
Ячейки КСО — полезная информация
В данной статье хотим рассказать о наших ячейках КСО, а именно КСО- 366, 393, 307, 298 и новой разработке КСО-219. Почему именно эти ячейки, чем они отличаются друг от друга и в чем их преимущества.
Немного общей информации
Камеры сборные одностороннего обслуживания предназначены для приема и распределения электроэнергии трехфазного переменного тока частотой 50 Гц напряжением 6, 10 кВ в сетях с изолированной или заземленной нейтралью.
КСО применяются в составе распределительных устройств напряжением 6, 10 кВ при новом строительстве, расширении, реконструкции и техническом перевооружении следующих объектов:
- распределительных и трансформаторных подстанций городских электрических сетей;
- распределительных и трансформаторных подстанций объектов гражданского назначения и инфраструктуры;
- распределительных подстанций предприятий легкой промышленности;
- тяговых подстанций городского электрического транспорта и метрополитена.


В целом принцип работы КСО у всех производителей один и тот же. Разница может быть в названиях и в собственных доработках.
Раньше названия формировались по следующему признаку, например КСО-366 — трехсотая серия 66-го года разработки, КСО-298 двухсотая серия
98-го года разработки и т.д.
Детальная структура условного обозначения
Наши первые ячейки
В 2012 году мы производили только КСО-366. Это ячейка с разъединителями (РВЗ) и выключателями нагрузки (ВНА, ВНР). На сегодняшний день это самая простая конфигурация ячеек, которая обеспечивает выполнение требуемых задач на первичном уровне.
КСО-366, 2014 год
КСО-366, 2020 год
Но уже тогда рынок начал требовать более технологичное оборудование, которое обеспечит защиту не только самому изделию, но и обслуживающему персоналу. Для того чтобы отключать токи короткого замыкания требовался более технологичный выключатель.
Для того чтобы отключать токи короткого замыкания требовался более технологичный выключатель.
В советские времена эту задачу выполнял масляный выключатель. Со временем технологии развивались, появились вакуумные выключатели. Гашение дуги происходило в вакууме. Ячейки становились при этом компактнее, однако сохранялось самое главное требование — требование к безопасности обслуживания и защиты оборудования.
Рынок в Татарстане по средневольтному напряжению в целом был свободен, поэтому мы посчитали это направление весьма перспективным. Именно в 2012 году мы начали прорабатывать вопрос о производстве ячеек с вакуумными выключателями. Начались первые разработки в этом направлении. Так появились ячейки КСО-298, КСО-307 и КСО-393.
Откровенно говоря, первые шаги были очень непростыми – на тот момент в организации не было достаточно компетентных кадров, способных полноценно заняться реализацией этой задачи, да и в целом на рынке Татарстана не хватало конструкторов. В плане технического персонала рынок по среднему напряжению был скудный.
В плане технического персонала рынок по среднему напряжению был скудный.
Первая ячейка
Разработав своими силами конструктив ячейки КСО-298, детально изучив аппаратный состав и наполняемость, мы провели серьезную работу с представителями компании
Таврида Электрик по внедрению нового продукта в производство, в результате чего получили аккредитацию на производство ячеек на базе вакуумных выключателей BB/TEL их производства. Следует отметить, что
Таврида Электрик и по сегодняшний день является лидером рынка и одним из крупнейших поставщиков коммутационной аппаратуры среднего напряжения.
Успешное внедрение и последовавшая аккредитация от
Таврида Электрик позволили получить комплекс привилегий, специальные условия поставки, серьезные скидки и т.п. С этого и началось наше развитие в сфере производства ячеек с вакуумными выключателями.
В дальнейшем мы адаптировали конструктив наших ячеек практически под весь ассортимент присутствующих на рынке России вакуумных выключателей: VF-12 и VL-12 (Элтехника), ВВР (Росвакуум), VD 4 (ABB) и др.
Одним из важнейших достижений в развитии направления оборудования среднего напряжения стала совместная с компанией Schneider Electric проработка адаптации наших ячеек под вакуумные выключатели EasyPact EXE. Комбинация EasyPact EXE с релейной защитой SEPAM в наших ячейках стала одним из наиболее удачных и конкурентоспособных решений, что в дальнейшем позволило нам, в том числе, стать авторизованным сборщиком вакуумных выключателей компании Schneider Electric и получить специальный сертификат.
4 линейки как стандарт для завода
Проведя анализ рынка, изучив самые распространенные ячейки, их габаритные размеры, целевой рынок, для кого, куда и каким образом поставлять ячейки, мы остановились на 4-х типах:
КСО-366 – типовые ячейки на базе выключателей нагрузки ВНА, ВНР и разъединителей РВЗ;
КСО-393 – усовершенствованная КСО-366, с возможностью установки также и вакуумных выключателей;
КСО-307 – современная компактная ячейка (шириной от 400 мм) с элегазовыми и вакуумными аппаратами продольного исполнения;
КСО-298 – стандартная ячейка с вакуумными выключателями с локализацией по отсекам.
КСО-393, 2019 год
КСО-298, 2019 год
«5 элемент» или наша новая разработка КСО-219
В 2018 году мы снова изучили рынок и сделали вывод, что рынку нужна малогабаритная ячейка с высокой степенью локализации по всем отсекам, т.к. существовавшие ячейки не давали полной локализации для максимально безопасной эксплуатации оборудования. В основу нашей КСО-219 вошел принцип ячейки КРУ, где разделение идет по 4 отсекам: отсек вакуумного выключателя, отсек кабельных присоединений, отсек релейной защиты и отсек шин, которые 100% локализованы друг от друга, но при этом ячейка имеет уменьшенный габаритный размер.
Основные преимущества ячейки КСО-219 — уменьшенный габаритный размер в сочетании с лучшей ценой на рынке. Ее легко можно поставить как в существующую подстанцию, так и в подстанцию из бетона или утепленных сэндвич — панелей. За счет достаточно компактных габаритных размеров она полностью удовлетворяет последним тенденциям рынка, характеризующихся общим уменьшением габаритных размеров и компактностью изделий. Наши заказчики подтверждают, что такая ячейка действительно была нужна на рынке.
Наши заказчики подтверждают, что такая ячейка действительно была нужна на рынке.
Как итог, с освоением и внедрением в производство ячеек КСО-219 нам удалось сформировать именно такой модельный ряд ячеек КСО, который позволяет обеспечить все имеющиеся потребности заказчиков.
КСО-219 на форуме «Электрческие сети» в Москве, 2019 год
Камеры КСО и Ячейки КСО / Расшифровка, производство, цены
Расшифровка аббревиатуры КСО – камеры сборные одностороннего обслуживания, предназначенные для осуществления подачи напряжения на промышленных объектах.
Назначение
КСО – это камеры сборные одностороннего обслуживания. КСО являются камерами, предназначенными для осуществления приема и перераспределения электрической энергии трехфазного тока с напряжением до 10 киловольт и технической частотой 50 Гц. Эксплуатация камеры сборной (КСО) допускается в сети имеющей изолированную нейтраль.
КСО с вакуумными выключателями предназначены для эксплуатации в народном хозяйстве в умеренноклиматическом поясе, при температурах окружающего воздуха варьирующих в интервале от -25 до +40 градусов Цельсия. Изготовление камер осуществляется в соответствии с разработанными типовыми проектами. Внутри камер размещается соответствующая аппаратура управления и шины главных цепей. На фасаде камеры размещаются приводы управления выключателями нагрузки.
Компоновка
В соответствии со схемами первичных соединений камеры могут комплектоваться различным оборудованием. Среди разнообразного оборудования присутствуют автогазовые выключатели нагрузки типа ВН-РА имеющие пружинные приводы, разъединители типа РВ имеющие заземляющие ножи, вакуумные выключатели типа BB/TEL, BBEL имеющие электромагнитные приводы.
В состав КСО входит трансформатор напряжения, и трансформатор для собственных нужд. Автоматика и защита на основе реле выполняется с применением статических реле, помимо этого автоматика может быть выполнена на основе микропроцессорных устройств.
Варианты производства
КСО с выключателями вакуумной конструкции производятся в двух вариациях. Производится вариант со стационарным размещением выключателя вакуумной конструкции, а также выдвижное расположение. Выдвижное расположение осуществляется путем использования специальных кассет в которых применяются выключатели с диаметром 24 и 36 мм.
В соответствии с требованиями заказчика установка может комплектоваться панелями для собственных нужд НН, на которых монтируются автоматические выключатели отходящих линий.
КСО содержат в своем устройстве счетчики реактивной и активной электрической энергии.При оборудовании КСО счетчиками электрической энергии могут применяться многофункциональные устройства которые относятся к счетчикам типа ЩО-94-2401 и ЩО-94-2402.
КСО состоит из раздельных ячеек, которые снабжаются встроенными аппаратами измерения и сигнализации. В зависимости от используемой аппаратуры и присоединений зависит конфигурация камеры.
Ток при шинном вводе подключается посредством использования изоляторов проходного типа. Изоляторы размещаются на задней или верхней стенке. Боковые отводы осуществляются при помощи проходных изоляторов.
Изоляторы размещаются на задней или верхней стенке. Боковые отводы осуществляются при помощи проходных изоляторов.
Ввод при помощи кабеля в КСО делается посредством специальных кабельных каналов, располагающиеся внизу камеры с последующим присоединением внутри.
Камера дает возможность подключать две линии высокого напряжения при сечении кабеля 3х240 кв. мм или трех кабелей с одной токонесущей жилой имеющей сечение до 630 кв.мм.
Установка
Установка камеры проводится путем монтажа КСО на специальном фундаменте. КСО предназначается для осуществления одностороннего обслуживания.
Компоновка камер обладает удобством осмотра и проведения ремонтов в процессе эксплуатации КСО. При этом можно не проводить снятие напряжения с токоведущих шин. В фасаде имеются специальные дверки для облегчения доступа при обслуживании к оборудованию КСО. Производство КСО осуществляется специализированными заводами-изготовителями, которые специализируются на производстве электрооборудования. КСО имеет цену в зависимости от комплектации, которую заказывает заказчик производителю.
КСО имеет цену в зависимости от комплектации, которую заказывает заказчик производителю.
Принцип работы ячеек КСО
Чтобы понять принцип работы ячеек КСО обратимся к истории ее создания. С развитием в СССР в 60-е годы электроэнергетики появилась необходимость унификации и снижения стоимости большого количества распределительных устройств напряжением 6 и 10 кВ. Для повсеместно распространяющихся комплектных трансформаторных подстанций требовались малогабаритные унифицированные ячейки (камеры), выполняющие элементарные функции в закрытых распределительных устройствах. Такими решениями стали камеры с двухсторонним (КРУ) и односторонним обслуживанием (КСО).
Принцип работы КСО
Внутри ячеек располагается оборудование и устройства для приема и распределения электроэнергии. Производители предлагают ряд типовых схем ее главных (первичных) и вспомогательных (вторичных) внутренних цепей. Чаще всего это: подключение воздушной линии или кабельного ввода, заземление сборных шин, шинный мост, кабельные сборки, подключение трансформатора собственных нужд и другие.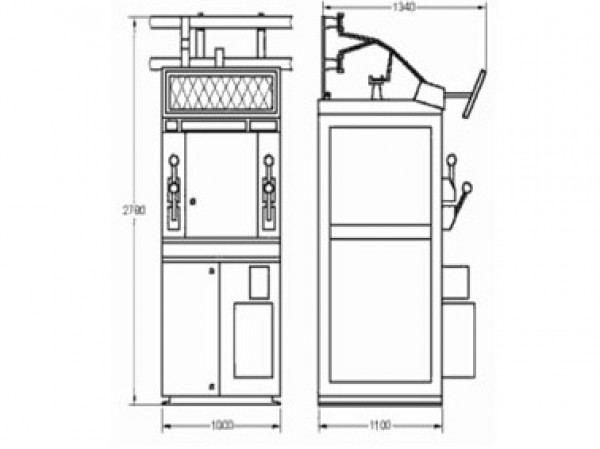 Возможно изготовление ячеек по индивидуальным схемам Заказчика.
Возможно изготовление ячеек по индивидуальным схемам Заказчика.
Состав ячейки
Ячейки представляют собой сварную металлоконструкцию (камеру). Внутри нее располагается оборудование главных цепей. Состав этого оборудования и схема его соединения определяется назначением ячейки.
В корпусе ячейки может быть установлено следующее оборудование: высоковольтные выключатели (в том числе выключатели нагрузки), разъединители, измерительные и силовые трансформаторы, ограничители перенапряжения, предохранители, указатели напряжения и другое оборудование различных производителей.
Также внутри ячейки устанавливается ящик с оборудованием вторичных цепей: механические или микропроцессорные реле, автоматами, приборами учета и измерения электроэнергии и т.п.
На фасад камеры вынесены органы управления коммутационными аппаратами и аппаратура вторичных цепей.
Для безопасной эксплуатации ячейки предусмотрена система механических и электрических блокировок от неправильных оперативных действий персонала.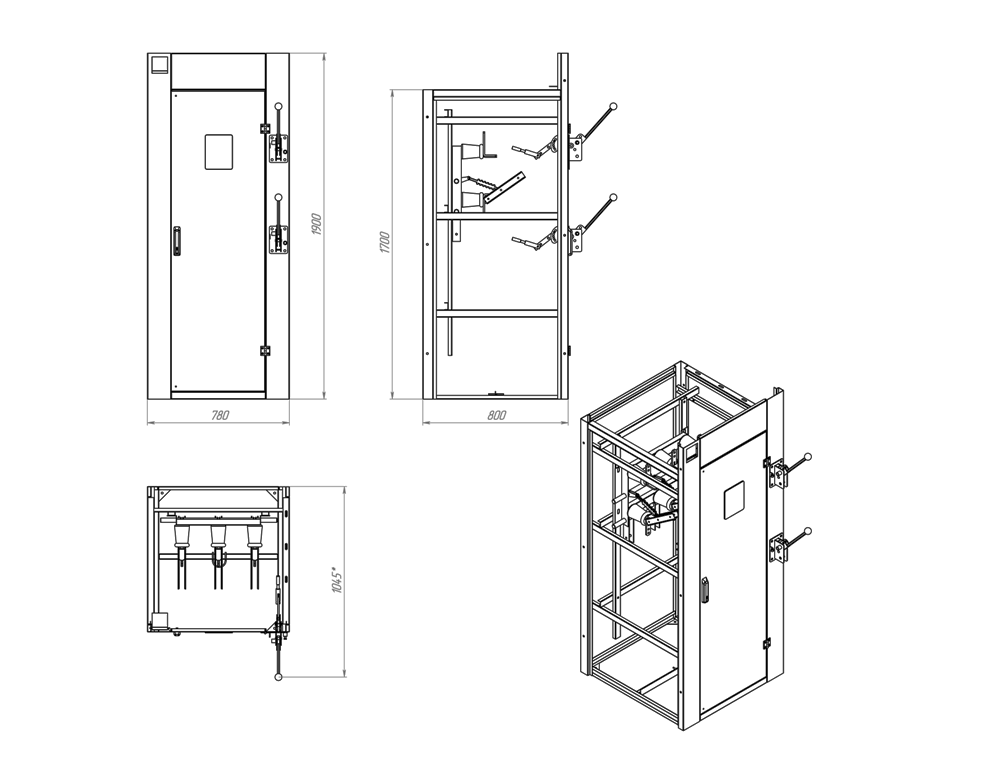 Имеются сетчатые ограждения открытых токоведущих частей ячейки, разделительные перегородки и другие защитные элементы.
Имеются сетчатые ограждения открытых токоведущих частей ячейки, разделительные перегородки и другие защитные элементы.
Сборные алюминиевые шины располагаются в верхней части камер, там же имеется короб с магистралями вспомогательных цепей и ряды зажимов. Зона кабельных подключений располагается внизу камер. Доступ в камеру осуществляется через дверь, имеющей замок и смотровое окно, предусмотрено освещение внутреннего оборудования ячейки.
Возможно как одно-, так и двухрядное размещение камер. Для соединения рядов используются шинные мосты с разъединителями.
Изоляция
В качестве естественного изолятора между токоведущими поверхностями и заземленными частями камер используется воздушный промежуток. Расстояния между элементами ячейки регламентируются ПУЭ. Такое решение несколько увеличивает габариты ячейки, но позволяет снизить ее стоимость.
Таким образом, принцип работы ячеек КСО определяется их назначением в схеме распределительного устройства, а значит схемами первичных и вторичных цепей.
Ячейки КСО (272-298) — ЗАВОД ЭНЕРГОСНАБ
КСО (камера одностороннего обслуживания) – один из видов распределительных устройств, представляющий собой шкафы со встроенными защитными элементами. Каждый конкретный шкаф предусматривает наличие вмонтированных в него измерительных, защитных, коммутационных аппаратов и называется ячейкой КСО.
Для чего нужны КСО?
Это оборудование применяется как в трансформаторных подстанциях с номинальной частотой тока 50 Гц и напряжением 6 кВ, так и в других аппаратах, имеющих схожие характеристики. Если устройства предусматривают наличие заземленной или изолированной нейтрали, напряжение на них может достигать 10 кВ. Производство КСО необходимо для промышленных и сельскохозяйственных нужд.
Ячейка КСО: условия эксплуатации
Для того, чтобы камеры одностороннего обслуживания исправно работали на протяжении долгого времени, необходимо учитывать следующие рекомендации:
- Эксплуатировать ячейку можно при определенной температуре: от -25 о С до +35 о С.
- Высота расположения устройства не должна превышать километра над уровнем моря.
- Если использовать камеры в закрытых помещениях, необходимо заранее убедиться, что в воздухе отсутствуют агрессивные газы и токопроводящая пыль, которые могут стать причиной повреждения металлической и изоляционной части КСО.

Размеры ячеек
Стандартный вариант этого устройства предусматривает:
- ширину от 80 до 100 см;
- глубину от 80 до 105 см.
Уточнить размеры конкретной модели ячейки можно при заказе.
Технические характеристики ячейки КСО-272
Номинальное напряжение – 6,0 или 10,0 кВ.
Рабочее напряжение – 7,2 или 12 кВ.
Номинальный ток:
- сборных шин – 1600 А;
- главных цепей – 630 или 1000 А.
Ток электродинамической стойкости – 51 кА.
Ток термической стойкости (2 с) – 20 кА.
Номинальный ток отключения вакуумных выключателей – 12,5 или 20 кА.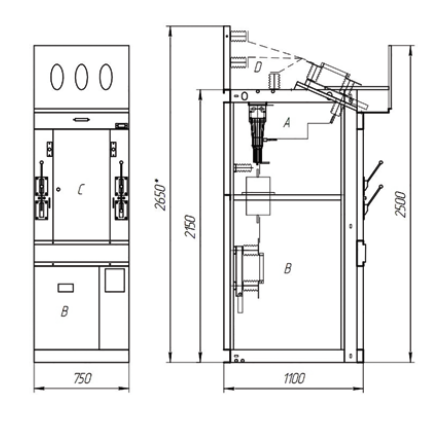
Номинальный ток плавкой вставки предохранителей – 4, 6,3, 10, 16, 20, 25, 31,5, 40, 50, 63, 80, 100, 160, 200 А.
Номинальный ток трансформаторов тока – от 75 до 1500 А.
Номинальное напряжение вспомогательных цепей:
- переменного тока – 110 или 220 В;
- постоянного тока – 110 или 220 В;
- освещения (ток переменный) — 36 В.
Класс защиты оболочки — IP31.
Габаритные размеры:
- ширина – от 80 до 100 см;
- глубина – от 80 до 105 см;
- высота – 216, 236, 195 см.
Масса ячеек не более:
- с силовыми выключателями – 490 кг;
- с разъединителями и выключателями нагрузки – 240 кг;
- с трансформатором собственных нужд – 570 кг.
Характеристики конкретных моделей ячеек КСО можно найти на соответствующих страницах.
Как обеспечивается безопасность КСО?
- Камера разделена на несколько изолированных друг от друга блоков, что способствует безопасности работы.
- На случай возникновения перенапряжений и коротких замыканий на задней панели ячейки трансформаторной подстанции предусмотрены разгрузочные отверстия, через которые выводятся продукты горения.
- Некорректные операции блокируются с помощью специальной системы.
- Заземлены все металлические элементы устройства, которые могут находиться под напряжением.

Цена на ячейки КСО 272-298 договорная, стоимость зависит от таких параметров, как комплектация и модель устройства.
Камеры сборные одностороннего обслуживания КСО
Ячейки КСО представляют собой одну из разновидностей распределительных электроустановок с односторонним обслуживанием. Камеры КСО предназначены для дальнейшей сборки распределительных устройств, необходимых для приема и передачи электрической энергии. Ячейка КСО – это сварная металлоконструкция из листовых профилей, внутри которой находятся устройства управления, аппаратура основных цепей, рукоятки приводов, реле управления и предохранения. На передней стороне камеры расположены измерительные и учетные приборы.Камера КСО оборудована двумя дверями для доступа к трансформатору напряжения, высоковольтному выключателю и предохранителю, а также к кабельным разъединителям силового трансформатора. Высоковольтный выключатель и приборы вспомогательных цепей ради безопасности отгорожены съемной перегородкой. Обозревать внутреннее пространство камеры можно через смотровые окна.
На передней стороне камеры расположены измерительные и учетные приборы.Камера КСО оборудована двумя дверями для доступа к трансформатору напряжения, высоковольтному выключателю и предохранителю, а также к кабельным разъединителям силового трансформатора. Высоковольтный выключатель и приборы вспомогательных цепей ради безопасности отгорожены съемной перегородкой. Обозревать внутреннее пространство камеры можно через смотровые окна.
Кроме того, ячейки КСО оборудованы специальным устройством, которое позволяет произвести замену перегоревшей лампы без необходимости снимать напряжение. Все устройства и приборы КСО заземлены. Приборы вспомогательных цепей, расположенные за верхней дверью, заземлены эластичным проводом. Каркас ячейки монтируется на заземленные металлические конструкции.
Характеристики КСО
Стандартные размеры ячеек КСО составляют 75 сантиметров в ширину и 80-105 сантиметров в глубину. Уточнить точные размеры камеры можно во время заказа оборудования.
КСО характеризуется следующими параметрами:
- Рабочее напряжение варьируется в пределах 12 – 7,2 кВ;
- Наибольшее рабочее напряжение – 10(6) кВ;
- Наибольшее рабочее напряжение основных цепей: 1000, 630 и 400 А;
- Электроток сборных шин – 630(1000) А;
КСО оборудуются разными электромагнитными и механическими предохранителями, которые блокируют ошибки в действиях персонала во время оперативных переключений. Все электромеханические реле и микропроцессорные блоки КСО защищены и автоматизированы. Если на распределительном устройстве есть два ввода (рабочий и резервный), возможно автоматический запуск резервного питания.
Все электромеханические реле и микропроцессорные блоки КСО защищены и автоматизированы. Если на распределительном устройстве есть два ввода (рабочий и резервный), возможно автоматический запуск резервного питания.
Назначение КСО
Из камер КСО комплектуются распределительные установки с наивысшей рабочей частотой электротока в 50Гц и напряжением в 6 кВ либо в 10 кВ для устройств с изолированной либо заземленной нейтралью. Ячейки КСО находят применение в промышленных и бытовых нуждах, при строительстве, реконструкции или техническом переоснащении городских, промышленных, транспортных распределительных и трансформаторных подстанций. Кроме того, ячейки КСО используются в качестве УВН (установки высшего напряжения) для КТП мощностью 100 – 2500 кВА.
Условия эксплуатации КСО
Для надежного, бесперебойного и максимально длительного функционирования ячеек КСО необходимо придерживаться следующих условий эксплуатации:
- Устанавливать камеры КСО в местности, расположенной не выше тысячи метров над уровнем моря;
- Камеры КСО хорошо функционируют в умеренном климате, при минимальных температурах не ниже -25 по Цельсию и максимальных – не выше +35 по Цельсию;
- При использовании ячеек в закрытых помещениях, следует избегать взрывоопасной и пожароопасной среды, газов, агрессивных сред и токопроводящей пыли.

Наши камеры КСО состоят из отдельных изолированных блоков, что повышает безопасность их эксплуатации. В случае возникновения непредвиденного короткого замыкания внутри металлоконструкции, все продукты горения удаляются через разгрузочные отверстия, которыми оборудованы задние ячейки. Грамотная система блокировок защищает аппаратуру от возможных ошибочных действий рабочего персонала во время обслуживания камеры.
Опросный лист КСО
Камеры КСО предназначены для работы в электрических установках трехфазного переменного тока частотой 50 и 60 Гц напряжением 6 и 10 кВ в системах с изолированной или заземленной через дугогасящий реактор нейтралью.
Из камер КСО собираются распределительные устройства, служащие для приема и распределения электроэнергии. Климатическое исполнение У и Т, категория размещения 3 по ГОСТ 15150 и ГОСТ 15543.
Распределительное устройство содержит набор коммутационных аппаратов, сборные и соединительные шины, вспомогательные устройства РЗиА и средства учёта и измерения.
Область применения
Комплектные распределительные устройства могут использоваться как для внутренней, так и для наружной установки (в этом случае их называют КРУН). КРУ широко применяются в тех случаях, где необходимо компактное размещение распределительного устройства. В частности, КРУ применяют на электрических станциях, городских подстанциях, для питания объектов нефтяной промышленности (нефтепроводы, буровые установки), в схемах энергопотребления судов.
Если основное оборудование КРУ заключено в оболочку, заполненную элегазом, то РУ сокращённо обозначают КРУЭ.
КРУ, у которого все аппараты размещены в одном отсеке, называется камерой сборной одностороннего обслуживания (КСО).
Как правило, шкаф КРУ разделён на 4 основных отсека: 3 высоковольтных — кабельный отсек (ввода или линии), отсек выключателя и отсек сборных шин и 1 низковольтный — релейный шкаф.
|
| |||||||||||||||||||
 Как правило, КСО действительно одностороннего обслуживания, чаще всего имеет открытые сборные шины, задняя стенка отсутствует.
Как правило, КСО действительно одностороннего обслуживания, чаще всего имеет открытые сборные шины, задняя стенка отсутствует.




Развитие ячеек КСО
Камеры сборные одностороннего обслуживания (ячейки или камеры КСО) в настоящее время являются одним из наиболее популярных видов распределительных устройств 6-10 кВ.
Между тем, в технической литературе мы не найдем упоминания о них до начала 90-х годов, хотя первые конструкции ячеек КСО относятся к началу 60-х годов прошлого века.
Как же происходило развитие ячеек КСО?
В середине 60-х годов ХХ века в связи с бурным развитием энергетики, строительством каскадов ГЭС в Сибири и началом формирования Единой Энергосистемы (ЕЭС) остро встал вопрос унификации и снижения стоимости распределительного оборудования среднего напряжения (6-10 кВ). Тогда родилась концепция комплектных трансформаторных подстанций (КТП), которая сейчас (правда, уже по другим причинам) стала почти доминирующей.
Комплектные трансформаторные подстанции должны были состоять из унифицированных ячеек, выполняющих набор стандартных функций. На среднее напряжение такими ячейками должны были стать камеры КСО-2, а на низкое напряжение 0,4 кВ — панели ЩО-70.
Но, как это часто бывает, на начальном этапе развитие прогрессивной концепции сдерживалось из-за отсутствия необходимой элементной базы.
Распределительные устройства в эпоху масляных выключателей
Основным коммутирующим устройством среднего напряжения в то время были масляные выключатели. Учитывая, что основные распределительные устройства 6-10 кВ проектировались на достаточно большие токи 1000-2000 А, габаритные размеры вакуумных выключателей получались довольно значительными.
Ширина самого масляного выключателя составляла 900 мм. Однако большой вес выключателя (300 кг) приводил к необходимости установки его на выкатную тележку. В результате ширина шкафа КРУ с выкатным масляным выключателем достигала 1360 мм, а глубина — 1650 мм при высоте 2850 мм. Понятно, что при таких габаритах обслуживание шкафов КРУ могло быть только двусторонним.
Для уменьшения габаритов распределительных устройств была разработана ячейка КСО-2. Ее принципиальным отличием от КРУ была малая глубина и возможность одностороннего обслуживания (что отражено в самом названии). Габариты ячейки КСО-2 составляли (ШхГхВ) 1000х1200х2800.
Чтобы уменьшить глубину камеры пришлось пожертвовать выкатной тележкой. Вакуумный выключатель стал стационарным. Это сразу породило проблему видимого разрыва.
По Правилам устройства электроустановок (ПУЭ) на время проведения ремонтных работ должен быть обеспечен видимый разрыв в электропитании. Когда тележка масляного выключателя выкатывалась, контакты автоматически размыкались, создавая тем самым требуемый видимый разрыв. Для обеспечения видимого разрыва в КСО с масляным выключателем пришлось добавить два разъединителя — шинный и линейный. Для отключения масляного выключателя использовался пружинный привод. В результате появилась ячейка КСО-2, которая стала прародительницей всех нынешних ячеек КСО-2хх (второй серии).
Из соображений безопасности сначала добавили заземляющие ножи линейного разъединителя, в результате появилась ячейка КСО-266, а затем и заземляющие ножи шинного разъединителя — ячейка КСО-272 (последние две цифры — год появления модели).
| Ячейка КСО-272 (a — фасад, б — разрез): 1 и 5 — приводы шинного и линейного разъединителей, 2 — приводы заземляющих ножей, 3 — защитное ограждение, 4 — световой карниз, 6 — пружинный привод ППВ-10, 7 и 11 — шинный и линейный разъединители, 8 — заземляющие ножи, 9 — масляный выключатель ВМГП-10, 10 — трансформаторы тока. |
Поскольку габариты ячеек все еще оставались довольно большими, помимо КСО-2, были разработаны также ячейки КСО-3 с уменьшенными габаритами (ШхГхВ) 1000х1000х1860 для размещения оборудования, которое не требовало таких больших объемов, как масляные выключатели. В этих ячейках размещались: выключатели нагрузки, разъединители, предохранители, трансформаторы напряжения, разрядники.
Впоследствии, аналогично ячейкам КСО-266, были добавлены стационарные заземляющие ножи, что привело к появлению ячеек КСО-366.
Ячейки КСО-266 на масляных выключателях с пружинным приводом
производства 70-х годов XX века (в рабочем состоянии)
Ячейки КСО на выключателях нагрузки
Рыночные реформы 90-х годов сыграли значительную роль в развитии ячеек КСО. На рынок вышло большое количество небольших самостоятельных потребителей электроэнергии. Их требования к номинальным потребляемым токам были гораздо ниже, чем у крупных промышленных предприятий эпохи Советского Союза. В результате на рынке стали доминировать комплектные трансформаторные подстанции мощностью до 630 кВА. Номинальные токи по стороне высокого напряжения (с учетом перехода напряжения с 6 кВ на 10 кВ) снизились с 1000 А до 100 А. Для работы в этих условиях вполне хватало выключателей нагрузки типа ВНА-10.
Для удовлетворения потребности в недорогом и сравнительно маломощном распределительном оборудовании уже не требовались серьезные производственные мощности. Распределительные устройства можно было изготавливать даже в кустарных условиях. На рынок электротехнического оборудования обрушилась лавина малых предприятий (с количеством работников менее 50 человек), которые стали выпускать распределительные устройства (и в частности, ячейки КСО) по собственным техническим условиям. Часто одно и то же изделие у разных производителей имело разные наименования (КСО-266, КСО-272, КСО-285, КСО-292, КСО-298, КСО-366, КСО-393 с различными индесами по названию производителя). Как следствие, номенклатура ячеек КСО увеличилась в десятки раз.
Среднее напряжение класса 6-10 кВ практически не используется конечными потребителями без преобразования (исключение составляют, пожалуй, только электродвигатели на насосных станциях). По этой причине большинство устройств среднего напряжения являются распределительными, а камеры КСО, как элементарные ячейки распределительных устройств выполняют ограниченный набор функций.
По своему назначению наиболее распространенными являются следующие типы ячеек: линейная, трансформаторная и секционная. В соответствии с названиями, линейные ячейки обеспечивают коммутацию и защиту входящих и отходящих линий, трансформаторные — подключение и защиту силовых трансформаторов, секционные ячейки — управляют вводом резерва. Кроме того, часто присутствуют вспомогательные ячейки, в которые, например, устанавливаются трансформаторы собственных нужд (ТСН).
Такая ограниченность функций позволила унифицировать однолинейные схемы ячеек КСО и создать альбомы типовых схем первичных соединений. По сути все схемы отличаются только наличием и расположением предохранителей и трансформаторов. Выбор конкретной схемы в том или ином случае определяется проектировщиком на основе требований к безопасности и личных предпочтений.
Логика наполнения ячеек элементами достаточно проста:
- На каждой линии 6-10 кВ должен быть выключатель нагрузки с заземляющими ножами для отключения линии
- Последовательно с выключателем нагрузки должен быть включен разъединитель для организации видимого разрыва цепи при проведении ремонтных и регламентых работ
- Аналогичное требование должно выполняться для линии, отходящей на трансформатор
- Если в системе предусмотрено секционирование для ручного ввода резерва, то между двумя секциями распределительного устройства должен находиться разъединитель для коммутации и организации видимого разрыва. Поскольку ввод резерва, как правило, осуществляется при отключенной линии, то наличие выключателя нагрузки в этом случае не обязательно.
- Если необходимо защитить трансформатор от коротких замыканий на линии или защитить линию от коротких замыканий в самом распределительном устройстве, то в соответствующие ячейки добавляются предохранители.
- Если требуется учет электроэнергии на линии 6-10 кВ, то в соответствующие ячейки устанавливаются трансформаторы тока (ТТ) и трансформаторы напряжения (ТН) для передачи сигнала на счетчик электроэнергии.
Используя приведенные выше правила можно легко разобраться в однолинейной схеме распределительного устройства, а также понять назначение органов управления ячеек КСО.
Камеры КСО-393 на выключателях нагрузки
Кроме функции коммутации и защиты линии и трансформаторов (с помощью предохранителей), ячейки КСО выполняют важную функцию защиты обслуживающего персонала от поражения электрическим током. Для этого в ячейках КСО предусмотрен целый ряд блокировок.
Состав блокировок регламентируется межгосударственным стандартом ГОСТ 12.2.007.4-75 «Система стандартов безопасности труда. Шкафы комплектных распределительных устройств и комплектных трансформаторных подстанций, камеры сборные одностороннего обслуживания, ячейки герметизированных элегазовых распределительных устройств».
Камеры КСО должны быть оборудованы заземляющими ножами, если это указано в стандартах или технических условиях на конкретные виды изделий.
Основные типы блокировок КСО:
- блокировка, не допускающая включения или отключения разъединителей при включенном выключателе первичной цепи;
- блокировка между разъединителем и ножами заземления, не допускающая включения разъединителей при включенных ножах заземления либо включения ножей заземления при включенных разъединителях;
- блокировка стационарных разъединителей с дверями или сетчатыми ограждениями, выполненными в виде дверей, не допускающая открывания дверей при включенных разъединителях;
- блокировка, не допускающая включения заземляющего разъединителя, при условии, что в других камерах КСО, от которых возможна подача напряжения на участок главной цепи камеры, где размещен заземляющий разъединитель, коммутационные аппараты находятся во включенном положении;
- блокировка, не допускающая при включенном положении заземляющего разъединителя, включения любых коммутационных аппаратов в других камерах КСО, от которых возможна подача напряжения на участок главной цепи камеры, где размещен заземляющий разъединитель.
В камерах КСО, которые снабжены заземляющими разъединителями, должна быть предусмотрена возможность запирания привода заземляющего разъединителя при включенных ножах при помощи замка.
Впервые столкнувшихся с ячейками КСО всегда удивляет наличие большого свободного пространства внутри ячеек, что кажется очень нерациональным. Однако, это является прямым следствием того, что в качестве изолятора используется воздушный промежуток. Расстояния между токоведущими поверхностями и от токоведущих поверхностей до заземленных проводников жестко регламентируются Правилами устройства электроустановок (ПУЭ). Эти правила многократно проверены практикой. Встречались случаи, когда в весенний и осенний период, при повышенной влажности воздуха, уменьшение расстояния между проводниками всего на 1 см приводило к межфазному пробою.
Революция вакуумных выключателей
Массовое появление на рынке вакуумных выключателей, которые по коммутирующей способности не уступали масляным выключателям, но при этом имели в несколько раз меньшие габариты, произвело революцию в распределительных устройствах. Ячейки КСО второй серии КСО-2хх, а затем и третьей серии КСО-3хх стали активно переделывать под вакуумные выключатели…
Читайте далее:
- Ячейки КСО на вакуумных выключателях
- «Выкатные» ячейки на новом этапе
- «Авторские« и импортные ячейки
- Замена КСО на элегазовые моноблоки
Т-клеток не требуются для CSR
B-клетки могут быть стимулированы для прохождения ДНК-события, называемого рекомбинацией с переключением классов (CSR), которое позволяет B-клетке изменять константный хвост тяжелой цепи продуцируемых ею антител. Изотип антитела [иммуноглобулин M (IgM), IgG, IgE или IgA] определяется этим хвостом, который связывается с рецепторами на клетках-мишенях, чтобы инициировать различные клеточные ответы на один и тот же антиген. Литинский и др. описывает механизм CSR, который не требует взаимодействия Т-ячейка-В-ячейка.Культивируемые B-клетки, стимулированные экзогенным BLyS или APRIL, подвергались CSR, что обнаруживается небольшими количествами ДНК переключателя, внехромосомного продукта CSR, и экспрессией транскриптов из этого события. Добавление BLyS или APRIL с интерлейкином (IL) -10, IL-4 или трансформирующим фактором роста-β (TGF-β) усиливало CSR. BLyS и APRIL стимулировали активность фактора транскрипции NF-κB. BLyS или APRIL стимулировали повышенную секрецию антител IgA и IgG. Авторы определили, что интерфероны, лиганд CD40 и бактериальный липополисахарид стимулировали продукцию BLyS и APRIL в дендритных клетках или моноцитах, что способствовало CSR в B-клетках.CSR в ответ на активированные дендритные клетки блокировался обработками, которые нейтрализовали действие BLyS или APRIL. Когда рецептор В-клеток был активирован и клетки подвергались действию активированных антигенпрезентирующих клеток, тогда IgA и IgG секретировались посредством процесса, который требовал BLyS и APRIL. Таким образом, одни антигенпрезентирующие клетки могут быть способны контролировать продукцию антител к В-клеткам, не требуя Т-клеточного промежуточного звена. Макферсон и Ламар обсуждают актуальность этих результатов для аутоиммунных заболеваний.
А. Дж. Макферсон, А. Ламар, BLySful взаимодействия между DC и B-клетками. Нац. Иммунол. 3 , 798-800 (2002). [Интернет-журнал]
М. Б. Литинский, Б. Нарделли, Д. М. Гильберт, Б. Хе, А. Шаффер, П. Казали, А. Черутти, ДК индуцируют CD40-независимое переключение класса иммуноглобулинов через BLys и APRIL. Нац. Иммунол. 3 , 822-829 (2002). [Интернет-журнал]
Существенная роль исходного сигнала активации в выборе изотипа при делеции коммитированного транскрипцией промотора
CRISPR / Cas9-опосредованная делеция промотора Iα в линии клеток Ch22F3-2
Линия клеток мыши Ch22 происходит от Ch22.Линия клеток лимфомы LX. Эта клеточная линия транскрипционно коммитируется в промотор Iα, который обладает базовой активностью даже в отсутствие стимуляции, а активированные клетки Ch22 переключаются исключительно на IgA 20,21 . На протяжении всего этого исследования мы использовали субклон Ch22F3-2 22 (далее называемый линией или клетками Ch22).
Чтобы исследовать влияние удаления коммитированного промотора Iα на активацию вышестоящих промоторов I и CSR, мы разработали две направляющие РНК CRISPR / Cas9, специфически нацеленные на промотор / экзон Iα (рис.1А). Поскольку в Ch22 непродуктивный аллель уже подвергся рекомбинации Sµ / Sα 22,23 (дополнительный рис. S1A), в результате чего удалены все промоторы I, кроме энхансера / промотора Eµ / Iµ (рис. 1A), gRNAs нацелены исключительно на продуктивный аллель. ПЦР-скрининг и секвенирование выявили восемь клонов с желаемой делецией (дополнительный рисунок S1A – D).
Рисунок 1
Делеция промотора / экзона Iα ингибирует CSR к IgA. ( A ) Схематическая структура локуса IgH в линии Ch22F3-2.Неэкспрессируемый аллель представляет собой частично реаранжированный аллель DJ H , который подвергся рекомбинации Sµ / Sα, в результате чего были удалены все индуцибельные промоторы I, расположенные выше. Сверху указаны митоген и цитокины, индуцирующие различные промоторы I. Сайты, фланкирующие промотор / экзон Iα, на которые нацелены гРНК, указаны стрелками. Показаны энхансер / промотор Eµ / Iµ между вариабельной и константной областями и суперэнхансер 3’RR ниже локуса. ( B ) Анализ проточной цитометрии клонов с делецией Iα.8 клонов, полученных с помощью CRIPR / Cas9, анализировали с помощью FACS на предмет поверхностной экспрессии IgA. Родительскую линию Ch22 использовали в качестве контроля до (UNS) и после стимуляции LIT (LPS + IL4 + TGFβ). LIT-активированные В-клетки селезенки также были включены в качестве контроля (n = 3). ( C ) Типичный график FACS, полученный с клетками Ch22 и клоном с делецией Iα (№1) до и после стимуляции LIT.
Анализ FACS показал, что в ответ на LIT (LPS + IL4 + TGFβ) клетки Ch22 претерпевают устойчивый CSR к IgA до уровней выше, чем в активированных B-клетках селезенки (рис.1Б). Как и ожидалось, ни один из клонов с делецией Iα не переключается на IgA (рис. 1B, C). Мы проверили на трех случайных мутантных клонах, что не произошло никакого сплайсинга trans между экзоном VDJ продуктивного аллеля и областью Cα непродуктивного аллеля (дополнительный рисунок S1E).
Переключение транскрипции и CSR в активированных клетках Ch22 и делеционных клонах
Чтобы определить, возникает ли CSR в отсутствие коммитированного промотора Iα, мы сначала протестировали переключение в условиях стимуляции, которые, как известно, вызывают переключение в первичных B-клетках; Стимуляция LPS индуцирует CSR к IgG3 и IgG2b, LPS + IL4 к IgG1 и IgE, LPS + IFNγ к IgG2a и LIT к IgG2b и IgA.
Анализ FACS показал, что, в отличие от активированных В-клеток селезенки, в Ch22, а также во всех клонах с делецией Iα, LPS, LPS + IFNγ и LPS + IL4 не индуцируют CSR к IgG3, IgG2a и IgG1 соответственно. Ни один из этих стимулов не индуцировал CSR для IgA, как ожидалось (рис. 2A – C, дополнительные рис. S2 и S3). Эти результаты были подтверждены в трех случайных клонах путем количественной оценки RT-qPCR транскриптов после переключения 24 (дополнительный рисунок S4).
Рисунок 2
клонов с делецией Iα неспособны подвергнуться CSR после специфической стимуляции.( A — C ) Клетки Ch22, три клона с делецией Iα и В-клетки селезенки были активированы LPS ( A) , LPS + IFNγ ( B ) или LPS + IL4 ( C ) и окрашены на IgG3, IgG2a и IgG1 соответственно. Представительные графики показаны для нестимулированных (UNS) и активированных клеток Ch22, клонов с делецией Iα (клон 5) и первичных B-клеток. (D — F) Количественная оценка RT-qPCR транскриптов до переключения (транскриптов Sx) в нестимулированных и активированных на 2 день В-клетках селезенки, клетках Ch22 или клонах 5, 6 и 8 в ответ на LPS (Sγ3) ( D ), к LPS + IFNγ (Sγ2a) ( E ) или к LPS + IL4 (Sγ1 и Sε) ( F ) (n = 3).
Количественная оценка транскриптов до переключения в нестимулированных (UNS) клетках показала, что уровни транскрипта s были выше в клонах с делецией, чем в Ch22, за исключением Sγ2a (Рис. 2D – F, Дополнительный Рис. S5). Более того, за исключением транскриптов Sε, уровни которых увеличиваются в активированных клонах с делецией Iα (фиг. 2F), переключающие транскрипты больше не индуцируются после стимуляции (фиг. 2D-F).
Мы пришли к выводу, что делеция коммитированного промотора Iα активирует большинство некоммитированных промоторов I.
Промотор Iγ2b индуцируется в ответ на LIT, но не на стимуляцию LPS в Ch22
В первичных B-клетках CSR к IgG2b индуцируется в ответ на LPS или LIT. Мы спросили, индуцируется ли CSR к IgG2b в клетках Ch22 либо стимулом, либо только в ответ на индуктор коммитированного изотипа ( i . e . LIT).
FACS-анализ показал, что, в отличие от первичных В-клеток, стимуляция ЛПС не индуцирует CSR для IgG2b ни в клонах с удаленным Ch22, ни в Iα (фиг. 3A).В то время как в ответ на LIT клетки Ch22 также не смогли переключиться на IgG2b, клетки с делецией Iα претерпели значительное переключение на IgG2b (фиг. 3B). Хотя они варьировались от клона к клону, уровни CSR для IgG2b в мутантных клонах всегда были ниже по сравнению с CSR для IgA в клетках Ch22 (фиг. 1B, C и 3B). Соответственно, уровни транскриптов Iµ-Cγ2b увеличивались только после стимуляции LIT (дополнительный рисунок S6). Более того, в клонах, стимулированных LIT, были обнаружены более высокие уровни транскриптов до переключения Sγ2b по сравнению с аналогами, стимулированными LPS (рис.3C – E). Удивительно, но в отличие от первичных B-клеток, где LPS индуцирует транскрипцию Sγ2b, LPS репрессировал транскрипцию Sγ2b в линии Ch22, а также во всех клонах с делецией Iα (рис. 3C).
Рисунок 3
CSR к IgG2b частично восстанавливается в ответ на TGF-β, но не на стимуляцию LPS. ( A , B ) Клетки Ch22, три клона с делецией Iα (2, 3 и 5) и В-клетки селезенки были активированы LPS ( A ) или LIT ( B ) в течение 4 дней, и окрашен на IgG2b. Представительные графики показаны для нестимулированных (UNS), активированных клеток Ch22, клонов с делецией Iα (клон 5) и первичных B-клеток.( C , D ) Количественная оценка RT-qPCR уровней транскриптов Sγ2b до переключения в нестимулированных клонах 5, 6 и 8 и в ответ на LPS ( C ) или на LIT ( D ) (день 2) . ( E ) Сравнение уровней транскриптов перед переключением Sγ2b в клонах 5, 6 и 8 с делецией Iα после стимуляции LPS и LIT (n = 3).
Эти данные показывают, что транскрипция Sγ2b и последующая CSR в IgG2b индуцируются в клонах с делецией Iα, но только в ответ на индуктор коммитированного изотипа (LIT).
Дифференциальная индукция переключающей транскрипции и CSR в присутствии TGF-β
Неожиданное открытие, что IgG2b отвечает только на стимуляцию LIT, позволило нам предположить, что промотор Iγ2b по-разному реагирует в линии Ch22 по сравнению с первичными В-клетками. Учитывая, что высокие уровни переключения на IgA могут быть достигнуты при активации клеток Ch22 анти-CD40 + IL4 + TGFβ (CIT) 21,22 , мы задались вопросом, повлияет ли и как этот стимул на CSR на IgG2b.
В Ch22 клетки переключились на более высокие уровни IgA при CIT, чем LIT (рис.1B и 4) и низкий процент клеток, переключившихся на IgG2b в присутствии CIT (рис. 3B и 4). Точно так же в клонах с делецией In Iα переключение на IgG2b было значительно выше с CIT, чем с LIT (фиг. 4). Учитывая влияние CIT на CSR для IgG2b, мы проанализировали CSR для других изотипов в клетках, обработанных CIT. Мы обнаружили, что переключение на IgG3 происходило с разной эффективностью, но уровни переключения были выше в ответ на CIT, чем на LIT (рис. 4). Переключение на IgG1 происходило только с CIT, в то время как CSR на IgG2a не определялся независимо от стимуляции (рис.4).
Рисунок 4
Дифференциальная индукция CSR в ответ на LIT и CIT. Клетки Ch22 и три клона с делецией Iα (3, 5 и 8) активировали LIT или CIT в течение 4 дней и окрашивали на указанные изотипы. Представительные графики показаны для активированных клеток Ch22 и клонов с делецией Iα (клон 5) (n ≥ 3).
Затем мы спросили, сопровождалось ли увеличение переключения с CIT по сравнению с LIT увеличением транскрипции переключения. В то время как уровни транскриптов перед переключением Sγ2b увеличивались при стимуляции CIT в клетках Ch22, не было различий в уровнях транскриптов между LIT и CIT в делеционных клонах, тогда как в B-клетках селезенки уровни транскриптов Sγ2b были выше при LIT (рис.5А). Неожиданно, хотя FACS обнаружил более высокий CSR к IgA с CIT, уровни транскриптов Sα в клетках Ch22 были выше с LIT, чем с CIT, но не было различий в активированных B-клетках селезенки (фиг. 5B). Клоны с делецией Iα, как и ожидалось, не продуцировали транскрипты Sα.
Рисунок 5
Дифференциальная индукция переключающей транскрипции в ответ на LIT и CIT. ( A — F ) Количественная оценка RT-qPCR Sγ2b ( A ), Sα ( B ), Sγ2a ( C ), Sγ3 ( D ), Sγ1 ( E ) и Уровни транскриптов Sε ( F ) до переключения в клетках Ch22 и Iα-делетированных клонах 3, 5 и 8 после стимуляции LIT или CIT.Уровни транскриптов в В-клетках селезенки, активированных LIT и CIT для Sγ2b ( A ) и Sα ( B ), и с LPS + IL4 (LI), LIT и CIT для Sγ1 и Sε ( E , F ) ) упакованы (n ≥ 3).
Хотя наблюдалось повышение CSR на IgG3 с CIT и отсутствие переключения на IgG2a, независимо от стимуляции, уровни транскриптов Sγ2a (за исключением клона 5) и Sγ3 были сопоставимы между LIT- и CIT-активированными делеционными клонами (рис. 5C, D).
В клетках Ch22 и в клонах с делецией LIT репрессировал транскрипцию Sε, но не Sγ1, за исключением клона 8, в котором количество транскриптов Sγ1 увеличивалось.CIT индуцировал транскрипцию Sγ1 и Sε (рис. 5E, F и дополнительный рис. S7). В B-клетках селезенки CIT был так же эффективен, как LPS + IL4 (LI) в индукции транскрипции Sγ1 и Sε, и уровни были выше, чем с LIT (рис. 5E, F).
Таким образом, в отсутствие коммитированного промотора Iα CSR на IgG2a не происходит, и хотя переключение на IgG1 происходит только с CIT, переключение на IgG2b и IgG3 происходит в ответ как на CIT, так и на LIT. Однако переключение было постоянно выше в ответ на CIT.Удивительно, но эффективность переключения не всегда коррелировала с уровнями транскриптов переключения.
Отсутствие корреляции между уровнями транскриптов
Aicda и эффективностью CSR
Поскольку фермент AID абсолютно необходим для инициирования CSR, мы задались вопросом, не связано ли отсутствие корреляции между переключаемой транскрипцией и CSR в некоторых случаях более низкой экспрессией Ген Aicda , кодирующий AID.
Мы обнаружили, что уровни транскриптов Aicda были выше в клетках Ch22 и производных клонах по сравнению с первичными В-клетками как в нестимулированных, так и в стимулированных условиях (рис.6). LI (LPS + IL4) не индуцировал транскрипцию Aicda в клонах с удаленными Ch22 и Iα, а только в первичных В-клетках (фиг. 6). В отличие от LI, LIT и CIT эффективно индуцировали ген Aicda во всех клетках (рис. 6).
Рисунок 6
Стимул-зависимая индукция транскрипции гена Aicda . Клетки Ch22, три клона с делецией Iα (3, 5 и 8) и В-клетки селезенки активировали LPS + IL4 (LI), LIT или CIT в течение 2 дней. Суммарные РНК собирали из нестимулированных и активированных клеток, и транскриптов Aicda количественно определяли с помощью RT-qPCR.Актин Транскрипты использовали для нормализации (n ≥ 3).
Следовательно, хотя отсутствие переключения на Sε в Ch22 и делеционных клонах после стимуляции LI коррелирует с отсутствием индукции гена Aicda , то же самое неверно для различий в CSR для IgG2b и IgA в присутствии LIT и CIT.
Повышенная транскрипция энхансера hs1-2 в ответ на LIT, но не на CIT
Энхансерные РНК (эРНК) продуцируются на 3’RR после активации В-клеток селезенки и являются отличительным признаком активности 3’RR 25,26, 27 .Затем мы задались вопросом, можно ли объяснить отсутствие корреляции между переключаемой транскрипцией и CSR в случаях, когда имеется сходная активация Aicda , различиями в транскрипционной активности 3’RR. Чтобы проверить это, мы количественно определили уровни транскриптов hs3a, hs1-2 и hs3b.
Уровни
эРНК были сопоставимы между нестимулированными клетками Ch22 и делеционными клонами и были выше, чем в В-клетках селезенки (фиг. 7). Как для клеток Ch22, так и для делеционных клонов уровни эРНК hs3a и hs3b существенно не различались в зависимости от LIT и CIT (несмотря на некоторые клональные вариации) (рис.7), наблюдалась устойчивая тенденция к увеличению транскрипции hs1-2 с LIT (рис. 7).
Фигура 7 Уровни транскриптов
3’RR после стимуляции LIT и CIT. Количественная оценка уровней эРНК энхансеров hs3a, hs1-2 и hs3b с помощью RT-qPCR. Тотальные РНК экстрагировали из нестимулированных клеток Ch22 и клонов с делецией Iα (3, 5 и 8) на 2 день после стимуляции. Актин Транскрипты были использованы для нормализации, и (-RT) контроль был включен повсюду (n ≥ 3).
В то время как усиленная транскрипция hs1-2 в ответ на LIT коррелирует с низкими уровнями транскриптов Sγ1 и Sε, высокие уровни транскрипции Sγ2b и Sα были обнаружены в присутствии LIT.Следовательно, усиление транскрипции энхансера hs1-2 не может само по себе объяснить диссоциацию между переключением транскрипции и CSR.
Садбхавана — Ячейка CSR
×
Приветствие от Института менеджмента и массовых коммуникаций Сурядатта (SIMMC)
Согласно директивам, полученным от компетентных органов, наш кампус закрыт в связи с распространением COVID 19 (CORONA VIRUS).
Мы уверены, что ваша семья и вы в безопасности, и принимаем необходимые меры предосторожности против печального распространения COVID 19.
Мы всегда доступны для всех заинтересованных сторон и хотели бы помочь вам всеми возможными способами.
В связи с COVID 19, в настоящее время GD / PI для MBA осуществляется посредством телефонных звонков, видеоконференцсвязи, видеозвонков Whatsapp или других виртуальных режимов. Заинтересованные кандидаты должны отправить sms свое имя, курс, штат и город, интересующий курс на 8956932418/9763266829
Для профориентации отправьте SMS с вашим именем, городом, номером мобильного телефона и курсом, для которого требуется консультация, на номер 9763266829.
Для справок и приема обращайтесь:
г-жа Пуджа 8956932418, г-жа Свапниша 8956932412, г-жа Мили 9112297607,
г-жа Абхишри 8956943821, г-жа Анага 8956932402, г-жа Радхика 8669602051
По вопросам учебы и экзаменов обращаться:
Доктор Пратикша Вейбл (профессор и декан)
7588943101, др[email protected]
Для отдела консультирования, обучения и трудоустройства MDP
Проф. Сунил Дхадивал (декан по корпоративным отношениям — обучение и трудоустройство)
8956932417/9763266829, [email protected]
По вопросам маркетинга, СМИ и координации с выпускниками
Г-н Акшит Кушал (Исполнительный директор и секретарь — Ассоциация выпускников)
8956943820/9762478166/9763266829, акшит[email protected]
Для администрирования и покупки
Г-н Рохит Санчети (Центральный управляющий)
8956932401/9763266829 [email protected]
Также можете позвонить:
Д-р Шайлеш Касанде (генеральный директор и директор группы)
99211 11099 [email protected]
Оставайтесь в доме Оставайтесь в безопасности… !!!
Центральная серозная хориоретинопатия (CSR) — Центр диагностики сетчатки
Что такое центральная серозная ретинопатия?
Центральная серозная ретинопатия (CSR) обычно поражает молодых мужчин и женщин в возрасте 30, 40 и 50 лет.Симптомы находятся в центре поля зрения — размытость, искажение, минификация объектов и темное пятно при фиксации. Он представляет собой жидкость, спонтанно просачивающуюся из сосудистой оболочки сетчатки в пространство сетчатки. Сетчатка располагается поверх большого слоя ткани, заполненной кровеносными сосудами и капиллярами, называемыми сосудистой оболочкой. Хориоидея функционально отделена от сетчатки мембраной (мембрана Бруха) и одним слоем клеток, называемым пигментным эпителием сетчатки (ППЭ). Этот слой работает как барьер, облегчающий поступление питательных веществ в сетчатку из нижележащих сосудов.Это также позволяет отходам сетчатки удаляться хориоидальными сосудами. Слой RPE препятствует свободному течению жидкости. При CSR это уплотнение разрушается, и жидкость просачивается в субретинальное пространство. Это может происходить в обоих глазах, но обычно проявляется по одному. Через пару месяцев состояние может исчезнуть само по себе. Если у кого-то был начальный эпизод, вероятность повторного эпизода на любом глазу составляет до 50%. Повторяющиеся эпизоды в одном и том же месте или постоянная утечка в одном месте могут нанести непоправимый вред зрению.Редкое, но серьезное осложнение возникает, когда рубцовая ткань и кровеносные сосуды от сосудистой оболочки проникают в пространство под сетчаткой. Эта ткань, если ее не остановить, может продолжать расти и разрушать центральную сетчатку (или макулу). Это чаще встречается у пожилых пациентов с рецидивами.
Каковы причины?
Существует много предположений о том, что вызывает центральную серозную ретинопатию. Догма предполагает, что стресс может играть большую роль в его развитии. В нашей клинике мы обнаружили, что пациенты не всегда испытывают стресс, а корпоративная социальная ответственность может быть больше связана с усталостью или истощением.Это чаще встречается у пациентов, у которых слишком много вещей на тарелке, и обычно связано с недостатком сна. Большинство пациентов сообщают о 5-6 часах сна ночью, а не о рекомендуемых 8 часах. Продолжительное и постоянное недосыпание может сократить продолжительность вашей жизни. Также было связано воздействие любых стероидных препаратов (назальный спрей, крем с кортизоном, инъекции в суставы и т. Д.).
Какие методы лечения?
Лечение центральной серозной ретинопатии в настоящее время осуществляется одним из трех способов.
Наблюдение: Наблюдение часто используется в течение первых месяцев или двух, чтобы увидеть, может ли центральная серозная ретинопатия пройти сама по себе. Пациенту следует попытаться спать по 8 часов в сутки, установив обычное время сна. Регулярные упражнения, кажется, помогают добиться более регулярного цикла сна.
Лазерное лечение: Лазерное лечение центральной серозной ретинопатии было одним из основных направлений активной терапии. Идея состоит в том, что при небольшом количестве термической лазерной стимуляции клетки РПЭ могут образоваться рубцы, и утечка жидкости прекратится.Это лечение можно использовать в любое время после постановки диагноза центральной серозной ретинопатии. Он показан ранее при зрении только на один глаз, и этот глаз поражен центральной серозной ретинопатией. Первым кандидатом на немедленное лечение может быть человек, который не может выполнять свои обычные задачи из-за нарушения зрения, например, пилот авиакомпании.
Фотодинамическая терапия (Visudyne): Это лечение кажется достаточно эффективным в некоторых случаях центральной серозной ретинопатии и не оставляет таких рубцов, как при использовании лазера.Лечение Visudyne, по-видимому, потенциально эффективно не только против центральной серозной оболочки, но также и против редкого осложнения в виде неоваскулярной мембраны под сетчаткой или роста рубцовой ткани, лежащей в основе утечки в глазу. Визудин вводится в вену и предпочтительно связывается с аномальным кровеносным сосудом. Затем используется «холодный лазер» для активации препарата и закрытия аномально протекающих сосудов.
Copyright 2007, Центр диагностики сетчатки глаза.
Факторы ответа на двухцепочечный разрыв ДНК влияют на характеристики концевых соединений переключателя класса IgH и общие транслокационные соединения переключающая рекомбинация (CSR).Во время CSR разрывы ДНК вводятся в две специфические области локуса антитела, и эти два разрыва ДНК соединяются, образуя новую форму гена антитела. Как и другие разрывы генома, разрывы ДНК CSR активируют путь ответа на повреждение клеточной ДНК, который помогает обеспечить их надлежащее восстановление. Теперь мы показываем, что, когда определенные компоненты пути ответа на повреждение ДНК инактивированы, соединения CSR В-клеток показывают разные молекулярные сигнатуры, что указывает на то, что они репарации репарации менее эффективны альтернативным путем репарации ДНК вместо обычного общего репарации разрыва клеточной ДНК. путь.
Abstract
Переключающая рекомбинация класса тяжелой цепи (IgH) (CSR) в B-лимфоцитах переключает константные области IgH для изменения функций антител. CSR инициируется двухцепочечными разрывами ДНК (DSB) внутри донорской области переключения (S) IgH и расположенной ниже акцепторной области S. CSR завершается слиянием концов DSB донорной и акцепторной S-области за счет классического негомологичного соединения концов (C-NHEJ) и, в его отсутствие, за счет альтернативного соединения концов, которое более склонно к использованию более длинных соединительных микрогомологий (MHs).Факторы дефицита DSB-ответа (DSBR), включая мутированную атаксию и телеангиэктазию (ATM) и 53BP1, по-разному нарушают соединение концов CSR, причем наибольшее влияние оказывает дефицит 53BP1. Однако исследования потенциального влияния дефицита фактора DSBR на MH-опосредованное соединение концов CSR были технически ограничены. Теперь мы используем надежный анализ соединения DSB, чтобы выяснить влияние дефицита факторов DSBR на CSR и соединения хромосомной транслокации в первичных B-клетках мыши и клетках B-лимфомы Ch22F3.По сравнению с диким типом, соединения транслокации CSR и c-myc в S-область в отсутствие 53BP1 и, в меньшей степени, других факторов DSBR увеличивают утилизацию MH; действительно, профили MH с дефицитом 53BP1 напоминают профили, связанные с дефицитом C-NHEJ. Однако транслокационные соединения между c-myc DSB и общими DSB в масштабе всего генома не смещены по MH в ATM-дефицитных клетках по сравнению с Ch22F3 дикого типа и менее смещены в 53BP1- и C-NHEJ-дефицитных клетках, чем CSR-соединения или Переходы транслокации c-myc в S-область.Мы обсуждаем потенциальную роль факторов DSBR в подавлении повышенного MH-опосредованного присоединения концов DSB и особенности S-регионов, которые могут сделать их DSB склонными к MH-смещенному концевому соединению в отсутствие факторов DSBR.
Зрелые В-клетки подвергаются переключающей рекомбинации (CSR) класса тяжелой цепи Ig (IgH) для изменения константной области цепей IgH и, таким образом, модуляции эффекторных функций антитела. У мышей шесть наборов экзонов C H лежат на 100-200 т.п.н. ниже первоначально экспрессированного экзона C H (Cμ) и экспрессируются в контексте CSR; эти наборы нижележащих экзонов C H включают Cγ3, Cγ1, Cγ2b, Cγ2a, Cε и Cα (1).Каждый набор экзонов C H фланкирован выше длинной (от 1 до 10 т.п.н.) повторяющейся областью переключателя ДНК (S), которая содержит большое количество коротких мотивов ДНК, которые являются мишенями для индуцированной активацией цитидиндезаминазы (AID) ( 2). AID инициирует CSR, вводя повреждения дезаминирования цитидина в ДНК донора Sμ и в расположенные ниже акцепторные области S, которые затем преобразуются в двухцепочечные разрывы ДНК (DSB) за счет координированной активности путей удаления оснований и / или репарации ошибочного спаривания (2). Продуктивная CSR завершается делеционным соединением вышестоящего конца донорного Sμ DSB с нижним концом акцепторной S-области DSB в реакции CSR, которая механически принудительно вызывает продуктивные делеционные события присоединения CSR с гораздо большей частотой (10 — в 20 раз), чем непродуктивные инверсионные соединения (3).
Нормальное соединение концов CSR DSB осуществляется преимущественно классическим негомологичным соединением концов (C-NHEJ) (1, 4). В соответствии с C-NHEJ, многие соединения CSR в нормальных B-клетках происходят посредством прямого («тупого») соединения концов DSB (до 30% в зависимости от S-области), а остальная часть связана с соединением концов DSB через короткие (1 п.н. до 4 п.н.) перекрытия микрогомологий (MH) (4). В-клетки, дефицитные по основным компонентам C-NHEJ (например, XRCC-4, лигаза 4, Ku70 или Ku80), имеют умеренно сниженную (от 40 до 50% от нормы) CSR и повышенные AID-зависимые хромосомные разрывы IgH; CSR-соединения в таких ядерных C-NHEJ-дефицитных B-клетках образуются посредством альтернативных концевых соединений (A-EJ) и в основном (90% или более) опосредованы MH (4⇓⇓ – 7).CSR DSB запускают атаксию, зависимую от телеангиэктазии (ATM) -зависимый DSB-ответ (DSBR), при котором киназа ATM фосфорилирует нижележащие субстраты, включая гистон h3AX и 53BP1, которые накапливаются в фокусах, окружающих DSB (8). Дефицит ATM или h3AX умеренно нарушает CSR (от 30 до 50% от нормы) с соответствующим повышенным накоплением разрывов IgH , что указывает на роль этих белков в соединении концов CSR (1, 8). Напротив, дефицит 53BP1 более серьезно нарушает CSR (5% от нормы), что также сопровождается увеличением разрывов IgH (1, 8).ATM DSBR, вероятно, способствует соединению концов CSR путем связывания изолированных DSB S-области для повторного соединения, тем самым поддерживая интактный хромосомный субстрат для продуктивного соединения CSR (1, 3, 9). Факторы DSBR также действуют для предотвращения резекций DSB-концов S-области, которые могут генерировать длинные выступающие концы, не подходящие для C-NHEJ, но которые могут обнажать MH-богатые субстраты в форме коротких повторяющихся последовательностей S-области для A-EJ (3, 10, 11). В связи с этим большее влияние дефицита 53BP1 на соединение CSR может отражать как нарушение привязки концов, так и существенную активацию увеличения резекции DSB из-за неспособности активировать связанный фактор блокирования резекции Rif1 (3).
Вопрос о том, способствует ли дефицит фактора DSBR увеличению использования MH в CSR, окончательно не решен. Ряд исследований пришел к выводу, что дефицит ATM (12), h3AX (13), 53BP1 (11, 14) или Rif1 (15) не приводит к изменению использования MH в соединениях CSR по сравнению с таковым у B-клеток WT. С другой стороны, в других исследованиях сообщалось, что дефицит ATM действительно приводит к увеличению использования MH в соединениях CSR мыши и человека (16, 17). Основным ограничением всех предыдущих исследований соединения концов CSR в В-клетках с дефицитом фактора DSBR является то, что благодаря технологии, доступной в то время для анализа, длинная повторяющаяся природа S-областей позволяла оценивать только относительно короткие последовательности на периферийных краях S-области, расположенные между двумя праймерами ПЦР.В результате такие исследования соединений CSR на основе ПЦР дали небольшое количество соединений, которые не происходят из повторяющихся ядер S-области, где возникает большинство нормальных соединений CSR. Кроме того, выводы многих предыдущих исследований были основаны на относительно небольших размерах выборки, опять же на основе ограничений технологий, доступных в то время.
Мы разработали и усовершенствовали высокопроизводительное геномное секвенирование транслокаций (HTGTS) для картирования с разрешением нуклеотидов соединений между HTGTS «приманкой» DSB и эндогенными DSB в масштабе всего генома (18–20).DSB приманки HTGTS могут быть введены эктопически с помощью дизайнерских нуклеаз или могут быть обеспечены повторяющимися эндогенными DSB, включая те, которые инициируют IgH CSR или рекомбинацию V (D) J (3, 20). В последнем контексте мы адаптировали HTGTS для использования в качестве надежного эндогенного анализа CSR, используя серию AID-инициированных DSB приманки в области 5’Sμ для анализа AID-индуцированных DSB жертвы в нижележащих акцепторных S-областях в CSR-активированных. нормальные первичные В-клетки или клетки В-лимфомы Ch22F3 (3, 21). Этот анализ HTGTS CSR предлагает много преимуществ по сравнению с предыдущими анализами соединения концов CSR, поскольку он позволяет восстановить на несколько порядков больше соединений CSR и идентифицирует соединения CSR по всей длине повторяющихся S-областей акцептора (3).Чтобы дополнительно выяснить потенциальную роль факторов DSBR в MH-опосредованном соединении концов во время CSR и хромосомных транслокаций, мы проанализировали паттерны соединений большого количества соединений DSB, которые произошли либо во время CSR, либо во время транслокаций между Cas9 / ведущей РНК (gRNA)-индуцированной. c-myc приманка DSB к DSB S-области или другим общим DSB по всему геному в WT и дефицитных по фактору DSBR первичных B-клетках и клетках Ch22F3, активированных для CSR.
Результаты
HTGTS Анализ профилей соединений CSR.
Мы изучили паттерн соединения инициированных AID разрывов S-области во время CSR в B-клетках селезенки мышей, активированных в культуре с αCD40 / IL-4 для стимуляции CSR между Sμ и Sγ1 или Sε. Сначала мы проанализировали десятки тысяч соединений CSR, восстановленных в наших предыдущих исследованиях зависимости ориентации CSR на основе HTGTS, в которых сравнивали WT и В-клетки с дефицитом DSBR (3). Эти тесты HTGTS CSR были основаны на использовании праймера 5’Sμ, который позволил серии инициируемых AID DSB в самой 5′-области Sμ служить в качестве комбинированной приманки HTGTS для захвата соединений с AID-индуцированными DSB жертвы в нисходящем направлении. core области Sγ1 или Sε (рис.1 А ). Узлы CSR, проанализированные в настоящем исследовании, можно разделить на два общих класса (4). Первый — это «прямые» соединения, состоящие из последовательностей приманки, за которыми непосредственно следуют последовательности жертвы без перекрывающихся нуклеотидов. Второй — это «MH-опосредованные» соединения, которые состоят из соединений с перекрывающимися нуклеотидами ≥1 п.н., которые, скорее всего, обеспечиваются внутри выступающих лоскутов резецированных концов приманки и жертвы (Fig. 1 A ). Мы также восстановили значительное количество (около 20%) соединений с ≥1 п.н. встроенной последовательности, потенциально возникающих в результате активности PolQ (22) или неизвестных источников, из которых нельзя отличить прямые механизмы соединения от опосредованных MH ( SI Приложение ). , Таблица S1).Мы проанализировали тысячи переходов и выразили процент прямых переходов и количество переходов с различной длиной MH для данной S-области как процент от общего числа переходов в эту S-область (рис.1 B и E ). Результаты этих экспериментов были хорошо воспроизводимы (Рис. 1 B и E и SI Приложение , Рис. S1 и Таблица S2).
Рис. 1.
Повышенное использование MH в Sμ-Sγ1 и Sμ-Sε CSR-соединениях из первичных B-клеток с дефицитом фактора DSBR.( A ) Иллюстрация результатов присоединения DSB области Sμ к нижележащим разрывам области Sγ1 и Sε, претерпевающим прямое или опосредованное MH соединение. WT, ATM — / , — и 53BP1 — / — первичные В-клетки селезенки стимулировали αCD40 / IL4 в течение 96 часов. ( B и E ) Использование MH от переходов с прямым и до 10 пар оснований MH для 5’Sμ ( B ) к Sγ1 и ( E ) к Sε DSB показано как процент от общего числа переходов.( C , D , F и G ) Процент прямых соединений от 5′Sμ до ( C ) Sγ1 и ( F ) Sε, а также процент соединений с 4-bp или более Сравниваются MH в ( D ) Sγ1 или ( G ) Sε в различных генетических фонах. Непарный двусторонний тест t был использован для расчета значений P для значимой разницы между образцами (количественные данные = среднее ± SEM; ** P ≤ 0,01, *** P ≤ 0.001). Для каждого эксперимента использовали не менее трех независимых выборок. SI Приложение , таблица S2 содержит подробную информацию, включая номера соединений и процент прямых и опосредованных MH соединений, найденных для каждой из согласованных библиотек без размера, проанализированных для генерации данных, описанных на этом рисунке. Мы также получили очень похожие результаты при анализе трех репликативных библиотек для каждого генотипа, в которых порядковые номера соединений Sμ-Sγ1 или Sμ-Se из каждого экспериментального образца были нормализованы путем случайного отбора к таковому из экспериментального образца с наименьшим количеством восстановленных соединений (см. SI Приложение , рис.S1 для подробностей).
Прямые и опосредованные MH соединения CSR в WT по сравнению с В-клетками селезенки с дефицитом DSBR.
Суммарные (комбинированные делеционные и инверсионные) Sμ – Sγ1 и Sμ – Sε CSR-соединения в WT анти-CD40 плюс IL-4-обработанных В-клетках были примерно на 30% прямыми, при этом большинство других использовали от 1 до 3 пар оснований MH и небольшая фракция, использующая более длинные MH (рис. 1 B и E ). Напротив, ATM-дефицитные B-клетки имели заметное увеличение MH-опосредованных соединений Sμ – Sγ1 и Sμ – Sε, причем около 20% каждого из них были прямыми (рис.1 C и F ) и соответственно увеличенная доля MH-опосредованных соединений (рис. 1 B и E ). Примечательно, что у 53BP1-дефицитных В-клеток с серьезным нарушением CSR наблюдается наибольшее уменьшение прямых соединений, при этом менее 10% соединений Sμ – Sγ1 и Sμ – Sε CSR являются прямыми (рис. 1 C и F ) и значительно увеличенная фракция MH-опосредованных соединений (рис. 1 B и E ). h3AX-дефицитные и Rif1-дефицитные B-клетки демонстрируют ~ 20% прямых соединений и общий паттерн CSR-соединений, очень похожий на таковой у ATM-дефицитных B-клеток и отличный от такового для 53BP1-дефицитных B-клеток ( SI Приложение , рис.S2). В этом отношении, клетки WT имели среднюю длину MH 1,67 п.н. для Sμ-Sγ1 и 1,48 п.н. для Sμ-Sε CSR-переходов, в то время как, с другой стороны, В-клетки с дефицитом 53BP1 имели среднюю длину MH 3,17 п.н. и 2,8 п.н. для этих двух наборов соединений ( SI Приложение , Таблица S3). ATM, h3AX и Rif1-дефицитные B-клетки имели CSR-соединения с увеличенной средней длиной MH, которые были опосредованы между WT и 53BP1-дефицитными B-клетками ( SI Приложение , Таблица S3). Наконец, мы также оценили относительное использование MH в делеционных и инверсионных соединениях Sμ – Sγ1 и Sμ – Sε в генотипах WT по сравнению с генотипами с дефицитом DSBR и наблюдали те же общие закономерности, указывающие на то, что использование прямого и опосредованного MH соединения концов не оказывает заметного вносят вклад в инверсионную рекомбинацию CSR по сравнению с делеционной ( SI, приложение , таблица S3).
Для дальнейшего выяснения влияния дефицита различных факторов ответа DSB на концевое соединение, используемое для формирования соединений CSR, мы оценили относительный процент соединений CSR Sμ – Sγ1 и / или Sμ – Sε для каждого генотипа, который использовал MH 4 п.н. или больше, длина считается неблагоприятной для присоединения через C-NHEJ (23). В соответствии с преобладающим присоединением соединений CSR через C-NHEJ в клетках WT, только около 12% и 8% соединений Sμ – Sγ1 и Sμ – Sε соответственно достигли длины 4 п.н. или более (рис.1 D и G ). Напротив, относительные уровни этих более длинных MH-опосредованных соединений из ATM-дефицитных B-клеток были увеличены примерно до 24% и 20% соответственно. для соединений Sμ – Sγ1 и Sμ – Sε (рис. 1 D и G ), и h3AX- и Rif1-дефицитные В-клетки показали аналогичное увеличение ( SI Приложение , рис. S2 C , F , и I ). Примечательно, однако, что уровни этих длинных MH-опосредованных соединений были самыми высокими для соединений CSR в 53BP1-дефицитных В-клетках, достигая 35% и 33%, соответственно, для соединений Sμ – Sγ1 и Sμ – Sε, примерно от 3 до 4 уровней. раз больше, чем наблюдаемые для В-клеток дикого типа (рис.1 D и G ).
CSR-соединения в WT по сравнению с клетками Ch22F3, дефицитными по DSBR или лигазе 4.
Мы исследовали относительные уровни прямых и MH-опосредованных соединений CSR в Sα посредством исследований клеток B-лимфомы Ch22F3 мыши, которые можно стимулировать, чтобы они подвергались устойчивому CSR к IgA при стимуляции αCD40 / IL4 / TGF-β в культуре (24) . С этой целью мы генерировали ATM-дефицитные и 53BP1-дефицитные клетки Ch22F3 путем делеции гена, опосредованной CRISPR-Cas9 ( SI Приложение , рис.S3 A — D ). Чтобы избежать потенциально мешающих эффектов ранее переключенного непродуктивного аллеля IgH с переключением IgA на оценку соединений CSR, мы также удалили предварительно переключенную область Sμ-Sα непродуктивного аллеля IgH во всех генотипах (рис.2 A и SI, приложение , рис. S3 E ). Чтобы учесть клональные вариации в клетках Ch22F3, мы проанализировали две независимо полученные клеточные линии для каждого генотипа. После 72 часов стимуляции αCD40 / IL4 / TGF-β клетки Ch22F3 с дефицитом ATM и 53BP1 показали уровни CSR, которые, соответственно, составляли в среднем ∼35% и 25% от уровней переключения класса IgA дикого типа ( SI Приложение , рис.S3 F ). В соответствии с известными клональными вариациями CSR в клеточных линиях Ch22F3 (25), мы наблюдали значительные различия в общих уровнях переключения классов IgH между двумя независимыми клонами каждого генотипа ( SI, приложение , рис. S3 F ).
Рис. 2.
Повышенное использование MH в Sμ-Sα CSR-соединениях из WT, ATM-, 53BP1- и клеток Ch22F3, дефицитных по лигазе 4. ( A ) Иллюстрация конструирования непродуктивных аллель-удаленных клеток Ch22F3 и стратегии HTGTS для анализа соединений Sμ-Sα с приманкой 5′Sμ после 72-часовой стимуляции αCD40 / IL4 / TGF-β.( B ) Использование MH в соединениях от присоединения AID-индуцированных разрывов 5’Sμ к разрывам Sα в непродуктивных аллель-удаленных WT, ATM — / — , 53BP1 — / — и лигазе 4 — / — Клетки Ch22F3 наносили на график как процент соединений с указанной длиной MH от общего числа соединений в области Sα. ( C и D ) Процент прямых соединений ( C ) и ( D ) соединений с 4-пн и более длинными MH в соединении Sμ − Sα в непродуктивном WT с удаленным аллелем, ATM — / — , 53BP1 — / — и лигаза 4 — / — Ch22F3 клеток.Двусторонний тест t использовался для статистического анализа (количественные данные = среднее ± SEM; *** P ≤ 0,001). Для каждого эксперимента использовались три независимых образца, подробные сведения приведены в приложении SI , таблица S4.
Мы использовали анализ HTGTS CSR для восстановления 5’Sμ-Sα-переходов из двух наборов стимулированных WT и мутантных клеток Ch22F3 и проанализировали использование MH в восстановленных соединениях из трех независимых экспериментов для каждой линии; Эти эксперименты дали согласованные результаты между независимыми клонами каждого генотипа (рис.2 B — D и SI Приложение , таблица S4). Суммарные Sμ-Sα CSR-соединения в клетках WT Ch22F3 были на ~ 25% прямыми, с большинством других соединений MH от 1 до 4 пар оснований (фиг. 2 B ). Напротив, в клетках Ch22F3 с дефицитом ATM наблюдалось заметное увеличение MH-опосредованных соединений Sμ-Sα, причем только около 15% прямых соединений (Fig. 2 C ). Клетки Ch22F3 с дефицитом 53BP1, как наблюдали для первичных B-клеток с дефицитом 53BP1, показали наибольшее уменьшение прямых соединений Sμ-Sα, при этом менее чем на 10% были прямые (рис.2 С ). ATM-дефицитные и 53BP1-дефицитные клетки Ch22F3 также имели соответствующее увеличение средней длины MH переходов 5’Sμ-Sα ( SI Приложение , Таблица S5) и существенно увеличивали частоту переходов 5′Sμ-Sα, в которых использовались MH-соединения. 4 п.н. и более (рис. 2 D ).
Чтобы напрямую сравнить использование MH между В-клетками с дефицитом DSBR и C-NHEJ-дефицитом, мы также восстановили Sμ-Sα-соединения из клеток Ch22F3, дефицитных по основному фактору c-NHEJ лигазы 4, из которых мы также удалили предварительно переключенный Sμ -Sα область непродуктивного аллеля IgH ( SI Приложение , рис.S3 E ). По сравнению с WT «подобранными» (т. Е. Предварительно переключенными Sμ-Sα-удаленными) клетками Ch22F3, клетки Ch22F3 с дефицитом лигазы 4 показали значительно повышенное использование MH, повышенное среднее использование MH и повышенную частоту соединений Sμ − Sα с более чем 4 п.н. MH (Рис.2 B — D и SI Приложение , Таблица S5). Для каждого измеренного параметра значения были очень похожи на значения соответствующих 53BP1-дефицитных клеток Ch22F3 (рис. 2 B — D и SI, приложение , таблица S5).Наконец, по сравнению с WT или ATM-дефицитными клетками, клетки Ch22F3, дефицитные по 53BP1 и лигазе 4, также имели значительную долю направленных соединений, отображаемых в неповторяющихся 5′- и 3′-областях, прилегающих к Sα ( SI Приложение , рис. S4 A ), которые, как мы ранее пришли к заключению, происходят в контексте длинных резекций Sα DSB (3). Как показано ранее (3), соединения CSR клеток WT, ATM- и 53BP1 Ch22F3 соответственно показали возрастающий вклад длинных резекций CSR и соответствующую потерю зависимого от ориентации смещения соединения ( SI, приложение , рис.S4 B и C и таблицы S6 и S7). Примечательно, что, несмотря на сходство с 53BP1-дефицитными клетками в MH-опосредованных CSR-соединениях, дефицитные по лигазе 4 клетки Ch22F3 показали только промежуточно увеличенные длинные резекции и более умеренное влияние на ориентационно-зависимое соединение ( SI Приложение , рис. S4 B и C и таблицы S6 и S7).
Влияние дефицита DSBR на переходы транслокаций к общегеномным DSB по сравнению с CSR.
Чтобы определить, распространяется ли повышенное использование MH в соединениях CSR, наблюдаемое в клетках Ch22F3 с дефицитом DSBR-факторов, на другие DSB в масштабе всего генома, мы ввели опосредованный CRISPR-Cas9 DSB в локусе c-myc на хромосоме 15 и использовали HTGTS для анализ транслокационных соединений с Sμ и Sα DSB и другими DSB по всему геному.Мы провели эти исследования на αCD40 / IL4 / TGF-β-стимулированных клетках WT Ch22F3 и двух независимо полученных клонах, каждый из ATM-, 53BP1- и ligase 4-дефицитных клеток Ch22F3, все с непродуктивным аллелем Sμ-Sα интактным (рис. 3 А ). Соединения между DSB приманки c-myc и AID-инициированными Sμ и Sα DSB были удивительно похожи для каждого исследуемого генотипа на соединения Sμ-Sα CSR с точки зрения общих профилей соединений, частоты прямых соединений, среднего использования MH. , а также частота контактов с MH более 4 п.н. (рис.3 B — D и SI Приложение , рис. S5 и таблицы S8 и S9). По всем характеристикам объединения в этом наборе анализов транслокационные соединения клеток Ch22F3, дефицитных по 53BP1 и лигазе 4, были очень похожи друг на друга (рис. 3 B — D и SI, приложение , рис. S5). . Подобно CSR-соединениям в клетках, дефицитных по 53BP1 и лигазе 4, мы обнаружили, что значительная часть транслокаций между c-myc привлекает DSB в локали Sμ и, в частности, Sα, по сравнению с таковыми в других фонах, вовлекает DSB в неповторяющиеся области резекции за пределами S-областей ( SI Приложение , рис.S6).
Рис. 3.
Сравнение транслокационных соединений с S-областью или общими DSB в WT и мутантных клетках Ch22F3. Клетки Ch22F3 с дефицитом WT, ATM-, 53BP1 и лигазы 4 стимулировали αCD40 / IL4 / TGF-β, и через 12 ч после стимуляции вводили DSB приманки cmyc -Cas9. Библиотеки HTGTS были сделаны с использованием c-myc -Cas9 DSB в качестве приманки, с получением геномной ДНК через 72 часа после стимуляции. ( A ) Показаны результаты транслокации c-myc -Cas9 DSB в полногеномные разрывы или разрывы Sμ / Sα.( B и E ) Использование MH в соединениях от c-myc -Cas9 разрыва, соединяющегося с ( B ) разрывами Sα и ( E ) разрывами по всему геному, были нанесены на график как процент соединений с указанной длиной. MH по общему количеству перекрестков в соответствующих регионах. ( C , D , F и G ) Процент прямых соединений из c-myc -Cas9 разрывает соединение с ( C ) Sα разрывами и ( F ) разрывами на уровне всего генома показаны проценты соединений с 4 п.н. и более MH от разрывов c-myc -Cas9 до разрывов ( D ) Sα и разрывов по всему геному ( G ) в различных генетических фонах.Непарный двусторонний тест t использовали для статистического анализа (количественные данные = среднее ± SEM; н.у. означает P > 0,05; *** P ≤ 0,001). Для каждого эксперимента использовались три независимых образца, подробности представлены в Приложении SI , Таблица S9.
Для анализа профилей соединений транслокаций DSB c-myc bait с общегеномными DSB в том же наборе Ch22F3 WT и мутантных клеток мы исключили картирование обоих соединений в локусе IgH, чтобы избежать DSB S-области, а также те, которые находятся в области 20 mb, окружающей сайт разрыва c-myc , чтобы избежать соединений сайтов разрыва, опосредованных в основном резекцией (18, 19).Мы также исключили последовательность в известных точках соединения AID. Эти исследования показали, что транслокационные соединения между DSB c-myc bait и общими DSB по всему геному в WT и ATM-дефицитных клетках Ch22F3 были неотличимы в отношении частоты прямых соединений (рис. 3 E и F ). Напротив, клетки Ch22F3 с дефицитом 53BP1 и лигазы 4 имели значительно сниженные прямые транслокационные соединения по всему геному по сравнению с таковыми у WT и ATM-дефицитных клеток (фиг. 3 E и F ).Несмотря на аналогичные уровни прямых соединений с клетками WT, ATM-дефицитные клетки имели немного повышенную частоту соединений c-myc с общими соединениями транслокации DSB с MH более 4 п.н., и этот класс соединений был дополнительно увеличен в 53BP1- и лигазе. 4-дефицитные клетки (рис. 3 G ). Примечательно, что относительный уровень прямого соединения c-myc с общими соединениями транслокации DSB по сравнению с соединениями транслокации c-myc с Sα, по сравнению с соответствующими контролями дикого типа, был увеличен, а частота соединений с MH более 4 п.н. фоны с дефицитом ATM, 53BP1 и лигазы 4 (ср. рис.3 C и D с рис.3 F и G ). Наконец, по всем характеристикам объединения в этом наборе анализов транслокационные соединения клеток Ch22F3, дефицитных по 53BP1 и лигазе 4, снова были очень похожи (рис. 3 E — G ).
Discussion
Предыдущие исследования показали, что дефицит фактора DSBR практически не влияет на использование MH в соединениях CSR (12⇓⇓ – 15). Однако теперь мы демонстрируем, что дефицит различных ATM-зависимых факторов DSBR в активированных первичных B-клетках мыши и / или линии Ch22F3 мыши по сравнению с аналогами WT приводит к значительному общему увеличению MH-опосредованных по сравнению с прямыми CSR-соединениями между Sμ и все протестированные нижестоящие S-области (Sγ1, Sε и Sα).В этом контексте дефицит 53BP1 имел наиболее резкое увеличение, в то время как дефицит ATM, h3AX и Rif1 имел промежуточное увеличение. Относительное влияние этих различных DSBR-дефицитных фонов на уровень использования MH в соединениях CSR также хорошо коррелировало с восстановлением соединений с длинными MH (более 4 п.н.), что свидетельствует об увеличении использования A-EJ (4). Несколько факторов могут способствовать отличию наших текущих результатов от результатов предыдущих исследований. Во-первых, в большинстве предыдущих исследований из-за технических ограничений было проанализировано небольшое количество соединений CSR, тогда как наши текущие исследования соединений CSR на основе HTGTS восстановили на два-три порядка больше (3).Второй фактор заключается в том, что во многих предыдущих исследованиях анализировались только соединения, расположенные на периферийных краях S-регионов, из-за ограничений на праймеры, которые можно было бы использовать для используемых анализов на основе ПЦР (26). Напротив, наш анализ CSR на основе HTGTS восстанавливает соединения через повторяющееся ядро нижележащих акцепторных S-областей, где происходит большая часть CSR (3). Наше четкое открытие повышенного использования MH в соединениях CSR от ATM-дефицитных B-клеток мыши подтверждает вывод предыдущих исследований с более ограниченными соединениями, которые обнаружили повышенное использование MH в соединениях CSR в контексте дефицита ATM у мышей и человек (16, 17).
Поразительным результатом наших текущих открытий является то, что дефицит 53BP1 и дефицит лигазы 4 приводят к удивительно похожему влиянию на использование MH в Sμ-Sα-соединениях CSR, восстановленных из активированных клеток B-лимфомы Ch22F3. Таким образом, для соединений CSR на этих двух фонах частота MH-опосредованных соединений CSR, общее использование MH и частота MH более 4 п.н. вполне сопоставимы. Эта поразительная корреляция между использованием MH для соединения концов между этими двумя генотипами также применима к транслокационным соединениям между DSB, генерируемыми bait c-myc Cas9 / gRNA и Sμ / Sα, и к более общим соединениям транслокации в масштабе всего генома, подтверждая идею, что 53BP1 способствует соединению концов C-NHEJ во время CSR и, потенциально, других классов полногеномных DSB (11).Таким образом, в отсутствие 53BP1 может преобладать MH-опосредованный A-EJ. Значительная часть CSR-ассоциированных DSB, которые соединяются в отсутствие 53BP1, возникает из S-регионов, подвергшихся длинным резекциям, которые могут обнажать длинные одноцепочечные области, которые ингибируют C-NHEJ и, таким образом, вносят вклад в соединительный фенотип MH (3 ). Примечательно, однако, что дефицит лигазы 4 оказывает гораздо менее серьезное влияние на CSR, чем дефицит 53BP1, и не способствует почти такой же степени длинных S-резекций. Таким образом, драматический эффект дефицита 53BP1 на CSR и ориентацию восстановленных соединений CSR явно отражает дополнительные факторы, помимо доминирующего использования MH-смещенного пути A-EJ для CSR, потенциально включая нарушение привязки концов (3).Относительное увеличение MH-смещенных соединений CSR на других фонах также может более сильно отражать относительное влияние резекции конца (3), но требуется дополнительная работа, чтобы прояснить эту возможность, поскольку наш анализ надежно обнаруживает только длинные резекции (3). Наконец, умеренное влияние дефицита лигазы 4 на ориентационно-специфическую CSR также вовлекает факторы, помимо использования соединительных MH, в гораздо более серьезное влияние дефицита 53BP1 на ориентационную специфичность CSR.
В клетках Ch22F3, дефицитных по WT, ATM-, 53BP1 и лигазе 4, мы обнаружили, что частота использования MH в соединениях транслокации, общие паттерны и использование MH, а также использование длинных гомологий в приманке c-myc с транслокацией Sα переходы удивительно похожи на переходы Sμ-Sα CSR.Таким образом, отличительные фенотипы использования MH в отношении соединительного MH, используемого для CSR у WT и различных мутантных фонов, не ограничиваются соединениями между двумя повторяющимися последовательностями S-области или локализацией в домене локуса IgH . С другой стороны, паттерны MH соединений между c-myc DSB и общими DSB в масштабе всего генома у WT и различных протестированных мутантных фонов имели менее выраженные различия по сравнению с теми, которые наблюдались для Sμ-Sα CSR или c-myc приманка DSB к соединениям Sα.Наиболее ярким примером таких менее выраженных различий были соединения между DSB приманки c-myc и общими DSB в WT и ATM-дефицитных клетках Ch22F3, в которых уровень использования MH, о чем свидетельствует уровень прямых соединений, был существенно ниже. неотличимы. Чтобы проверить общность этого удивительного последнего открытия, мы также проанализировали транслокационные соединения в WT и ATM-дефицитных первичных нервных стволовых и прогениторных клетках и сделали аналогичные выводы в отношении сходства в использовании MH ( SI, приложение , рис.S7). Потребует дальнейшего изучения, почему DSB S-области значительно более склонны использовать MH для соединения концов DSB в отсутствие ATM и, в меньшей степени, других факторов DSBR, чем многие другие DSB в геноме. Однако отличительные особенности-кандидаты, которые могут вносить вклад, включают DSB S-областей, инициируемые AID, встречающиеся в очень повторяющихся последовательностях и подвергающиеся усиленной резекции в фонах с дефицитом DSBR.
Экспериментальные процедуры
Мыши.
ATM — / — , h3AX — / — , 53BP1 — / — и Rif1 f / f CD19 cre линий уже сообщалось ранее (3).Работа с мышами выполнялась в соответствии с протоколами, утвержденными Бостонской детской больницей.
Плазмиды и олигонуклеотиды.
Все олигонуклеотиды пРНК для CRISPR / Cas9 были клонированы в вектор pX330 (плазмида Addgene ID 42230). Все олигонуклеотиды перечислены в приложении SI , таблица S10.
Культура клеток.
Первичные В-клетки селезенки выделяли с помощью CD43-отрицательного набора для селекции и культивировали в среде R15 (RPMI1640, 15% FBS, 1-глутамат, 1 × пенициллин и стрептомицин).Первичную стимуляцию В-клеток выполняли анти-CD40 (1 мкг / мл; eBioscience) плюс IL4 (20 нг / мл; PeproTech) в течение 96 часов. Клетки Ch22F3 культивировали в среде R15 и стимулировали анти-CD40 (1 мкг / мл; eBioscience) и IL4 (20 нг / мл; PeproTech) плюс TGF-β (0,5 нг / мл; R&D Systems) в течение 72 часов. Нуклеофекцию плазмиды c-myc Cas9 проводили через 12 ч после стимуляции.
Сотовые линии.
Генерация некодирующих линий клеток Ch22F3 с удаленным аллелем была описана ранее (3).Для получения клеток ATM — / — клетки WT Ch22F3 сначала подвергали нуклеофекции с использованием 4D-нуклеофектора X (раствор SF, протокол CA-137; Lonza) с двумя векторами pX330 с последовательностью гРНК, нацеленной на ген ATM. Субклоны единичных клеток высевали в 96-луночные планшеты через 12 часов после нуклеофекции, и полученные клоны подвергали скринингу с помощью ПЦР и саузерн-блоттинга. Два независимых клона, использованных в этом исследовании, были подтверждены вестерн-блоттингом с антителом против α-ATM [(2C1) sc23921; Санта-Круз Биотехнология. Клетки 53BP1 — / — Ch22F3 были получены аналогично тому, как описано выше, и два полученных независимых клона были подтверждены с помощью саузерн-блоттинга и вестерн-блоттинга с антителом α-53BP1 (# 4908; Cell Signaling Technology).Ch22F3 с дефицитом лигазы 4 (клон 2) был получен аналогичным образом ранее (21).
HTGTS и анализ данных.
HTGTS-соединений из различных генетических фонов клеток Ch22F3 были выделены, как описано ранее (3). Описано выделение первичных HTGTS-соединений B-клеток (3). Анализ данных HTGTS, включая анализ соединений MH (21), всех библиотек HTGTS был выполнен и охарактеризован с использованием биоинформатического конвейера (v2), описанного ранее (3), а необработанные данные были отправлены в Национальный центр биотехнологической информации по экспрессии генов. Омнибус (GEO).MH определяется как самая длинная область на переключающем соединении с идеальной непрерывной идентичностью донор / акцептор. Последовательности с разрывом между донорной и акцепторной последовательностями рассматривались как вставки и исключались из расчетов. Прямые соединения определялись как соединения без MH и без вставок (MH = 0). Для построения паттерна MH прямые соединения и соединения с MH были объединены и отсортированы по длине, а количество соединений с указанной длиной MH было подсчитано и рассчитано как процент от общего числа соединений, отображенных в интересующей области.Координаты каждой области S для расчета описаны ранее (3).
Соединение смещения ориентации и расчет резекции S-области акцептора.
Соединения S-области акцептора, сопоставленные с Sα ( SI Приложение , рис. S4 A ), были разделены на шесть областей (обозначенных буквой f) в ориентации (-) или (+), как показано ниже: a | b | cd | e | f (-) (+).
Номера соединений, обозначенные в каждой из вышеуказанных областей, указанных в трех повторах данного генотипа ( SI Приложение , рис.S4 A ) были использованы для расчета отношения смещения инверсия (INV): удаление (DEL) следующим образом: Коэффициент смещения = (d + e) / (b + c). Процент соединений с длинной резекцией был рассчитан с помощью c / (b + c) × 100.
Статистический анализ.
Непарный двусторонний тест Стьюдента t использовался для проверки значимости различий между выборками. Значение P <0,05 считалось значимым.
Размещение данных.
Данные секвенирования HTGTS, проанализированные здесь, депонированы в базе данных GEO под номерами доступа.GSE71005, GSE106922 и GSE74356 ( SI Приложение , рис. S7).
Благодарности
Мы благодарим доктора Кефей Ю за предоставление линии клеток лигазы 4 — / — Ch22F3 (обозначенной клоном 1). Эта работа была поддержана грантом R01AI077595 Национальных институтов здравоохранения (для F.W.A.). J.D. поддерживается программой Гуандун по внедрению инновационных и предпринимательских команд (грант 2016ZT06S252). F.W.A. — следователь, З.Д. научный сотрудник Медицинского института Говарда Хьюза.
Сноски
Вклад авторов: R.A.P., X.Z., J.D. и F.W.A. спланированное исследование; R.A.P., X.Z., V.K., P.-C.W., J.K. и J.D. проводили исследования; Z.D. внесены новые реагенты / аналитические инструменты; R.A.P., X.Z., J.D. и F.W.A. проанализированные данные; и R.A.P., X.Z. и F.W.A. написал газету.
Рецензенты: J.C., Мемориальный онкологический центр им. Слоуна Кеттеринга; и K.Y., Университет штата Мичиган.
Авторы заявляют об отсутствии конфликта интересов.
Размещение данных: данные, представленные в этой статье, были депонированы в базе данных Gene Expression Omnibus (GEO), https://www.ncbi.nlm.nih.gov/geo (инвентарные номера GSE71005, GSE106922 и GSE74356).
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1719988115/-/DCSupplemental.
Какое будущее ждет маршрутизатор сотовой связи 5G?
Нужен ли маршрутизатор сотовой сети в 5G?
Когда я получаю этот вопрос, я начинаю задавать некоторые уточняющие вопросы.Иногда разговор приводит к обсуждению интегрированного транзитного соединения доступа (IAB). В других случаях обсуждение приводит к плану развития основной полосы частот и к тому, как функции маршрутизации интегрируются в платформы Ericsson RAN Compute. А в других случаях мы обсуждаем мобильные пограничные вычисления и виртуальную маршрутизацию.
Но после уточнений я обычно говорю: «Вы задаете неправильный вопрос». На самом деле вам нужно спросить: «Сколько функций маршрутизации потребуется для сотовой сети 5G?» Это совершенно другой вопрос… и мой ответ обычно не устраивает покупателя: «Это зависит от обстоятельств.”
Итак, теперь к вопросу «На чем?» Это сочетание трех основных факторов:
- Архитектура RAN
- RAN Модели развертывания
- Предлагаемый вариант использования 5G
Сочетание факторов влияет на требования к маршрутизации
Комбинация трех вышеуказанных факторов приводит к различным ответам на требования к маршрутизации. Я объясню на нескольких примерах.
Пример 1:
Архитектура RAN: распределенная RAN (DRAN)
Модель развертывания: концентратор и луч
Вариант использования: расширенная служба MBB
В этом случае функциональность маршрутизатора не критична.Однако узлы 5G с высокой пропускной способностью могут состоять из множества основных полос частот, которые требуют тесной координации друг с другом с использованием эластичной RAN. В этой ситуации CSR также действует как локальное соединение с агрегацией и основной полосой частот.
Пример 2:
Архитектура RAN: Централизованная RAN (CRAN)
Модель развертывания: десятки совмещенных базовых полос
Вариант использования: спектр услуг 5G от eMBB для увеличения емкости, фиксированный беспроводной доступ (FWA) для бытовых услуг и выделенные сетевые сегменты для конкретного предприятия варианты использования
В этом случае функциональность маршрутизации на сайте хаба CRAN может стать жизненно важной.
Дополнительные сведения
Обсуждения функциональности маршрутизации для сайтов 5G и ее расположения в сетевой архитектуре — это только начало. Я прошу клиента подумать о некоторых других критически важных функциях, необходимых для предоставления услуг 5G. Такие вещи, как:
Приоритезация и буферизация трафика
Мобильный трафик по своей природе является прерывистым, и нет обозримого конца потреблению мобильного видео.Видео составляло 60% мирового трафика в 2018 году и составит почти три четверти трафика к 2024 году (подробнее см. В последнем отчете Ericsson Mobility Report). Очевидно, что мобильная связь больше не связана с голосом и данными о максимальных усилиях! Операторы должны начать думать о том, как это повлияет на чувствительные ко времени варианты использования с низкой задержкой, совместно использующие транспортную сеть. Крайне важно иметь возможность классифицировать, планировать и определять приоритеты трафика от нескольких служб с различными требованиями. Кроме того, буферы должны быть достаточно большими, чтобы гарантировать пропускную способность для некоторых служб, обеспечивая при этом сохранение потокового контента и мультимедийных приложений с высоким QoE.Вдобавок к этому CSR 5G должен иметь возможность обрабатывать новые интерфейсы 5G, такие как eCPRI и F1, и для этого требуется собственная приоритезация из-за строгих требований к задержке.
Распределение времени и синхронизации
При использовании 5G радиостанции с дуплексной связью с временным разделением (TDD) развертываются вместе с другими радиостанциями TDD и FDD. Радиостанции TDD в значительной степени зависят от надежного источника фазовой и временной синхронизации для правильной работы. Если вы потеряете местный GPS, у вас будет всего час, прежде чем вы потеряете радиостанции TDD, которые управляют подавляющим большинством пропускной способности радиосайтов.Резервное копирование синхронизации через обратное соединение является обязательным. Подробнее о времени и синхронизации в сетях 5G в этом блоге, 5G — это все вовремя.
Мультидоменное нарезание
Достижения в области разделения сети в первую очередь сосредоточены на базовой сети. Что действительно необходимо, так это многодоменное разделение на RAN, транспортную и базовую сеть. Это может обеспечить управление трафиком E2E для настройки и оптимизации транспорта с использованием SDNc домена (программно определяемых сетевых контроллеров) и иерархических контроллеров SDN для охвата нескольких транспортных доменов.
Сетевая безопасность
Сетевая безопасность имеет решающее значение для реализации обещаний 5G и сетевого сегмента. С помощью сегментации сети операторы мобильной связи могут разделить свою физическую сеть на несколько виртуальных сетей. Нарезка позволяет поставщикам услуг экономично масштабировать различные типы услуг для разных сегментов клиентов. Однако поставщики услуг должны гарантировать, что разные виртуальные сетевые срезы адекватно изолированы друг от друга. Приложения с «высоким уровнем безопасности» не должны быть доступны из сегмента «с более низким уровнем безопасности».В некоторых случаях будет достаточно функциональности традиционного шлюза безопасности (SecGW). В других случаях из-за требований к производительности может потребоваться распределение SecGW на сайт 5G RAN или концентратор CRAN. В дополнение к разделению сети добавление внутренних сетей и небольших ячеек через ненадежные домены также требует новых подходов к безопасности на транспортном уровне.
Тип сайта РАН
Расположение функций маршрутизации также зависит от доступности и стоимости недвижимости на сайте RAN.Площадки могут варьироваться от рельсовых до установленных на опорах, уличных шкафов, внутренних хижин и центральных офисов. При использовании архитектур 5G DRAN, CRAN и VRAN, маршрутизация и другие важные функции, описанные выше, должны быть доступны для соответствия типу вашего RAN-сайта. При планировании 5G обязательно учитывайте размер устройства транспортной сети, форм-факторы и степень защиты.
Где должны находиться функции маршрутизации в сети 5G?
Как обсуждалось выше, потребности в функциональности чистой маршрутизации могут варьироваться в зависимости от архитектуры 5G RAN, моделей развертывания и сценариев использования.А как насчет всех дополнительных аспектов, которые мы обсудили? Где вся эта функциональность должна находиться на сайте RAN?
- В gNB? Неа. Добавление всех этих функций к gNB потребует больше места и энергии, ограничит варианты развертывания и в конечном итоге потребует от клиента дополнительных
- На текущем маршрутизаторе сотовой связи? Неа. Маршрутизаторы сайтов, представленные сегодня на рынке, не имеют этой функции. Они не могут поддерживать ряд типов сайтов и моделей развертывания, необходимых для 5G
- На хранении / вычислении в расположении MEC? Возможно, но он должен быть устойчив к высоким температурам и рентабельным для масштабирования до мощности, требуемой 5G
.
Итак, что можно сделать?
Компания Ericsson, лидер в области 5G, задумалась об этом.Наше решение — предоставить эти критически важные функции нашим клиентам в семействе продуктов Router 6000. Ericsson Router 6000 — это самая надежная, эффективная и экономичная платформа для предоставления услуг 5G на рынке сегодня. Он предоставляет глубокие буферы, HQoS, встроенный источник синхронизации stratum 3E, возможность распределенного шлюза IPSec и доступен во всех форм-факторах RAN, что обеспечивает согласованность в ваших развертываниях.
Ой… кстати… он тоже трассирует.
Связанное чтение:
Fierce eBrief: решение проблемы транзитной сети 5G
Транспортные решения Ericsson 5G
Белая книга для интенсивного чтения: определение транспортной сети для 5G
* Зачем вашей сети 5G транспорт 5G
Центральная серозная ретинопатия — Macular Society
Что такое центральная серозная ретинопатия (CSR)?
Центральная серозная ретинопатия (CSR) также известна как центральная серозная хориоретинопатия или CSCR.При CSR макула отделяется от ткани глаза позади нее, и в созданном пространстве накапливается жидкость.
Это в шесть раз чаще встречается у мужчин, чем у женщин, и чаще всего поражает людей в возрасте от 20 до 50 лет.
Обычно поражается только один глаз, и до половины людей, получивших это однажды, заболеют снова.
Что такое макула?
Макула — это часть сетчатки в задней части глаза. Его диаметр составляет всего около 5 мм, но он отвечает за все наше центральное зрение, большую часть нашего цветового зрения и мельчайшие детали, которые мы видим.Здоровое пятно имеет толщину около 250 микрон (четверть миллиметра).
Что вызывает центральную серозную ретинопатию?
К сожалению, неизвестно, почему макула отделяется от ткани глаза и накапливается жидкость, но исследования показывают, что стресс может усугубить ситуацию.
Группа препаратов, называемых кортикостероидами, также может вызвать CSR или усугубить ситуацию, и повышенный уровень гормона кортизола (который, как правило, выше у людей с апноэ во сне или синдромом Кушинга), похоже, также повышает вероятность этого.
Есть также некоторые свидетельства того, что это может быть вызвано бактериальной инфекцией, называемой H. pylori, некоторыми другими лекарствами или даже беременностью.
Каковы симптомы центральной серозной ретинопатии?
По мере того, как макула набухает и отделяется от тканей позади нее, вы можете заметить нечеткое или отсутствующее пятно в центральном зрении, искажение или что-то меньшее, чем обычно.
Вы также можете заметить изменения в своем цветовом зрении или испытывать трудности с адаптацией к изменениям уровня освещенности.
Как лечится центральная серозная ретинопатия?
В большинстве случаев жидкость реабсорбируется без какого-либо лечения. Большинство людей с КСО вернут большую часть или все прежнее зрение. Если через три-шесть месяцев жидкость все еще остается, вам может потребоваться лечение, чтобы предотвратить более длительное повреждение макулы.
Одним из вариантов лечения является использование лазера для закрытия утечки, но его нельзя использовать слишком близко к центру макулы, так как это может повредить ваше центральное зрение.
В этом случае вам может быть предложена фотодинамическая терапия. Это когда лекарство вводится в кровоток и активируется в глазу с помощью низкоэнергетического лазера, который не повредит желтое пятно.
Вам также следует помочь внести любые изменения в образ жизни, которые могут помочь, например уменьшить стресс или отменить любые лекарства, которые могут ухудшить состояние.
Текущие исследования
Одно недавнее исследование изучало, может ли лекарство, используемое в настоящее время для лечения сердечной недостаточности, лечить CSR, но, к сожалению, обнаружило, что оно не лучше, чем плацебо (фиктивное лечение).
Другое исследование тестирует терапию тепловым шоком: использование низкоэнергетического лазера для обновления клеток сетчатки и помощи им в более быстром избавлении от жидкости.
Узнайте больше о последних исследованиях состояний желтого пятна и о том, как вы можете помочь нам быстрее найти лекарство.