздух или газы низкого давления.
Термоэдс или термоэлектрический эффект Зеебека
Возможно, вы думали, что эффект Зеебека применим только к термопарам? Но, как и термопары, платиновые RTD также построены с использованием двух разных металлов — платинового элемента RTD и меди выводных проводов.
Для некоторых приложений эти соединения в контуре датчика могут генерировать напряжения Зеебека, которые могут противодействовать падениям ИК-излучения, возникающим в резистивном элементе, и слегка искажать показания.
Например, если вдоль чувствительного элемента может развиваться температурный градиент, то термоэлектрическое напряжение приблизительно 7 мкВ / ° C может возникнуть в результате переходов между платиновым чувствительным элементом и медным подводящим проводом.
Для большинства приложений эта малая противоэдс не будет значительным источником ошибок, но может привести к проблемам в очень точных измерительных системах, работающих при низких токах возбуждения (возможно, для минимизации ошибок самонагрева) — условия обычно встречаются только в лабораторных измерениях.
Материал и конструкция RTD делают его относительно громоздким элементом, что также затрудняет использование RTD для измерения температуры в одной точке контакта.
Тем не менее, RTD обеспечивает отличное средство измерения средней температуры по поверхности, и он делает это, распределяя контакт резистивного провода по площади поверхности.
Но если этот поверхностный контакт также распространяется на некоторое расстояние, так что соединения выводных проводов на каждом конце элемента смещены слишком далеко друг от друга, то это может привести к ошибке Зеебека, которая является побочным продуктом температурного градиента, возникающего между два соединения платина-медь с выводами.
Эти ошибки можно предотвратить, используя соответствующий подводящий провод и осторожное позиционирование датчика относительно выводных проводов.
Короче говоря, другой свинцовый материал, такой как медь, может создавать Т / С переход, где он соединяется с платиновым элементом, а затем другой Т / С переход на другом конце.
Если два перехода находятся при разных температурах, то разовьется термоэлектрическая ЭДС, которая может нарушить ИК-измерения элемента RTD.
Алгебраическая сумма термоэлектрической ЭДС в цепи, состоящей из любого количества разнородных материалов, равна нулю, если все переходы поддерживаются при постоянной температуре.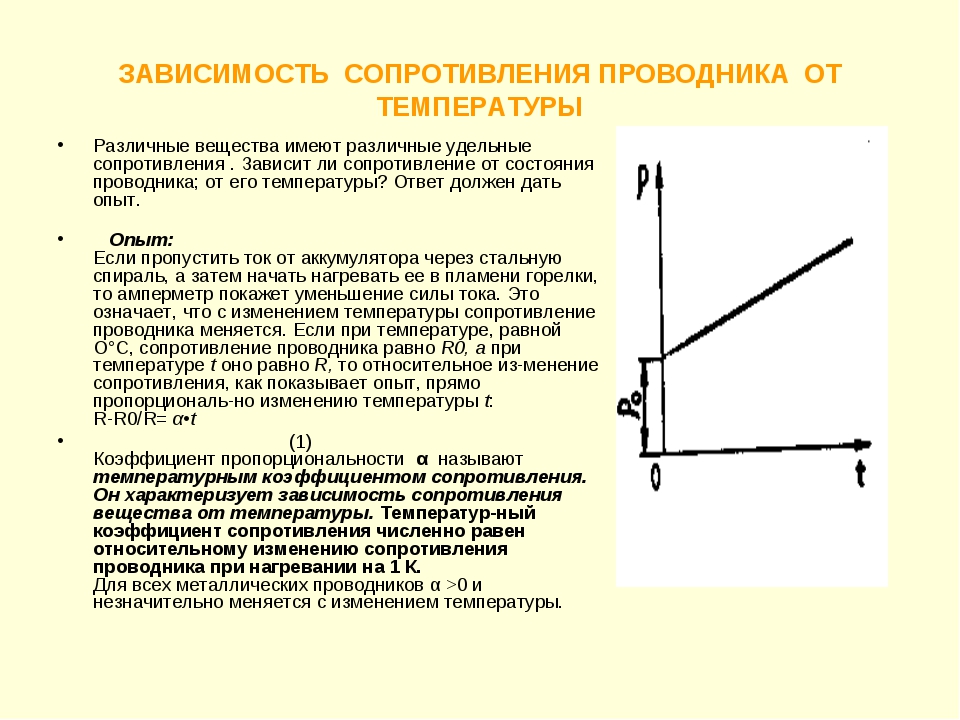
Таким образом, у вас есть только два средства для борьбы с этим эффектом: либо используйте выводной провод из того же материала, что и элемент (не практично, так как это было бы очень дорого для платинового элемента с длинными выводами), либо просто поддерживайте температуру на уровне все соединения одинаковы (т. е. вдоль элемента) или почти одинаковы, что приведет к незначительному вкладу чистой ЭДС в измерение напряжения.
Итоги:
Погрешности термо-ЭДС также известны как эффект термопары.Эта ошибка вызвана различным составом проводов, однородностью материалов соединений проводов и температурными градиентами внутри PRT (RTD).
Стандарты ASTM и IEC предлагают рекомендации для высоких чувствительных токов — хотя, когда существует влияние ЭДС, оно будет иметь больший эффект при более низких токах, близких к стандартному рабочему току.
Эта ошибка возникает в основном в системах постоянного тока. Чтобы свести к минимуму ошибки термо-ЭДС, выберите PRT с низкой указанной ЭДС.
Кроме того, использование схемы переменного тока и надлежащим образом выбранных передатчиков может устранить влияние ЭДС.
Время отклика или время отклика
Постоянная времени RTD — это скорость, с которой его элемент изменяет сопротивление в ответ на изменение температуры контакта.
Быстрая постоянная времени помогает уменьшить ошибку в системе измерения, которая обнаруживает быстрые изменения температуры.
Когда мы рассматриваем конструкцию RTD, мы можем сделать вывод, что время отклика будет сильно зависеть от массы чувствительного элемента и его изолирующей конструкции, а также от способности теплопередачи к измеряемому материалу.
Это напрямую влияет на скорость передачи тепла от внешней чувствительной поверхности к сердечнику чувствительного элемента.
Для сравнения, поскольку RTD измеряет температуру на большей площади, а не в небольшой точке контакта, такой как термопара, и поскольку чувствительный элемент RTD должен быть изолирован, он имеет гораздо более медленное время отклика, чем термопара.
Аналогичным образом, датчик RTD в защитной гильзе будет реагировать медленнее, чем тот же датчик, погруженный непосредственно в жидкость.
Датчик в прочно соединенном внутреннем узле будет реагировать в два раза быстрее, чем датчик с одним незакрепленным интерфейсом в том же узле.
Поверхностный RTD быстрее реагирует на изменение температуры поверхности.
Время отклика для данного датчика обычно определяется как время, за которое датчик достигает 63% своего конечного значения при тепловом равновесии в ответ на скачкообразное изменение температуры контакта.
Это время обычно выражается в измерении скорости потока воды 1 м / сек (3 фута / сек) и / или скорости потока воздуха 3 м / сек (10 футов / сек).
Хотя это встречается реже, иногда время отклика относится к временному интервалу, за который платиновый RTD достигает 90% от своего окончательного значения (в отличие от 63%).
Обязательно обратите внимание на это различие при сравнении типов датчиков.
Итоги:
Ошибка, связанная со временем отклика, может возникнуть во время температурных переходных процессов, если PRT (RTD) не может достаточно быстро реагировать на изменение температуры.
В установившемся или почти установившемся режиме эта ошибка равна нулю.ASTM и IEC не определяют эту ошибку, хотя существует метод тестирования для определения времени отклика PRT в целях сравнения.
Когда важно отслеживать переходные условия, эту ошибку можно свести к минимуму, выбрав датчик с более быстрым временем отклика, проверенным в лаборатории, и оценив соответствующую скорость изменения процесса, чтобы наилучшим образом соответствовать характеристикам времени отклика датчиков.
Статья Источник: Acromag
Температурная зависимость
Колебание μ в зависимости от температуры
Умеренная зависимость μ определяется двумя факторами; рассеяние фононов и рассеяние на ионизованных примесях. С повышением температуры тепловые колебания (фононы) внутри полупроводника увеличиваются и вызывают повышенное рассеяние. Это приводит к снижению подвижности носителей. Мы знаем, что
С повышением температуры тепловые колебания (фононы) внутри полупроводника увеличиваются и вызывают повышенное рассеяние. Это приводит к снижению подвижности носителей. Мы знаем, что
, где tau — среднее время свободного пробега между столкновениями. Среднюю длину свободного пробега можно записать как пропорциональную
Теперь рассмотрим, что длина свободного пробега обратно пропорциональна вероятности рассеяния, и что вероятность рассеяния можно считать пропорциональной энергии решетки (т.е.е. к Т). Это позволяет записать тау как
.
Таким образом, мы ожидаем, что это снижение мобильности будет пропорционально T -3/2 .
Однако эффект рассеяния ионизованной примеси уменьшается с повышением температуры из-за более высоких средних тепловых скоростей носителей. Носители проводят меньше времени рядом с ионизированной примесью по мере прохождения, и, таким образом, эффект рассеяния уменьшается. Это моделируется, предполагая, что никакого рассеяния не произойдет, если электрон не окажется достаточно близко к иону, чтобы электростатическая энергия была сопоставима с тепловой энергией.
, что дает радиус
.
Далее мы можем утверждать, что рассеивающая способность иона может быть представлена сечением рассеяния
.
Наконец, предполагая, что длина свободного пробега обратно пропорциональна сечению рассеяния, мы получаем соотношение
.
Следовательно, можно ожидать, что подвижность из-за рассеяния ионизованных примесей будет увеличиваться при повышении температуры согласно T +3/2 .
Эти два эффекта действуют одновременно, при этом общее время столкновения равно
.
Обычно рассеяние на ионизованных примесях преобладает при низких температурах, тогда как при более высоких температурах преобладает рассеяние на фононах. Экспериментальные значения температурной зависимости подвижности в Si, Ge и GaAs приведены в таблице ниже [1].
| Si | Ge | GaAs | |
Подвижность электронов | Т -2. 4 4 | т -1,7 | Т -1,0 |
| Отверстие Мобикиты | Т -2,2 | Т -2,3 | т -2,1 |
Влияние температуры на V H
Чтобы понять, как напряжение Холла изменяется с температурой, мы должны сначала рассмотреть проводимость измеряемого образца. Электропроводность полупроводника можно выразить следующим образом:
Обратите внимание, что как концентрации носителей (N e и N h ), так и подвижности зависят от температуры, хотя и имеют разные функциональные формы.
В диапазоне температур примесей, где T достаточно велико только для ионизации любых доноров в полупроводнике, концентрации носителей остаются постоянными, и единственное изменение сигма с температурой является результатом изменения подвижностей. Если предположить, что рассеяние фононов преобладает, мы знаем, что μ изменяется как T -3/2 . Таким образом, мы знаем σ ~ T -3/2 , и, учитывая, что σ = 1 / ρ и ρ∝R, мы можем определить, что R ~ T 3/2 .
Таким образом, мы знаем σ ~ T -3/2 , и, учитывая, что σ = 1 / ρ и ρ∝R, мы можем определить, что R ~ T 3/2 .
Как указывалось ранее, напряжение Холла можно записать как
, только ток зависит от температуры.Этот ток I можно записать в виде
, что означает, что I и, следовательно, V H изменяются в соответствии с T 3/2 . На самом деле эти колебания температуры довольно малы, и V H на практике можно считать постоянным во внешнем режиме.
В собственной области, в которой температура достаточно высока для возбуждения носителей заряда из валентной зоны в зону проводимости, концентрации носителей (N e и N h ) экспоненциально возрастают с ростом T.Это доминирует над любым воздействием подвижности на электрическую проводимость и приводит к быстрому уменьшению удельного сопротивления полупроводника с повышением температуры. Таким образом, напряжение Холла падает с увеличением T.
Определение энергетического разрыва
Изменение проводимости с температурой также позволяет нам измерить энергетический зазор между валентной зоной и зоной проводимости. И снова проводимость полупроводника
.
Что касается внутреннего материала, мы знаем, что
Объединение этих двух уравнений дает
Мы можем еще раз игнорировать изменение T 3/2 , которое будет в основном незначительным по сравнению с экспоненциальным изменением температуры.Таким образом, график зависимости ln (σ) от 1 / T будет иметь наклон –Eg / 2k, что дает значение для Eg.
В случае легированных полупроводников отметим, что при высоких температурах они являются внутренними по поведению и становятся псевдо-собственными при низких температурах, при этом энергетическая щель равна щели между примесным уровнем и краем зоны. Следовательно, на графике, подобном упомянутому выше, мы ожидаем двух прямолинейных участков с разными наклонами. В области между этими наклонами температура достаточно высока для полной ионизации доноров, но недостаточно высока для ионизации значительного числа электронов из решетки.Следовательно, в этом температурном диапазоне плотность носителей не будет сильно зависеть от температуры, а подвижность, которой ранее пренебрегли, будет определять форму кривой.
В области между этими наклонами температура достаточно высока для полной ионизации доноров, но недостаточно высока для ионизации значительного числа электронов из решетки.Следовательно, в этом температурном диапазоне плотность носителей не будет сильно зависеть от температуры, а подвижность, которой ранее пренебрегли, будет определять форму кривой.
Список литературы
- Bart van Zeghbroeck, Principles of Semiconductor Devices , Boulder, 2006
2.9 Подвижность — Удельное сопротивление — Сопротивление листа
2.9 Подвижность — Удельное сопротивление — Сопротивление листа
Содержание —
Глоссарий —
Учебные пособия —
В этой секции:
- Массовая мобильность
- Температурная зависимость подвижности
- Удельное сопротивление
- Листовое сопротивление
2.9.1 Массовая мобильность
Подвижность электронов и дырок в объемном кремнии равна
показано на рисунке ниже.
resistiv.xls — мобильность.gif
- Рис. 2.9.1 Зависимость подвижности электронов и дырок от
плотность легирования кремния
Это активная фигура, которую можно использовать для поиска
объемная подвижность для конкретных концентраций легирования, а также
связанные удельное сопротивление и сопротивление листа.
Обратите внимание, что мобильность связана с общим количеством человек.
ионизированные примеси или сумма донора и акцептора, а не плотность свободных носителей, которая
в первую очередь связано с разницей между донором
и концентрация акцептора.
Подвижность неосновных носителей также зависит от общей примеси
плотности, используя кривую, соответствующую неосновному носителю
тип.
Кривые рассчитаны из
эмпирическое выражение:
- (моб10)
где м мин ,
м макс ,
a и N r являются подходящими параметрами. Эти
параметры для легированных мышьяком, фосфором и бором
кремний
приведено в таблице ниже:
tmob1.gif
Пример 006
2.9.1.1 Температурная зависимость
мобильность
2.9.2 Удельное сопротивление
Проводимость материала определяется как
плотность тока, деленная на приложенное электрическое поле. поскольку
плотность тока равна произведению заряда
мобильные носители, их плотность и скорость могут быть
выражается как функция электрического поля с помощью
мобильность. Чтобы учесть также вклад электронов
в качестве дырок к проводимости добавляем плотность тока
из-за дырок в электроны, или:
- (моб8)
Проводимость за счет электронов и дырок равна
затем получено из:
- (моб9)
Удельное сопротивление определяется как величина, обратная величине
проводимость, а именно:
- (моб5)
Результирующее удельное сопротивление, рассчитанное по выражению
выше показано на рисунке ниже:
резистив.xls — resistiv.gif
- Рис. 2..9.2 Удельное сопротивление n-типа (красная кривая) и p-типа
(синяя кривая) кремний против
плотность легирования
Пример 003 —
Пример 004
2.9.3 Удельное сопротивление пластины толщиной 14 мил
Понятие сопротивления листа используется для характеристики обеих пластин как тонких.
легированные слои, поскольку обычно легче измерить сопротивление листа
а не удельное сопротивление материала.Листовое сопротивление
слой с сопротивлением, r , и
толщина, т , определяется их соотношением:
- (моб7)
Строго говоря, единицы сопротивления листа — Ом,
один относится к числу Ом на квадрат. Эта номенклатура
пригодится, когда сопротивление прямоугольного куска материала с
длина должна быть L и ширина W . Это
равно произведению
сопротивление листа и количество квадратов или:
- (моб6)
где количество квадратов равно длине, деленной на ширину.
На рисунке ниже показано сопротивление пластины кремниевой пластины толщиной 14 мил.
который является n-типом (синяя кривая) или p-типом (красная кривая)
resistiv.xls — sheetres.gif
- Рис. 2..9.3 Удельное сопротивление листа толщиной 14 мил.
n-тип (красная кривая) и p-тип
(синяя кривая) кремниевая пластина
плотность легирования. Эту активную цифру можно изменить на
подходит для любой толщины слоя.
2,8
2.10
Барт Дж.Ван Зегбрук, 1996, 1997
Frontiers | Температурная зависимость сна
Введение
У всех млекопитающих сон необходим и совпадает с сохраненным циркадным температурным ритмом. Когда наша внутренняя температура и температура мозга быстро снижаются, мы, скорее всего, предпочтем спать, а если мы откажемся от этого цикла охлаждения тела, мы испытаем бессонницу (Hayward, 1968; Campbell and Broughton, 1994; Lack et al., 2008). . Здесь мы рассматриваем доказательства того, что механизмы терморегуляции имеют фундаментальное значение для сна, и рассматриваем нейронные цепи, которые связывают эти две физиологии.Эти схемы используют теплый микроклимат для выхода из режима сна и могут улучшить циркадное охлаждение тела по мере приближения нашего первого боя с медленным движением глаз (NREM). Те же нейроны напрямую связывают инициирование NREM с охлаждением тела и могут объяснить, почему переходы от бодрствования к NREM-сну в течение всего цикла сна немедленно сопровождаются снижением температуры мозга, в то время как переходы обратно в REM или WAKE сопровождаются согреванием (Alföldi et al. др., 1990; Ландольт и др., 1995). Разделение охлаждения мозга во время медленного сна и координация суточного ритма основной температуры важны для эффективного сна.Это может иметь определенные последствия для энергетического гомеостаза и может открыть окно для функции сна.
Подготовка ко сну — это терморегуляторное поведение
Млекопитающие обладают рядом способов терморегуляции, которые позволяют адаптироваться к колебаниям температуры окружающей среды в течение дня, но они наиболее заметны при подготовке ко сну (Peever, 2018). Эти поведения включают в себя тепло и поиск убежища, строительство гнезда, свертывание калачиком и сбивчивание (см. Рисунок 1A).Мыши, которые неактивны или спят, гораздо чаще контактируют с материалом для гнездования (Gaskill et al., 2011). Как маленькие грызуны, они демонстрируют удивительно сложные тепловые приспособления. По мере снижения температуры окружающей среды качество гнезд повышается для компенсации и приводит к ощутимым улучшениям изоляции (Gaskill et al., 2013a). По возможности мыши сбиваются в кучу с членами группы (Gaskill et al., 2011, 2012; Gordon et al., 2014). У них также есть четкое тепловое предпочтение во время фазы сна (свет включен), выбирая более теплую среду, приближающуюся к термонейтральности (27–30 ° C), и сводя к минимуму расход энергии (Gordon et al., 1998; Gaskill et al., 2012). Такое поведение согласовывает снижение суточной температуры с циклом света и темноты и наступлением сна. Пример циркадного цикла внутренней температуры в течение нескольких дней можно увидеть на рисунке 1B. Снижение внутренней температуры пересекает цикл свет-темнота и изменяется в диапазоне примерно 2 ° C при переходе от активной фазы мышей (свет выключен) к фазе сна (свет включен) (рисунки 1C, D).
Рис. 1. Подготовка ко сну — это терморегулирующее действие. (A) показывает типичное поведение при гнездовании у четырех видов.Гнездо мыши ( Mus musculus , C57Bl6 / J), домашняя кошка ( Felis catus ) свернувшись калачиком, строит гнездо у шимпанзе ( Pan troglodytes verus ) и подстилки ( Homo sapiens ). (B) Пример циркадного температурного цикла в течение 6 дней у самца мыши C57Bl6 / J. (C) Среднее количество переходов от одной и той же мыши за 16 последовательных дней за 2 часа до и после смены освещения. (D) Минимальная температура ( n = 21) во время светлой фазы по сравнению с минимальной ( n = 21) и максимальной температурой ( n = 16) в темной фазе, построенная как изменение от нуля для группы самцов мышей C57Bl6 / J.Данные, представленные в (B – D) , взяты из (Harding et al., Неопубликовано). Все изображения используются с разрешения или разрешения авторских прав. Фотография гнездящегося шимпанзе: Кателийн Купс. Фотография гнездящейся кошки: Изобель Хардинг, спящий человек, доступна по универсальной версии CC0-1.0, а гнездящаяся мышь адаптирована из Дикона (2006).
Температура окружающей среды является критическим фактором, определяющим расход энергии, и невыполнение режима тепловой защиты имеет последствия для гомеостаза энергии (Yu et al., 2018). Например, удаление меха сибирских хомяков увеличивает потребление корма почти на четверть; в то время как в холодных условиях групповое скопление или предоставление материала для гнездования может снизить потребление пищи на 15–20% соответственно (Kauffman et al., 2003; Batavia et al., 2010). Точно так же обеспечение гнездового материала при температурах ниже термонейтральности увеличивает эффективность размножения, обеспечивая больший помет, более высокий вес щенков и снижение смертности детенышей (Gaskill et al., 2013b).
Терморегуляторное поведение перед сном является основной частью поддержания энергетического баланса, по крайней мере, у более мелких млекопитающих, где следствием тепловой неэффективности является повышенная потребность в пище.Однако у более крупных млекопитающих стремление к тепловой подготовке ко сну не слабее. Шимпанзе и другие приматы выбирают свои спальные места на древесине (рис. 1A), по крайней мере, частично, исходя из тепловых характеристик, а в более холодную погоду даже меняют места своих гнезд, чтобы они были более изолированными (Koops et al., 2012; Samson and Hunt, 2012; Stewart и др., 2018). Кроме того, люди активно регулируют температуру во время сна, подсознательно увеличивая открытую площадь поверхности при повышении температуры окружающей среды.При оптимальной комнатной температуре, примерно 19–21 ° C, мы пытаемся установить микроклимат кожи между 31 и 35 ° C, и отклонение от этого диапазона отрицательно влияет на сон (рис. 2A) (Muzet et al., 1984; Okamoto-Mizuno et al., 2003; Raymann et al., 2005). Ключевым фактором использования микроклимата является то, что, по крайней мере у людей, его нельзя заменить нагреванием окружающей среды при той же температуре, возможно, потому, что это нарушает самонастройку, необходимую в течение ночи (Muzet et al., 1984; Raymann. и другие., 2008).
Рисунок 2. Терморегуляция важна для сна человека. (A) Люди используют подстилку для создания теплого микроклимата во время сна. Они активируют центральные гипоталамические механизмы, вызывая сон и периферическую вазодилатацию. (B) Градиент от дистального к проксимальному отделу и снижение внутренней температуры предсказывают начало сна (адаптировано из Krauchi et al., 2000).
Таким образом, терморегуляторное поведение перед сном сохраняется у всех видов млекопитающих, что позволяет предположить, что оно не просто вопрос комфорта, а может иметь более функциональную роль в инициировании и поддержании сна.
«Эффект теплой ванны»
У людей погружение в горячую воду до, но не непосредственно перед периодом сна уменьшает латентность сна и увеличивает глубину сна. Это известно как «эффект теплой ванны» (Horne, Reid, 1985; Parmeggiani, 1987; Bunnell et al., 1988; Shapiro et al., 1989; Jordan et al., 1990; Dorsey et al., 1999). . Фактически, согревание на срок до 4 часов, между 1 и 8 часами перед сном, увеличивает медленный сон (SWS), увеличивает консолидацию NREM и уменьшает REM-сон.Этот эффект олицетворяет ключевую связь между температурой и сном. Согревание в нужное время причинно связано с засыпанием. Однако начало сна происходит при снижении суточной температуры, а NREM ассоциируется с дальнейшим снижением температуры как ядра, так и мозга (Alföldi et al., 1990; Landolt et al., 1995; Kräuchi and Wirz-Justice, 2001). . Многие исследования сна пытались примирить эту противоречащую интуиции взаимосвязь, чтобы объяснить два условия: как нагревание может инициировать сон и быть совместимым с охлаждением тела и как мы можем столкнуться с этим потеплением в «повседневных» условиях.
Оптимальная температура окружающей среды в сочетании с подстилкой, по-видимому, имеет решающее значение для эффективного наступления сна у людей (Haskell et al., 1981; Okamoto-Mizuno et al., 2003; Raymann et al., 2008). Реакция на внешнюю температуру также, по-видимому, важна, поскольку степень расширения сосудов, особенно в руках и ногах (дистальная часть кожи), является хорошим предиктором начала сна (Krauchi et al., 1999). Это расширение сосудов обычно считается частью снижения суточной температуры и наблюдается за 2 часа до начала первого эпизода сна, во время фазы бодрствования (Krauchi et al., 2000). С понижением внутренней температуры это совпадает со снижением самооценки настороженности (Cheisler et al., 1980; van den Heuvel et al., 1998). В экспериментах, в которых участники «самостоятельно выбирали» время отхода ко сну, испытуемые чаще всего выбирали момент, когда температура тела максимально снижалась (Campbell and Broughton, 1994). По мере приближения ко сну внутренняя температура и частота сердечных сокращений падают, и их самое резкое снижение пересекает «выключение света» и начало сна (рис. 2B). В этот момент перепад температуры от проксимального к дистальному отделу равен 1.5 ° C, но при понижении внутренней температуры градиент уменьшается примерно до 0,5 ° C; новая уставка охлаждения достигается сразу после перехода в спящий режим. Самая низкая внутренняя температура наблюдается примерно через 2 часа после «выключения света» и начала сна у Homo sapiens (Krauchi et al., 2000). В естественных условиях повышенный уровень циркулирующего мелатонина также совпадает с понижением внутренней температуры тела перед засыпанием (Krauchi et al., 1997; Krauchi et al., 2006; Logan and McClung, 2019).
Изучение температурной зависимости сна у людей всегда затрудняло нашу способность управлять окружающей средой и избегать дневных колебаний света и температуры.Чтобы обойти это, Йетиш и др. (2015) рассматривали сон в трех географически различных доиндустриальных обществах. Они обнаружили, что начало сна наиболее сильно совпало с понижением температуры окружающей среды. Сон чаще всего начинался после наступления темноты, и весь период сна приходился на снижение температуры окружающей среды. Пробуждение также происходило перед рассветом, когда температура окружающей среды достигла самой низкой точки и совпадала с сужением сосудов, что измерялось температурой пальцев (Йетиш и др., 2015). Изменение температуры в пальцах является хорошей мерой изменения кровотока, и поэтому кажется вероятным, что эти субъекты начали спать в состоянии расширения сосудов, которое постепенно сменялось сужением сосудов до пробуждения (Rubinstein and Sessler, 1990; van Marken Lichtenbelt и др., 2006). Аналогичный результат наблюдал также Han et al. (2018), в условиях лаборатории сна с большим количеством датчиков температуры кожи, распределенных по телу. Это указывало на прогрессирующую вазодилатацию от начала сна до пробуждения.Однако это в основном было представлено в туловище, а руки и ноги не регистрировались (Han et al., 2018).
Циркадный цикл и начало первого эпизода NREM тесно связаны. Если засыпание откладывается из-за недосыпания, нарушается циркадный температурный ритм. Аналогичным образом, задержка снижения внутренней температуры более чем на 2 часа наблюдается у пациентов с расстройствами отсроченной фазы сна (DSPD) (Ozaki et al., 1996; van den Heuvel et al., 1998; Watanabe et al., 2003). Нарушения периферической вазодилататорной реакции достаточно, чтобы нарушить сон. Например, у людей с проблемами периферической вазодилатации (вазоспастических расстройств) латентный период сна больше, чем у здоровых людей (Pache et al., 2001). У пациентов с нарколепсией также сильно изменен градиент температуры кожи от проксимального к дистальному отделу во время дневного бодрствования (Fronczek et al., 2006). Но изменение отношения проксимально-дистально может изменить склонность ко сну. Нагревания сердцевины (проксимальной части кожи) менее чем на 1 ° C, легко в пределах диапазона, встречающегося в течение циркадного дня, достаточно для сокращения латентного периода сна (Raymann et al., 2005). Манипуляции с температурой также могут выборочно и предсказуемо изменить состояние бдительности у пациентов с нарколепсией (Fronczek et al., 2008a, b). Дополнительная работа в клинике показала, что новорожденные в три раза чаще засыпают в течение 30 минут, если градиент их кожи от дистального к проксимальному отделу превышает 2,5 ° C (Abe and Kodama, 2015). Дистальная вазодилатация и более высокая температура стопы у недоношенных новорожденных также коррелируют с более короткими периодами бодрствования (Barcat et al., 2017).
Понимание того, как тепло может встречаться ежедневно, чтобы ускорить эти изменения, инициирующие сон и расширение сосудов, имеет решающее значение.Но кажется, что «эффект теплой ванны» более тонкий, чем считалось ранее. Raymann et al. (2008) расширили парадигму согрева с помощью сделанного на заказ «термокостюма» для управления температурой кожи. Небольшие изменения температуры кожи всего на 0,4 ° C (в диапазоне 31–35) могут сократить латентный период сна без изменения внутренней температуры. Они могут даже способствовать более глубокому сну у более сложных групп пациентов, таких как пожилые люди, страдающие бессонницей (Raymann et al., 2008). Эта последняя группа была особенно восприимчивой к такому регулированию температуры, что подтверждает гипотезу о том, что проблемы со сном у пожилых людей связаны с нарушениями нормальной терморегуляции (Raymann and Van Someren, 2008).
Таким образом, люди и другие млекопитающие демонстрируют терморегулирующее поведение при подготовке ко сну, включая свертывание калачиком, использование подстилки и строительство гнезд. Это может создать микроклимат тепла вокруг кожи, который позволяет засыпать, облегчая расширение сосудов в «дистальных» руках и ногах. Это расширение сосудов может подготовить «проксимальное» ядро к более холодной и неактивной фазе циркадного цикла. Это потепление сохраняется в течение ночи, чтобы поддерживать состояние, позволяющее уснуть, что также допускает селективную вазодилатацию при медленной фазе сна и сужение в фазе быстрого сна и бодрствования.Это достигается при максимальном тепловом КПД сердечника. Причины совпадения охлаждения тела и наступления сна не ясны. Охлаждение тела и мозга как таковой не было показано, что инициирует NREM, но вместо этого является следствием расширения сосудов. Мы могли бы ожидать, что вышестоящий механизм в мозге координирует как начало NREM, так и вазодилатацию, и в следующем разделе мы обсудим, как это может функционировать (Van Someren, 2000).
Нейронный контроль термогенеза и его влияние на сон
Сон — это фундаментальный физиологический процесс, который, как широко считается, необходим для жизни, но его жизненная функция еще не определена.Нейронные цепи, управляющие сном, должны интегрировать информацию как минимум с двух разных входов. Согласно современным представлениям, они известны как процесс C и процесс S, циркадный и гомеостатический вход, соответственно, и являются частью двухпроцессной модели (Borbély, 1982). Переходы от бодрствования к медленному и быстрому сну осуществляются нейронами, которые реагируют на сигналы гомеостатического влечения, которые отслеживают время бодрствования, а также на более заметные сигналы циркадного ритма через супрахиазматическое ядро (SCN).Гомеостатический процесс отслеживает продолжительность периода бодрствования и рассеивает эту нагрузку во время сна. Однако, как мы видели, начало сна также определяется другими факторами: температура окружающей среды, а также уровни сытости, возможности спаривания и необходимость спасаться от хищников — все это определяет подходящий момент для начала медленной фазы сна (Borbély, 1982; Borbély et al. ., 2016; Eban-Rothschild et al., 2017; Logan, McClung, 2019). Нейроны, влияющие на сон, широко распространены по всему мозгу. Это может позволить интегрировать поведенческие и вегетативные факторы в классический гомеостатический и циркадный сон.Напр., Ингибирование дофаминовых нейронов вентральной тегментальной области (VTA) способствует как гнездовому поведению, так и инициации сна (Eban-Rothschild et al., 2016). В то время как гомеостатическое влечение способствует сну после длительного бодрствования, циркадные, поведенческие и вегетативные факторы являются благоприятными условиями для начала сна (см. Рисунок 3). Эти четыре входа работают вместе, чтобы врата спали.
Рисунок 3. Сенсорные и гомеостатические входы, которые закрывают сон. Начало сна определяется четырьмя конкурирующими входными факторами: гомеостатическим влечением ко сну и тремя разрешающими условиями, которые связаны со временем сна, поведенческим входом, циркадным входом и вегетативным входом.Эндокринные факторы также являются ключевой частью каждой категории. Грелин и лептин важны для ощущения голода / сытости соответственно, в то время как мелатонин является ключевым компонентом циркадного ритма. Аденозин и NO могут входить в состав гомеостаза. (Основные факторы, способствующие бодрствованию) Циркадные сигналы благоприятны для бодрствования, а гомеостатическое давление на сон низкое. Поведенческие факторы также способствуют бодрствованию, а вегетативная активность не способствует сну. Ядра, способствующие бодрствованию, управляют кортикальной и таламической возбудимостью, в то же время подавляя такие области, вызывающие сон, как PO и vPAG.Поведенческие потребности в пище и воспроизводстве превосходят потребности сна и теплового комфорта. Поведенческие данные также способствуют пробуждению и могут интегрировать эту информацию в VTA. Гормональные входы, такие как грелин, обнаруживаются в ARC и разрешают сон. Вегетативные сигналы, такие как температура окружающей среды, передаются через спинной мозг и проходят через LPb в PO для интеграции. Цепи, определяющие тепло окружающей среды, не активны, преобладает сужение сосудов и активен BAT. Нейроны AgRP сигнализируют о голоде и подавляют сон. (Внизу — факторы, способствующие медленному сну) Циркадные сигналы теперь разрешают сон, и гомеостатическое давление на сон высокое. Поведенческие факторы также способствуют сну, а вегетативные функции позволяют ему уснуть. В поисках убежища и тепла и после еды разрешается спать. Вегетативные сигналы, такие как температура окружающей среды, передаются через позвоночник и проходят через LPb в PO для интеграции. NOS1-глутаматные нейроны активируются теплом кожи и инициируют как NREM, так и охлаждение тела.Активация сосудорасширяющих цепей и цепей подавления BAT осуществляется через проекции NOS1 на ГАМКергические нейроны LPO или через прямые проекции на DMH и rRPA / RVLM. Поведенческие данные теперь способствуют сну и могут интегрировать эту информацию в VTA. Гормональные входы, такие как лептин, обнаруживаются в ARC и разрешают сон. Нейроны POMC обнаруживают чувство сытости и позволяют спать. NO, оксид азота; NOS1, синтаза оксида азота-1; ПО, преоптическая зона; ПОЛ, боковая преоптическая область; vPAG, вентральный периакведуктальный серый; TMN, туберомаммиллярное ядро; VTA, вентральная тегментальная область; ARC, дугообразное ядро; LPb, латеральный парабрахиальный; LC, голубое пятно; DR — спинной шов; BAT — коричневая жировая ткань; AgRP, пептид, родственный агути; DMH, дорсальный медиальный гипоталамус; rRPA, rostral raphe pallidus; RVLM, рострально-вентролатеральный мозговой слой; ПОМК, проопиомеланокортин (Leshan et al., 2012; Эбан-Ротшильд и др., 2016; Вебер и Дэн, 2016; Yu et al., 2016; Goldstein et al., 2018; Harding et al., 2018; Ю. и др., 2019).
Хотя начало сна и регуляция переходов во время сна могут вовлекать несколько ядер в головном мозге, одна область исторически ассоциировалась с началом медленной фазы сна. Преоптический гипоталамус (ПО) является ключевым местом для инициации NREM, но также считается интегратором терморегуляторной информации, включая защиту от холода и тепла (Szymusiak et al., 2007). Он состоит в основном из средней (MnPO), медиальной (MPO) и латеральной (LPO) областей, которые связаны с большим набором функций от сна до родительского поведения.
Преоптические схемы были предложены как механистическая связь между нагреванием всего тела и индукцией сна (Morairty et al., 1993). Самый простой вариант этой идеи заключается в том, что потепление вызывает активность нейронов, способствующих сну. Действительно, хорошо известно, что теплые стимулы повышают активность ПО (например, по экспрессии c-FOS) (Scammell et al., 1993; Gong et al., 2000). В соответствии с этой идеей, повреждения ПО кошек нарушают как защитное поведение от тепла, так и сокращают общее время сна (Szymusiak et al., 1991). Только значительное потепление этих кошек могло восстановить нормальное количество сна, возможно, за счет компенсации или механизмов за пределами PO (Szymusiak and McGinty, 1986). У крыс поражения PO изменяют поведение тепловых предпочтений, которое впоследствии сводится к более высоким температурам (~ 30 ° C), что способствует восстановлению сна (Ray et al., 2005).В решающих экспериментах использование «термодатчика», имплантированного в РО, нагревание, но не охлаждение, увеличивает дельта-мощность в ЭЭГ (Робертс и Робинсон, 1969; Глотцбах и Хеллер, 1976; МакГинти и др., 1994). Чтобы охарактеризовать преоптические нейроны в этой роли, Alam et al. (1995) повторили этот протокол, используя имплантированный микропривод, и записали свойства преоптических нейронов. Примечательно, что 21% были термочувствительными, и их можно было разделить на две группы — чувствительные к холоду нейроны (CSN) и чувствительные к теплу нейроны (WSN).Около 60% WSN также увеличили свою активность во время NREM (Alam et al., 1995). Во время потепления в мозгу крысы они могут подавлять важные ядра возбуждения, включая спинной шов и задние нейроны гипоталамуса (Krilowicz et al., 1994; Guzmán-Marín et al., 2000; Steininger et al., 2001). При подробном анализе нейронов MnPO Сунцова и соавт. (2002), более 75% продемонстрировали свойства, которые могут способствовать индукции NREM-сна. Это включало постепенное увеличение количества импульсов в фазе быстрого сна и его пика во время сна и, что неожиданно, даже более высокую частоту возбуждения во время фазы быстрого сна (Suntsova et al., 2002). Картирование нейрональных проекций с использованием ретроградных и антероградных индикаторов подтвердило, что MnPO посылает плотную иннервацию в области, способствующие бодрствованию, и может влиять на переходы от бодрствования ко сну путем модуляции латерального преоптического, латерального гипоталамуса и дорсального шва (Uschakov et al ., 2007). Наконец, некоторые нейроны MnPO экспрессируют c-FOS в ответ на депривацию сна, а также могут посылать проекции на LPO (Chou et al., 2002; Zhang et al., 2015).
WSN могут непосредственно определять температуру мозга и, как предполагается, модулируются пирогенами, такими как простагландин E2 (Scammell et al., 1996; Lazarus et al., 2007). Популяция глутаматергических нейронов в средней линии PO экспрессирует канал транзиторного рецепторного потенциального члена 2 (TRPM2), что позволяет напрямую определять локальную температуру мозга. Они могут выполнять тепловую защиту, но также могут модулировать реакцию на жар (Song et al., 2016).
За исключением лихорадки, неясно, может ли потепление кожи вызвать повышение температуры мозга, которое может ощущаться WSN (Tan et al., 2016; Siemens and Kamm, 2018).Вместо этого более вероятен синаптический путь. Нейроны, которые получают афферентную информацию о температуре, но не являются непосредственными «датчиками» температуры, были выделены термином «активируемые теплом» нейроны (Tan and Knight, 2018). Гипоталамус MnPO и MPO получает сенсорные афференты, передающие тепловую информацию от кожи (Hammel, 1968; Boulant and Gonzalez, 1977; Morrison and Nakamura, 2011) (Рисунок 4). Сенсорные нейроны передают информацию о температуре окружающей среды через спинной мозг к глутаматергическим релейным нейронам и к подобластям латерального парабрахиального ядра (LPb).LPb может также получать информацию от других частей тела, таких как внутренние органы, а затем передавать эти сигналы в области MnPO и MPO (Nakamura and Morrison, 2008, 2010). В первой точке интеграции глутаматергические нейроны передают синапс избыточного тепла в глутаматергические нейроны MnPO, выход которых инициирует охлаждение, способствуя вазодилатации и выключая термогенез бурого жира (Morrison and Nakamura, 2011) (Рисунок 4). Что это за нейроны в ПО, которые реагируют на потепление? Недавняя работа с использованием фотометрии GCaMP6 показала, что эти нейроны могут реагировать на внешние тепловые воздействия при температуре от 30 до 40 ° C, а секвенирование РНК выявило в них экспрессию полипептида, активирующего аденилатциклазу гипофиза (PACAP) и нейротрофического фактора мозга (BDNF) (Tan и другие., 2016). Эти нейроны являются преимущественно ГАМКерными и при активации могут вызывать переохлаждение. Они функционируют, по крайней мере частично, путем ингибирования глутаматергических нейронов дорсального медиального гипоталамуса (DMH), которые стимулируют термогенез BAT (Tan et al., 2016). Еще одна популяция ГАМКергических нейронов, которые действуют через ДМГ, также была обнаружена в соседнем вентральном ПОЛ (Zhao et al., 2017).
Рисунок 4. Интеграция сигнала в преоптическом гипоталамусе. Тепло на коже стимулирует сенсорные сигналы через LPb к преоптическим нитрергико-глутаматергическим нейронам, которые инициируют одновременную медленную медленную фазу и охлаждение тела.Это может быть связано с активацией отдельных ГАМКергических нейронов для сна и гипотермии в МПО и ПОЛ, но они также могут активировать галинергические-ГАМКергические нейроны, чтобы инициировать сон и охлаждение тела. Синаптическая роль NO в этих цепях неизвестна, но потенциальные сайты отмечены. NREM инициируется ингибированием ядер возбуждения, включая TMN и LH. Вероятно, будут задействованы и другие. Охлаждению тела способствует активация DMH и ингибирование нейронов rRPA, что вызывает вазодилатацию и подавление термогенеза BAT.Входы в латеральную парабрахиальную и преоптическую области модулируются посредством нейрон-опосредованного ингибирования AgRP из дугообразного отдела. Они обнаруживают голод и прерывают NREM. Сытость вызывает активацию нейронов POMC, которые также экспрессируют TRPV1, являются пермиссивными для NREM и вызывают локальное ингибирование нейронов AgRP. Нитрергически-глутаматные нейроны могут отвечать на лептин через лептин Rb, как и нейроны AgRP и POMC. Они или отдельная местная популяция также могут реагировать на изменения температуры мозга через ионный канал TRPM2.NO, оксид азота; NOS1, синтаза оксида азота-1; ПО, преоптическая зона; ПОЛ, боковая преоптическая область; vPAG, вентральный периакведуктальный серый; TMN, туберомаммиллярное ядро; ARC, дугообразное ядро; LPb, латеральный парабрахиальный; BAT — коричневая жировая ткань; AgRP, пептид, родственный агути; ПОМК, проопиомеланокортин; DMH, дорсальный медиальный гипоталамус; rRPA, rostral raphe pallidus; RVLM, рострально-вентролатеральный мозговой слой; TRPM2, временный катионный канал рецепторного потенциала; TRPV1, переходный рецепторный потенциал катионного канала валлиноид-1; ГАЛ, Галанин (Лешан и др., 2012; Вебер и Дэн, 2016; Yu et al., 2016; Goldstein et al., 2018; Harding et al., 2018; Jeong et al., 2018; Тан и Найт, 2018; Ю. и др., 2019).
PO представляет собой очень разнообразную область с множеством перекрывающихся популяций, но только некоторые из этих нейронов были функционально охарактеризованы (Moffitt et al., 2018). Например, нейроны ГАМК-галанина связаны как со сном, так и с родительским поведением, но также существуют популяции галанин-глутаматных нейронов (Sherin et al., 1998; Wu et al., 2014; Moffitt et al., 2018). PACAP / BDNF, TRPM2-глутаматные и нитрергико-глутаматные нейроны связаны с защитой от тепла и лихорадкой, но существует множество других субпопуляций (Song et al., 2016; Tan et al., 2016; Harding et al., 2018). Хотя последнее также связано с индукцией сна, популяции ГАМКергических-нитрергических нейронов были обнаружены, но не охарактеризованы (Harding et al., 2018; Moffitt et al., 2018). Учитывая обширное разнообразие подтипов нейронов ПО (Moffitt et al., 2018), особенно важны такие методы, как c-FOS-зависимая маркировка активности, позволяющая функционально отделить определенные цепи от окружающей среды (Zhang et al., 2015). Область PO, включающая как MPO, так и LPO, отвечает на восстановительный сон, сон после лишения сна, выражением c-FOS. Эти же области возбуждаются α 2A -адренергическим агонистом и седативным средством, дексмедетомидином (DEX) (Zhang et al., 2015). Чтобы понять, имеют ли эти физиологии одну и ту же схему, Zhang et al. (2015) использовали c-FOS-зависимую маркировку активности, чтобы отделить нейроны, активированные восстановительным сном или DEX, от других нейронов PO, которые реагируют на различные внешние и гомеостатические стимулы.Эти нейроны экспрессировали возбуждающий рецептор hM 3 d q DREADD, так что, когда этим мышам давали клозапин N -оксид, активировался только этот уникальный ансамбль. Это привело к консолидированному сну NREM, совместимому с восстановительным сном или седативным действием. Однако ансамбли РО, помеченные либо восстановительным сном, либо DEX, также вызывали гипотермию (Zhang et al., 2015). Фактически, практически все седативные препараты и общие анестетики, используемые в клинической практике, вызывают перераспределение тепла от ядра к периферии за счет расширения сосудов и, без нагревания, гипотермии (Díaz and Becker, 2010; Sessler, 2016).Это предполагает, что основная схема ПО может связывать естественную индукцию сна, индукцию охлаждения тела и механизмы действия препаратов класса седативных средств.
Мы предположили, что сами цепи терморегуляции могут играть важную роль в улучшении сна. Это также могло бы объяснить склонность внешнего или прямого потепления PO вызывать медленный сон. Мы снова использовали метку активности, но на этот раз пометили только те преоптические ансамбли, которые получали теплую сенсорную информацию. Реактивация этих «меченых теплом» нейронов вызывала одновременную NREM и охлаждение тела (Harding et al., 2018). Эти нейроны экспрессировали смесь маркеров клеточного типа, включая везикулярный транспортер глутамата 2 (VGLUT2), глутаматдекарбоксилазу (GAD67) и синтазу оксида азота 1 (NOS1). Когда мечение активности повторяли у мышей NOS1-CRE, они также испытывали одновременное NREM и охлаждение тела. Однако при повторении у мышей vGAT-CRE наблюдались только NREM и небольшое охлаждение тела. Поскольку эти нейроны NOS1 экспрессируют VGLUT2, наши данные предполагают наличие четкой нитрергико-глутаматергической цепи для связывания термальной сенсорной информации с началом NREM, которая может располагаться выше ГАМКергического «переключателя» сна (Harding et al., 2018). В этой схеме внешнее тепло является допустимым состоянием для инициации NREM. Без этого сенсорного входа начало NREM подавлено. Мы думаем, что это согласуется с данными, показывающими, что внешнее потепление способствует сну у людей и животных, а также предоставляет возможный механизм того, почему млекопитающие ищут места гнездования: создание микроклимата тепла кожи, позволяющего спать. Мы еще не знаем, используют ли нейроны NOS1 оксид азота (NO) в синаптической передаче. Однако NO участвует в модуляции возбуждения в других областях мозга (Геращенко и др., 2008; Калинчук и др., 2010; Cespuglio et al., 2012; Morairty et al., 2013; Ю. и др., 2019).
Что находится ниже по течению от нитрергически-глутаматных нейронов MnPO / MPO? Локальная преоптическая область содержит несколько популяций галаниновых нейронов, как возбуждающих, так и тормозных (Moffitt et al., 2018). Недавно эксперименты показали, что активация галаниновых нейронов вентролатеральной преоптической области (VLPO) может вызывать как NREM, так и гипотермию (Kroeger et al., 2018). Точно так же активация галаниновых нейронов при ПОЛ также может вызывать NREM и гипотермию (Ma et al., 2019). Последнее имеет параллели с активацией ГАМКергических нейронов, меченных активностью, во время восстановительного сна (Zhang et al., 2015). Поскольку известно, что MnPO посылает проекции как в LPO, так и в VLPO, мы предположили, что нейроны ГАМК-ергического галанина могут быть мишенями для нейронов нитрергического глутамата (Uschakov et al., 2007). В VLPO активация галаниновых нейронов с помощью рецепторов DREADD способствовала большему количеству NREM-сна, когда мыши были ближе к термонейтральности (29 ° C) и когда гипотермия притуплялась нагреванием до 36 ° C.Термонейтральность, по-видимому, обеспечивает оптимальное восстановление быстрого сна по сравнению с условиями окружающей среды (22 ° C) или тепла (36 ° C) (Kroeger et al., 2018). Это согласуется с идеей узкого диапазона температур для оптимального быстрого сна (Cheisler et al., 1980; Szymusiak and Satinoff, 1981). Галаниновые нейроны в ПОЛ, по-видимому, необходимы для активации гомеостатических механизмов, запускающих восстановительный сон. Делеция этих нейронов с использованием экспрессии каспазы устраняет отскок дельта-мощности после лишения сна (Ma et al., 2019). Необходимы дополнительные данные, чтобы подтвердить, являются ли ПОЛ-галанин подлинными мишенями для нейронов с нитрергическим глутаматом. У последних могут быть и другие дальние прогнозы.
Термогенез связывает сон с энергетическим гомеостазом
Терморегуляция, в частности тепловая неэффективность, влияет на гомеостаз энергии и изменяет потребности в питании. Это дополнительный гомеостатический двигатель, который добавляет свое собственное «давление» для модуляции сетей сна (рис. 3). После еды адипоциты выделяют гормон лептин.Этот гормон указывает на избыточное потребление энергии и препятствует кормлению. Лептин действует через хорошо установленные пути в дугообразном ядре гипоталамуса, где он ингибирует NPY-экспрессирующие нейроны AgRP (Williams et al., 2009). Однако рецепторы лептина есть и в других частях ЦНС, в том числе в ПО гипоталамусе. Глутаматергические нейроны PO, экспрессирующие рецептор лептина (leptinRb), возбуждаются (они экспрессируют c-FOS) при повышении температуры окружающей среды (Yu et al., 2016). Это приводит к снижению расхода энергии за счет подавления термогенеза и снижению потребления пищи (Zhang et al., 2011; Ю. и др., 2016). Нейроны, коэкспрессирующие NOS1 и leptinRb, были идентифицированы в других частях гипоталамуса, и они также могут ингибировать термогенез (Leshan et al., 2012). Следовательно, кажется вероятным, что существует некоторое совпадение между преоптическими глутаматергическими-лептин-Rb нейронами и популяциями NOS1, идентифицированными Harding et al. (2018). Аналогичным образом, многие BDNF / PACAP в PO экспрессируют VGLUT2, и недавно было показано, что субпопуляция этих нейронов, экспрессирующих c-FOS в ответ на теплый стимул, коэкспрессирует лептинRb, что предполагает еще большее перекрытие между этими популяциями (Moffitt et al., 2018). Помимо регуляции энергии, передача сигналов лептина, по-видимому, играет более прямую роль во сне. Системное введение рекомбинантного лептина мышам, лишенным пищи, увеличивает продолжительность как NREM, так и REM-сна, в то время как мыши с дефицитом лептина (ob / ob) имеют фрагментированный сон, а также более низкую среднюю температуру тела (Sinton et al., 1999; Laposky et al. , 2006). Ключевой остающийся вопрос заключается в том, обеспечивают ли нейроны NOS1, инициирующие медленный сон и охлаждение тела, более широкую связь между сном и энергетическим гомеостазом (Harding et al., 2018). Эти схемы представлены на Рисунке 4.
Последние данные позволили по-новому взглянуть на то, как энергетический баланс может влиять на сон. Goldstein et al. (2018) непосредственно оценили влияние активности нейронов AgRP / POMC в дугообразном ядре на влечение ко сну. Нейроны AgRP могут определять потребление энергии и считаются «сенсорами голода», подавляемые как циркулирующим лептином, так и инсулином. POMC противодействуют действию нейронов AgRP и активируются лептином (Cowley et al., 2001).Активность AgRP способствует поиску пищи даже за счет сна. Но если мышей лишены пищи, ингибирование этих нейронов спасает поведение во сне за счет еды (Goldstein et al., 2018). Поскольку тепловая неэффективность приводит к увеличению кормления, мы ожидаем, что схема связывает тепловые ощущения с контролем аппетита. В соответствии с этой идеей Jeong et al. (2018) показали, что нейроны POMC экспрессируют канал временного рецепторного потенциала ваниллоида-1 (TRPV1). Оптогенетическая активация этих нейронов вызывает подавление питания (Jeong et al., 2018). Хотя это исследование не оценивало сон, активация нейронов POMC Goldstein et al. (2018) может спасти животных, лишенных пищи (Goldstein et al., 2018). Это может быть связано с тем, что нейроны POMC плотно иннервируют области, способствующие сну, включая PO, и могут ингибировать локальные ГАМКергические интернейроны (Elias et al., 1999; Wang et al., 2015; Weber and Dan, 2016). Нейроны POMC также ингибируют нейроны AgRP, которые проецируются в несколько областей, способствующих сну, включая PO, вентральное периакведуктальное серое (vPAG) и парабрахиальное ядро (Pb) (Betley et al., 2013; Wang et al., 2015; Вебер и Дэн, 2016; Weber et al., 2018) (см. Рисунок 4). Дугообразные нейроны NPY, которые обычно стимулируют прием пищи, также подавляют термогенез BAT и, таким образом, могут иметь сходные роли в терморегуляторных связях со сном (Shi et al., 2013). Эти вегетативные сигналы интерпретируются как сильные поведенческие побуждения, например, поиск пищи.
Таким образом, кажется вероятным, что существует значительное перекрытие между популяциями нейронов, которые регулируют начало сна, термогенез и энергетический гомеостаз.Наступление сна можно частично контролировать путем интеграции этих сенсорных входных сигналов, включая температуру окружающей среды и энергетический статус. Менее понятно, почему ограничение сна с помощью этих входов было бы полезным.
Депривация сна нарушает терморегуляцию и энергетический баланс
Архитектура сна сильно зависит от тепловых факторов, но последствием полной потери сна является радикальное изменение терморегуляции и энергетического баланса. У крыс хроническая депривация полного сна и избирательная депривация быстрого сна, при использовании метода дискового над водой в течение многих дней, приводит к глубоким физиологическим эффектам и, в конечном итоге, к смерти (Everson et al., 1989). На ранних стадиях у этих крыс наблюдалось усиление метаболической функции, включая внутреннюю температуру тела, а вместе с ней и увеличение потребления пищи. Однако повышение температуры быстро прекратилось, и у крыс постепенно развивалось переохлаждение. Они также переместились в более теплые части температурного градиента по мере того, как их недосыпание усилилось (Prete et al., 1991). Это может быть стратегия сохранения энергии, снижение тепловой нагрузки, повышение аппетита и одновременное охлаждение тела (Rechtschaffen and Bergmann, 1995).Подобные стратегии наблюдаются у вялых животных при недостатке пищи (Ruf and Geiser, 2015). У недосыпающих животных эта стратегия в конечном итоге не увенчалась успехом, поскольку крысы быстро теряли вес (Bergmann et al., 1989; Everson et al., 1989; Rechtschaffen and Bergmann, 1995). Лишение сна, по-видимому, либо увеличивает метаболические потребности животного, либо другими способами способствует чрезмерной потере тепла, возможно, за счет чрезмерной активации цепей, инициирующих NREM, которые вызывают расширение сосудов.
Одним из механизмов, с помощью которого млекопитающие, и в частности мелкие грызуны, выделяют тепло, является термогенез коричневой жировой ткани (НДТ).Это также ключевой механизм регулирования энергетического гомеостаза. Несвязывающий белок 1 (UCP-1) является ключевым компонентом термогенеза в коричневой жировой ткани (BAT). Он отделяет цепь переноса электронов от АТФ-синтазы, облегчая выработку тепла за счет рассеяния протонного градиента без производства АТФ и компенсаторного метаболизма (Cannon and Nedergaard, 2004). У мышей с нокаутом UCP-1 наблюдается ослабление гомеостатического восстановления после лишения сна. Они также демонстрируют притупленный эффект индукции сна при более высоких температурах, наблюдаемый у контрольных мышей (Szentirmai and Kapas, 2014).Точно так же агонисты β3-адренергических рецепторов, которые активируют термогенез BAT, обычно вызывают сон у контрольных мышей, но этот ответ устраняется у мышей с химической деафферентацией BAT (Szentirmai and Kapás, 2017). Эти данные предполагают, что термогенез BAT, опосредованный UCP-1, полезен как для восстановления сна (сон после лишения сна), так и для индукции NREM-сна. UCP-1 также может играть роль в облегчении медленного сна во время системного воспаления (Szentirmai and Kapas, 2018). Тепло, генерируемое этими механизмами, может активировать сенсорные рецепторы в коже и, таким образом, запускать медленный сон (Harding et al., 2018).
Торпор и гибернация: слишком холодно, чтобы спать?
Взаимодействие охлаждения тела и медленного сна предполагает, что энергетический гомеостаз является важным фактором для сна, но естественно спросить, есть ли какая-либо связь с более экстремальными состояниями (рис. 5). Когда потребность в экономии энергии достаточно высока, многие млекопитающие жертвуют сном, чтобы принять альтернативную стратегию терморегуляции — ежедневное оцепенение или сезонную гибернацию (Ruf and Geiser, 2015). Ежедневное оцепенение — это состояние переохлаждения, вызванное нехваткой пищи.Млекопитающие, которые используют ежедневное оцепенение, такие как джунгарские хомяки ( Phodopus sungorus ), обычно понижают свою внутреннюю температуру до 15–20 ° C на много часов, но у многих видов суточное оцепенение может колебаться от 10 ° C и выше. как 30 ° C (Ruf and Geiser, 2015) (рисунок 5).
Рис. 5. Переходы между состояниями при различных температурах ядра. Сон, анестезия и оцепенение находятся в непрерывном потоке снижения внутренней температуры, что напрямую влияет на мощность ЭЭГ. В среднем приступы NREM холоднее, чем приступы бодрствования, тогда как температура мозга во время REM выше.Типичный пример NREM EEG при температуре приблизительно 37 ° C показан зеленым цветом. Седативные и анестетические средства вызывают дельта-колебания на ЭЭГ, а также гипотермию. Дексмедетомидин (DEX; 100 мкг / кг IP) вызывает устойчивый седативный эффект, но сила дельта-колебаний подавляется. Пример показан синим цветом через 2 часа после закачки при температуре ядра примерно 26 ° C. Если животному в теплой камере вводить такую же дозу DEX, сила дельта-колебаний восстанавливается. Пример показан красным через 2 часа после закачки с температурой ядра 34 ° C.Некоторые млекопитающие используют ежедневное оцепенение для экономии энергии в периоды нехватки пищи. В среднем они составляют от 15 до 20 ° C, но могут колебаться от 10 до 30 ° C. Приблизительно при 21 ° C состояние оцепенения может вызвать недосыпание, которое приводит к восстановительному сну при нагревании до 37 ° C. Искусственная гипотермия, иногда известная как синтетическое оцепенение, может быть вызвана 5-АМФ (0,5 г / кг IP). Это также вызывает дельта-колебания, которые подавляются переохлаждением. Пример показан синим цветом через 1,5 часа после закачки при температуре ядра примерно 23 ° C.Ниже примерно 10 ° C ЭЭГ изоэлектрическая, и колебания не видны. У гибернаторов есть периоды между периодами эвтермии с нормальной мощностью ЭЭГ, и обнаруживаются переходы между бодрствованием и медленной фазой сна и бодрствованием в фазе быстрого сна. Образцы видов помечены температурой, при которой они наблюдались в течение ежедневного оцепенения или гибернации. Это отражает условия окружающей среды, важные для измерений ЭЭГ, но не является строгой иерархией. Примеры ЭЭГ взяты из (Harding et al., Неопубликовано), за исключением примера гибернации, который адаптирован из Frerichs et al.(1994). IP, внутрибрюшинно; 5-АМФ, аденозинмонофосфат. Джунгарский хомяк ( Phodopus sungorus ), златопланый суслик ( Callospermophilus lateralis ), толстохвостый карликовый лемур ( Cheirogaleus medius ), полярный суслик ( Urocitellus ). Данные адаптированы из Frerichs et al. (1994); Руф и Гейзер (2015) и Вязовский и др. (2017).
Ряд млекопитающих, от сусликов до бурых медведей, также используют ежегодные стратегии гибернации для выживания и размножения зимой (Carey et al., 2003; Руф и Гейзер, 2015). У хибернаторов внутренняя температура обычно составляет от 0 до 10 ° C и поддерживается в течение недель или месяцев. Внешняя температура может опускаться ниже 0 ° C, а внутренняя температура поддерживается только на несколько градусов выше при 1% эвтермической (нормальной температуры) скорости метаболизма (Carey et al., 2003). В крайних случаях, например, у арктического суслика ( Urocitellus parryii ), брюшная и периферическая температура может быть стабильной на уровне около -2 ° C, в то время как голова и шея чуть выше 0 ° C (Barnes, 1989; Boyer and Barnes, 1999 ).Только при температуре тела около 0 ° C скорость метаболизма повышается, чтобы защитить животное от замерзания, что указывает на чрезвычайно низкую уставку температуры (Buck and Barnes, 2000) (рис. 5).
Животные, находящиеся в состоянии ежедневного оцепенения или гибернации, входят в состояние бездействия или покоя, но мощность сигнала ЭЭГ, наблюдаемого у этих животных, зависит от температуры: чем ниже температура тела, тем ниже мощность ЭЭГ (Deboer, 1998). При температуре ядра около 22 ° C или выше частотная составляющая остается в пределах дельта-диапазона 1–4 Гц и может быть классифицирована как сон, хотя мощность ЭЭГ значительно снижается (Walker et al., 1981; Даан и др., 1991). Однако при более низких температурах это не так. Если температура ядра и мозга достаточно низкая, то мощность ЭЭГ падает ниже порога атрибуции состояний сна (рис. 5). При температуре мозга от 10 ° C до 25 ° C дельта-колебания пониженной мощности все еще можно идентифицировать в сигнале ЭЭГ, тогда как ниже 10 ° C сигнал является изоэлектрическим (Walker et al., 1977). Неясно, насколько мощность этих колебаний важна для естественной функции сна.Например, у карликовых лемуров ( Cheirogaleus medius ), находящихся в спячке при низких температурах окружающей среды всего 5 ° C, записи ЭЭГ являются изоэлектрическими, а признаки медленного или быстрого сна отсутствуют. В этом случае только при согревании наблюдался сон (REM) (Krystal et al., 2013). Было высказано предположение, что оцепенение может быть формой лишения сна. Например, выздоровление джунгарских хомяков от ежедневного оцепенения с температурой ядра и мозга около 23 ° C приводит к периоду восстановительного сна с повышенной мощностью в дельта-диапазоне (Deboer, 1998).Точно так же этот сон может быть отложен из-за недосыпания, что предполагает накопление недосыпания во время оцепенения (Deboer, Tobler, 1994; Palchykova et al., 2002). Хотя этот недосып, измеренный с помощью дельта-мощности, накапливается во время оцепенения, он происходит почти в три раза медленнее при температуре мозга ниже 27 ° C по сравнению с временем бодрствования (Deboer and Tobler, 2003). Однако сравнение ЭЭГ восстановительного сна после депривации сна или оцепенения выявило различия в активности корковой сети, что позволяет предположить, что оцепенение не полностью эквивалентно депривации или естественному сну (Вязовский и др., 2017). Таким образом, может существовать критическая температура, ниже которой нарушается функция сна.
Чтобы понять взаимосвязь между сном и температурой у спящих людей, исследователи сравнили животных, которые впадают в спячку при разных температурах окружающей среды. Животные, впадающие в спячку при низких температурах, такие как арктические суслики ( Urocitellus parryii ), ненадолго прогреваются до уровня, сопоставимого с бодрствованием (36–37 ° C). Это периоды межквартальной эутермии (Boyer, Barnes, 1999; Carey et al., 2003). В эти периоды потепления белки переходят от бодрствования к медленному сну, а затем к быстрому сну, прежде чем вернуться в спячку (Daan et al., 1991). Зимняя спячка суслика с золотым покровом ( Callospermophilus lateralis ) в более теплых лабораторных условиях (22 ° C) вызвала непрерывный медленный сон (Walker et al., 1981). Во время гибернации при более низких температурах окружающей среды 11 ° C минимальная температура мозга, а не продолжительность периода гибернации, была лучшим предиктором дельта-силы отскока во время последующей эутермии между периодами.Те же авторы заметили, что при этой температуре (11 ° C) эвтермический (36–37 ° C) период после спячки почти на 70–80% состоит из NREM-сна, тогда как животные, находящиеся в спячке при 21 ° C, проводят только 40% своего времени. эвтермический период в NREM (Larkin and Heller, 1996). Это указывает на то, что температура, при которой наступает спячка, влияет на степень накопления недосыпания (см. Рисунок 5). Конечно, есть различия между видами. Когда европейские суслики входят в эвтермический период после зимней спячки в 5 лет.5 ° C, время, проведенное в медленном сне, пропорционально продолжительности периода гибернации, а не температуре как таковой (Strijkstra and Daan, 1997). В совокупности эти данные предполагают, что восстановительный компонент сна зависит от температуры.
Такая же температурная зависимость сна наблюдается у спящих приматов. Когда карликовые лемуры ( Cheirogaleus medius ) выбирают зимнюю спячку при более высоких температурах, их ЭЭГ напоминает медленный и быстрый сон, и эпизоды эутермии исчезают (Dausmann et al., 2004; Кристал и др., 2013). У других видов лемуров ( C. crossleyi и C. sibreei ), которые занимают более прохладную экологическую нишу, сон постоянно отсутствует во время фазы оцепенения, но возвращается в периоды межпадовой эутермии (Blanco et al., 2016). Черные медведи ( Ursus americanus ), которые всегда впадают в спячку при более высоких температурах 32–34 ° C и активно защищают эту заданную температуру, также не проявляют возбуждения между схватками, наблюдаемого у более мелких млекопитающих (Tøien et al., 2011 ).Как и у карликовых лемуров, эти более высокие температуры, по-видимому, позволяют бурым медведям проводить много времени в медленном сне (Tøien et al., 2015). Защита более низких уставок температуры у крупных млекопитающих имеет примечательные параллели с людьми, находящимися под наркозом. У людей, которым вводят седативный DEX или анестетик пропофол, порог дрожи снижается до 32-34 ° C соответственно (Matsukawa et al., 1995; Talke et al., 1997; Sessler, 2016). Следовательно, спячка может быть слишком холодной, чтобы облегчить сон, а эпизоды межконтинентальной эвтермии, длящиеся 12–24 ч, могут позволить восстановить процессы сна (Carey et al., 2003).
Нейронная схема, которая вызывает оцепенение и / или гибернацию, неизвестна. Однако возможно, что он использует компоненты естественной схемы сна-бодрствования. Например, NOS1-глутаматные нейроны в ПО, которые вызывают NREM-сон и устойчивую гипотермию (Harding et al., 2018), могут в более холодном климате играть роль в индукции оцепенения или гибернации, но с другими поведенческими и средовыми триггерами.
Таким образом, медленный сон в состоянии умеренного охлаждения тела может быть предпочтительным биологическим состоянием, но очевидно, что в экстремальных условиях, при выживании зимой или в периоды нехватки пищи восстанавливающие эффекты сна, по крайней мере частично, приносятся в жертву для сохранения энергии.Поскольку сон можно поддерживать только при более высоких температурах, он энергетически дороже, чем оцепенение или гибернация. При более низких температурах мозга и ядра недосыпание накапливается почти в три раза медленнее, чем во время бодрствования (Deboer and Tobler, 2003).
Зачем связывать NREM Sleep and Body Cooling?
Напомним, что сон у грызунов связан с температурным циклом: переходы от бодрствования к медленному сну совпадают с более прохладными телом и мозгом, чему способствует расширение сосудов хвоста. Действительно, эффективная терморегуляция и гнездование создают теплый микроклимат, который играет роль в стимулировании медленного сна и охлаждении тела.Мы предположили, что нейроны PO одновременно получают теплую тепловую информацию от кожи и одновременно координируют начало медленного сна и охлаждение тела (Harding et al., 2018). Переходы к бодрствованию или быстрому сну сопровождаются сужением сосудов и потеплением мозга (Alföldi et al., 1990; Imeri and Opp, 2009). Абсолютное изменение температуры мозга при каждом переходе NREM невелико, около 0,2–0,4 ° C, но может достигать больших значений, сопоставимых с общим суточным изменением температуры (примерно 2 ° C) во время продолжительных периодов сна.У людей внутренняя температура надежно падает примерно за 2 часа до начала сна, и первый эпизод медленной фазы сна с большей вероятностью произойдет в самый резкий момент падения температуры. Похоже, то же самое происходит с температурой мозга (Landolt et al., 1995). Эта скорость снижения может быть максимальной, когда схема PO активирована максимально, что в наибольшей степени способствует медленному сну. Другие сенсорные входы, такие как чувство насыщения, также разрешают сон, и их входные данные объединяются для определения точного момента начала медленной фазы сна.Эти изменения температуры могут иметь прямую роль в восстановительных функциях сна.
Одна из первых гипотез относительно более низких температур, совпадающих с медленным сном, заключалась в том, что они существуют специально для охлаждения мозга (McGinty and Szymusiak, 1990). Было высказано предположение, что более низкая температура мозга снизит церебральный метаболизм, сохранит энергию и поможет другим функциям от иммунной регуляции до циркадной координации (McGinty and Szymusiak, 1990). Также было предложено сохранение энергии для сна в целом (Berger and Phillips, 1995).Однако мы видели, что, когда млекопитающие всех размеров отдают приоритет сохранению энергии, крайние уровни гипометаболизма в оцепенении и гибернации выбираются за счет сна (Ruf and Geiser, 2015). Это говорит о том, что сохранение энергии не является основной функцией сна. Действительно, по оценкам использования энергии в течение 24 часов, стоимость сна составляет 85–95% от метаболических затрат при бодрствовании (Jung et al., 2011; Abreu-Vieira et al., 2015; Hibi et al., 2017). ).
Вполне возможно, что пониженная температура имеет более прямую функцию в мозге.При температурах 20 ° C или ниже, когда накапливается недосыпание, наблюдаются морфологические изменения в дендритных шипах (Peretti et al., 2015). Гибернаторы могут подвергаться синаптическому ремоделированию во время холода, как и животные в искусственном оцепенении, вызванном 5′-аденозинмонофосфатом (Попов, Бочарова, 1992; Magariños et al., 2006; Popov et al., 2007). В последнем случае общее количество синапсов уменьшается (GM). Наличие этих процессов может объяснить, почему сон, как восстановительный процесс, подавляется при более низких температурах.Большие изменения в экспрессии генов также наблюдаются как в головном мозге, так и во всем теле у гибернаторов (Williams et al., 2005). Более низкие температуры, особенно в головном мозге, могут вызывать экспрессию так называемых белков «холодового шока», включая индуцируемый холодом РНК-связывающий белок (CIRP) и РНК-связывающий мотив-белок 3 (RBM3) (Morf et al., 2012; Peretti et al. др., 2015; Hoekstra et al., 2019). Охлаждение тела и мозга во время естественного сна невелико как из-за снижения суточной внутренней температуры, так и из-за снижения температуры мозга при каждом переходе в NREM, но недавние данные показывают, что этого достаточно для увеличения экспрессии CIRP и, таким образом, влияния на экспрессию других генов, включая циркадные гены Period и Clock (Morf et al., 2012; Hoekstra et al., 2019). Это важно, поскольку изменения температуры коры в большой степени зависят от переходов сон-бодрствование, и переход в NREM-сон может затем влиять на экспрессию генов часов, чтобы управлять дальнейшими изменениями транскрипции. У мышей без CIRP лишение сна приводит к сокращению быстрого сна на 50%, что свидетельствует о силе этого механизма (Hoekstra et al., 2019). Это также обеспечивает один возможный механизм, с помощью которого мозг может отслеживать время, проведенное в медленном сне.
Обширные нейронные связи, которые перекрестно регулируют использование энергии, индукцию сна и температуру тела (см. Рисунок 3), намекают на то, что функция сна играет важную роль в энергетическом гомеостазе.Температурная зависимость накопления задолженности во сне, которая замедляется при более низких температурах, предполагает, что этот долг по своей природе является метаболическим процессом. Наконец, синхронизированные изменения температуры мозга во время сна могут координировать экспрессию генов, важных для функций сна, одновременно внося свой вклад в механизм измерения времени, проведенного во сне.
Авторские взносы
EH написал рукопись и нарисовал рисунки. Все авторы обсудили и отредактировали рукопись.
Финансирование
Эта работа финансировалась Британским научно-исследовательским институтом деменции, который получает финансирование от UK DRI, финансируется Британским советом медицинских исследований, Обществом Альцгеймера и Исследовательским центром Альцгеймера Великобритании (NF и WW), а также Wellcome Trust (107839 / Z / 15 / Z в NF и 107841 / Z / 15 / Z в WW).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Все изображения используются с разрешения или разрешения авторских прав. Спящий человек (рисунок 1А) доступен под универсальным стандартом CC0-1.0. Мы благодарны Кателин Купс за фотографию гнездящейся шимпанзе, Изобель Хардинг за фотографию спящей кошки и Алии душ Сантуш за вычитку рукописи.
Список литературы
Абреу-Виейра, Г., Сяо, К., Гаврилова, О., и Рейтман, М. Л. (2015). Интеграция температуры тела в анализ расхода энергии у мыши. Мол. Метаб. 4, 461–470. DOI: 10.1016 / j.molmet.2015.03.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Алам, М. Н., МакГинти, Д., и Шимусиак, Р. (1995). Нейрональные разряды преоптических / передних гипоталамических термочувствительных нейронов: связь с медленным сном. Am. J. Physiol. 269 (5 баллов 2), R1240 – R1249. DOI: 10.1152 / ajpregu.1995.269.5.R1240
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Альфельди, П., Рубичек, Г., Черни Г. и Обал Ф. Дж. П. А. (1990). Температура мозга и ядра и периферическая вазодвигательная реакция во время сна и бодрствования при различных температурах окружающей среды у крысы. Pflügers Archiv. 417, 336–341. DOI: 10.1007 / bf00371001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Barcat, L., Decima, P., Bodin, E., Delanaud, S., Stephan-Blanchard, E., Leke, A., et al. (2017). Дистальное расширение сосудов кожи способствует быстрому засыпанию недоношенных новорожденных. Дж.Sleep Res. 26, 572–577. DOI: 10.1111 / jsr.12514
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Батавиа, М., Мацусима, А., Эбоигбоден, О., и Цукер, И. (2010). Влияние теплоизоляции шерсти и температуры окружающей среды на потребление энергии и рост молоди сибирских хомяков. Physiol. Behav. 101, 376–380. DOI: 10.1016 / j.physbeh.2010.07.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бергер, Р. Дж., И Филлипс, Н.Х. (1995). Энергосбережение и сон. Behav. Brain Res. 69, 65–73. DOI: 10.1016 / 0166-4328 (95) 00002-B
CrossRef Полный текст | Google Scholar
Бергманн, Б. М., Эверсон, К. А., Кушида, К. А., Фанг, В. С., Лейтч, К. А., Шоллер, Д. А. и др. (1989). Депривация сна у крысы: V. Использование энергии и посредничество. Сон 12, 31–41. DOI: 10.1093 / сон / 12.1.31
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бетли, Дж.Н., Цао Чжэнь Фанг, Х., Ритола, Кимберли, Д., Стернсон и Скотт, М. (2013). Организация параллельных резервных цепей для гомеостатического контроля поведения при кормлении. Ячейка 155, 1337–1350. DOI: 10.1016 / j.cell.2013.11.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бланко, М. Б., Даусманн, К. Х., Фаэрти, С. Л., Клопфер, П., Кристал, А. Д., Шоплер, Р. и др. (2016). Спячка у приматов: наступает ли сон? R. Soc. Open Sci. 3: 160282.DOI: 10.1098 / RSOS.160282
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Borbély, A. A. (1982). Двухпроцессная модель регуляции сна. Hum. Neurobiol. 1, 195–204.
Google Scholar
Боулант, Дж. А., и Гонсалес, Р. Р. (1977). Влияние температуры кожи на гипоталамический контроль потери тепла и производства тепла. Brain Res. 120, 367–372. DOI: 10.1016 / 0006-8993 (77)
-7
CrossRef Полный текст | Google Scholar
Бойер, Б.Б. и Барнс Б. М. (1999). Молекулярные и метаболические аспекты гибернации млекопитающих. Выражение фенотипа гибернации является результатом скоординированной регуляции множества физиологических и молекулярных событий во время подготовки к оцепенению и входа в него. Bioscience 49, 713–724. DOI: 10.2307 / 1313595
CrossRef Полный текст | Google Scholar
Бак, К. Л., и Барнс, Б. М. (2000). Влияние температуры окружающей среды на скорость метаболизма, респираторный коэффициент и торпор в арктической спячке. Am. J. Physiol. Regul. Интегр. Комп. Physiol. 279, R255 – R262. DOI: 10.1152 / ajpregu.2000.279.1.R255
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Баннелл Д. Э., Агнью Дж. А., Хорват С. М., Джопсон Л. и Уиллс М. (1988). Пассивное нагревание тела и сон: влияние близости ко сну. Сон 11, 210–219. DOI: 10.1093 / сон / 11.2.210
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кэмпбелл, С.С.и Бротон Р. Дж. (1994). Быстрое снижение температуры тела перед сном: взбить физиологическую подушку? Хронобиол. Int. 11, 126–131. DOI: 10.3109 / 0742052940
99
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кэри, Х. В., Эндрюс, М. Т., и Мартин, С. Л. (2003). Гибернация млекопитающих: клеточные и молекулярные реакции на угнетенный метаболизм и низкую температуру. Physiol. Ред. 83, 1153–1181. DOI: 10.1152 / Physrev.00008.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cespuglio, R., Amrouni, D., Meiller, A., Buguet, A., and Gautier-Sauvigné, S. (2012). Оксид азота в регуляции состояний сна и бодрствования. Sleep Med. Ред. 16, 265–279. DOI: 10.1016 / j.smrv.2012.01.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чоу, Т. К., Бьоркум, А. А., Гаус, С. Е., Лу, Дж., Скаммелл, Т. Е., и Сапер, К. Б. (2002). Аференты к вентролатеральному преоптическому ядру. J. Neurosci. 22, 977–990. DOI: 10.1523 / JNEUROSCI.22-03-00977.2002
CrossRef Полный текст | Google Scholar
Коули, М.А., Смарт, Дж. Л., Рубинштейн, М., Сердан, М. Г., Диано, С., Хорват, Т. Л. и др. (2001). Лептин активирует анорексигенные нейроны ПОМК через нейронную сеть в дугообразном ядре. Природа 411, 480–484. DOI: 10.1038 / 35078085
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чейслер, К.А., Циммерман, Дж.К., Ронда, Дж. М., Мур-Эде, М. К., и Вайцман, Е. Д. (1980). Время быстрого сна связано с циркадным ритмом температуры тела человека. Сон 2, 329–346. DOI: 10.1093 / сон / 2.3.329
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Даан, С., Барнс, Б. М., и Страйкстра, А. М. (1991). Разминка перед сном? Суслики спят во время пробуждения от спячки. Neurosci. Lett. 128, 265–268. DOI: 10.1016 / 0304-3940 (91)
-Y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
.